КУРСОВА РОБОТА з загальної екології
На тему: «Демекологія ЯК екологія популяцій»
1. Організація на популяційному рівні
2. Динаміка популяцій. Загальні уявлення та поняття
3. Структура популяцій
4. Типи взаємовідносин між популяціями
5. Ріст чисельності популяції, криві росту та виживання
При будь-якому біологічному дослідженні виникає необхідність розмежування матеріалу, який ми маємо вивчити на певні одиниці, далі яких розподіл уже не проводиться. В генетиці
це ген,
в систематиці
- вид,
при вивченні екосистем біогеоценозу, характеристику якого ми будемо вивчати в наступних темах, це популяція,
яка володіє певними характеристиками системи. Популяція є цеглиною, з якої і розпочинається власне екологія організмів будь-якої екосистеми, є першою надорганізмовою біологічною системою.
Термін "популяція" запозичений з демографії В. Іогансеном у 1905 р. на позначення групи особин одного виду, які здатні до вільного схрещування, населяють певний простір протягом життя багатьох поколінь і відокремлені від інших подібних угруповань. Найістотнішими параметрами популяцій є динаміка чисельності особин, структура
- вікова, статева, просторова, ієрархічна і густота населення.
Як і більшість понять екології, термін "популяція"
неоднозначний. Спеціалісти різного профілю, виділяючи популяції у природі, користуються різними критеріями.
У генетиці
популяції розглядають як структурні одиниці, що утворюють таксономічний вид. Обов'язковою умовою виділення окремої популяції в цьому випадку є наявність вільного обміну генами серед усіма особинами даної популяції, що забезпечує спільність генофонду. Такі популяції називають менделівськими.
Вони невеликі за розмірами. Річ у тім, що у рослин при вітрозапиленні та при запиленні комахами пилок розноситься, як правило, на невеликі віддалі. У вітрозапильних чагарників та дерев вона не перевищує 100-150 м. В ентомофільних рослин комахи можуть розносити пилок на віддалі, не більші за 1-3 км, але звичайно взяток береться в радіусі всього декількох сот метрів. Невеликі шлюбні території й у тварин. У менделівських популяціях особини теоретично повинні бути повністю ідентичні. Але звичайно це не спостерігається. Така ідентичність має місце лише тоді, коли живі організми розмножуються нестатевим шляхом або автогамно. Так формуються популяції кореневищної рослини пирію, гермафродитних тварин типу паразитичного ціп'яка або партеногенетичних скельних ящірок.
У ботаніці
критеріями виділення популяції служить її розміщення в межах певного біоценозу. Такі популяції називають ценотичними. Розміри ценопопуляцій можуть бути різними. У невеликих ценозах вони невеликі, а в монотонно-однорідних типу тайгового лісу можуть охоплювати території у сотні та тисячі гектарів і складатися з багатьох мільйонів особин.
В екології та зоології популяції частіше виділяють за ознаками їхнього розподілу на певній території та достатній відмежованості від популяцій того ж виду. У цьому випадку популяцію називають локальною.
Механізми ізоляції окремих популяцій бувають двох типів: територіальні
та репродуктивні.
У першому випадку межами між популяціями виступають певні бар'єри: гірський хребет, річка і т.п. У другому випадку ізоляцією є неможливість схрещування між особинами різних популяцій. Наприклад, особини конюшини гірської на південному та на північному схилах одного й того ж пагорба можуть належати до різних популяцій, оскільки на північному схилі цвітіння починається тоді, коли цвітіння на південному схилі вже пройшло.
Вивчення популяційної структури виду має надзвичайно важливе теоретичне і практичне значення при здійсненні заходів з раціонального природокористування. Важливо знати загальні біологічні властивості виду, а також як впливає зовнішнє середовище на його формування. Знання екології популяцій особливо необхідні під час цілеспрямованої інтродукції. Досвід попередніх років показує, що навіть „корисні" види, їх інтродукція може призвести до повної перебудови екосистеми та зникнення аборигенних видів. Як класичний приклад можна згадати інтродукцію кролика, який за короткий період став основним конкурентом усім травоїдним тваринам на Австралійському континенті. Слід відмітити, що це явище краще прослідкувати на тваринах і набагато складніше на рослинах. Разом з тим рослини інтродуценти є не менш активними чинниками при деструктивних змінах екосистем. Як правило, інтродукований вид є агресивніший по відношенню до аборигенних видів, йому на початкових етапах людина створює більш сприятливі умови.
Отже, штучно скорочується час адаптації виду до нових умов.
Нерідко нові види привносять з собою і види, які трофічно з ними пов'язані, але як не дивно нерідко шкодять більше аборигенним видам.
Вивчаючи популяцію, ми визначаємо не тільки її сучасний стан, але намагаємось провести аналіз стану даної популяції в минулому та провести прогноз її росту під впливом різних факторів.
Розглянемо, які фактори впливають на чисельність популяції - загальну кількість особин, які належать до однієї популяції на даній території або в даному об'ємі. Повінь, пожежа, град, раптові морози, посуха, бурелом, Надмірне застосування хімпрепаратів, реконструкція ландшафту, вселення нових видів хижаків, паразитів, епідемії - все це може призвести до багаторазового скорочення чисельності популяції; навіть до повної її загибелі. Загибель або різке скорочення чисельності популяції, як правило, викликає ланцюгову реакцію в біоценозі і може спричинити коливання чисельності популяцій інших видів. Аналіз причин загибелі окремих видів свідчить про те, що зникнення одного виду рослин викликає загибель від 3-4 до 20-30 і навіть більше видів тварин.
При зниженні чисельності популяції зменшуються можливості обміну генетичною інформацією, що призводить до зниження життєздатності. Тому пізнання закономірностей динаміки чисельності популяцій має першорядне значення. У кожний конкретний момент будь-яка популяція складається з певної кількості особин, але ця величина досить динамічна. Часто вона залежить від народжуваності і смертності у популяції.
Народжуваність - кількість нових особин, які з'явилися за одиницю часу у процесі розмноження.
Смертність - кількість особин, які загинули за одиницю часу.
І Відтворення
потомства
- головне джерело поповнення популяції. У
рослин - це кількість насіння, у риб
- ікринок, у птахів - яєць і т.п.
Швидкість зростання популяції визначається біотичним потенціалом. Біотичний потенціал -
це кількість нащадків, яку здатна дати одна материнська особина. В одних видів біотичний потенціал може перевищувати мільярд, у інших - обмежуватись кількома десятками. Види, що живуть у сприятливих умовах і добре пристосовані до виживання, мають низький біотичний потенціал; і навпаки, висока смертність зумовлює надзвичайну плодовитість. Наприклад, риби, які не турбуються про потомство, відкладають тисячі і навіть мільйони ікринок. Біотичний потенціал деяких видів риб сягають - у місяць - до 3 млрд., а в акул, для яких характерне яйце живонародження, він обмежений десятками. Більшість шкідливих комах здатні плодити від кількасот до тисячі особин. Для стабілізації чисельності популяцій достатньо, щоб до розмноження доживало стільки потомків, скільки було батьків. Якщо відсоток виживання вищий за відсоток рівноваги, популяція зростає, якщо нижчий -зменшується. Це необхідно враховувати як при боротьбі із шкідниками, так і при охороні зникаючих видів. Чисельність будь-якої популяції коливається під впливом дії біотичних і абіотичних факторів. Існує поняття мінімальної чисельності
популяції, нижче якої популяція перестає існувати. В деяких випадках (рослинні угрупування) доцільно використовувати не показники кількості особин у популяції, а їх біомасу.
При характеристиці популяції часто використовують поняття щільності,
тобто кількості особин певного виду на одиницю площі. Особливо активно даний аспект використовується в лісовому господарстві, де показник щільності кормових рослин використовується для визначення параметрів ємності мисливських угідь по відношенню до мисливської фауни.
3. Структура популяцій
Як вказувалось, структура популяції є однією із ознак популяції. Виявлення структурних елементів популяції, дало змогу більш чітко сформувати сучасні уявлення про популяцію. Крім того, виділення структурних елементів популяції дає змогу проводити більш ефективний аналіз стану популяції того чи іншого виду. Особливо це стосується видів, які
перебувають у стані волі але експлуатуються людиною.
Просторова структура
- закономірне розміщення особин даної популяції в просторі в певний період часу існування популяції. Аналіз любої популяції показує, що особини розміщуються по різному.
У рослинних угрупованнях просторова структура популяцій виявляється через характерне розміщення особин даного виду: вони можуть виступати поодинці, парами, групами або ж скупченнями. їх розміщення залежить від біологічних особливостей виду, стадії розвитку популяції, умов місцезростання. Відносно ж тварин, то тут важливим фактором є пора року (лялечка зимує в лісовій підстилці, гусениця живе в кроні дерева).
В багатьох випадках трапляється скупчення особин одного виду у біогрупи або парцели. В грабово-буковому лісі таку горизонтальну структуру творять береза, граб, липа, жостір, а з трав'яних рослин - яглиця, печіночниця, копитень, плющ, осока волосиста. Таке скупчення особин дає їм можливість витримати дію несприятливих умов середовища, а також міжвидову конкуренцію.
Розподіл особин популяції може бути:
· Рівномірним
· Груповим
· Випадковим
Випадковий
розподіл в природі зустрічається дуже рідко тільки в тих випадках, коли середовище однорідне, а організми не намагаються об'єднатися в групи.
Рівномірний
розподіл може зустрічатись там, де між: особинами дуже сильна конкуренція або існує антагонізм, який сприяє рівномірному розподілу в просторі.
Груповий
- якщо особини в популяції намагаються утворити групи певного розміру. Деколи така структура нагадує випадковий розподіл. Такий розподіл зустрічається досить часто при утворенні пар, скупчень.
Такі скупчення виникають:
· як наслідок змін умов середовища (передміграційні скупчення, міграційні скупчення);
· як реакція на добові та сезонні зміни погоди; у зв'язку з процесом розмноження.
Будь-яке із таких скупчень має велике біологічне значення. У особин, які об'єднуються у певну групу, у порівнянні із одинокими особинами, спостерігається зниження смертності завдяки здатності групи протидіяти (певним чином) несприятливим умовам середовища.
Важливою ознакою територіальної структури популяції є знадність особини займати певну ділянку, яка є індивідуальною ознакою і для кожного виду різна. Підтримання меж ділянки досягається завдяки територіальності.
Вважається (Ю. Одум), що територіальність це будь-який метод підтримання меж ділянки, яка необхідна для повноцінного функціонування особини. Існують різні механізми підтримання територіальних меж. У нижчих рослин і тварин це переважно алелопатія,
у вищих тварин - захист території.
Яскравим прикладом територіальності є спів птахів, мічення території ссавцями і т.п. Явище територіальності сприяє регуляції чисельності популяції на рівні, який є нижчим від рівня насичення. При збільшенні щільності включаються регулюючі механізми - зниження репродуктивної активності, агресивність, безпліддя тощо. Цікавим фактом є те, що навіть штучне підтримання харчового ресурсу (в лабораторних умовах це можливо) не може компенсувати явище територіальності.
Вікова структура
- закономірне співвідношення різних вікових груп популяції.
При аналізі вікових категорій виявляється, дане явище має певну закономірність і стосується переважно організмів, яким властиве статеве розмноження. Разом з тим, реально існуючими є і одновікові популяції штучних систем - агроекосистем, де людина висіваючи певні культури створює одновікові популяції. В природі (при врахуванні віку насіння) такого не існує.
Порушення вікової структури веде до зниження репродуктивних властивостей популяції як наслідок зниження чисельності. Нехтування віковою структурою популяції веде до деструктивних змін системи (вітровали, ерозії, зсуви, міграції і т.п.).
Статева структура
- закономірний розподіл особин даної групи
IIпо статі.
Теоретично співвідношення статі однакове 1:1, але в природі рідко зустрічається таке співвідношення. Загалом вік особини популяції можна розділити на три періоди: до-репродуктивний, репродуктивний, пост-репродуктивний. Показники співвідношення статі є індивідуальною характеристикою популяцій різних видів. Помічено, що статева структура знаходиться у взаємозв'язку із віковою структурою. Тобто, якщо ми розмежуємо популяцію по віку, то різні вікові групи будуть представлені різним співвідношенням статі.
Отже, на наведеному прикладі бачимо, що при загальній чисельності 1526 особин різних вікові групи представлені різним співвідношенням Сі .і гей. В до-репродуктивний
період воно близьке до співвідношення 1:1, але и
репродуктивний
період воно наближається до співвідношення 1:3 і в пост-рспродуктивний
період близьке 1:1.
Основна функція підтримання чисельності особин лягає на особин І ні продуктивного періоду. Завдяки цьому забезпечується відносна сталість чисельності популяції. Зростання чисельності популяції може відбуватись 11рм збільшенні кількості особин репродуктивного віку, але в послідуючому н 1.11 ні популяції, в певний період свого існування, буде йти затримка росту, чисельності, зростання смертності і тільки по встановленню оптимального співвідношення статі чисельність особин даної популяції і коливатись в певних межах.
Теорія ієрархічної структури багатьма дослідниками відкидається. Але, разом з тим, будь-яку популяцію можна розмежувати за ієрархічною ознакою, так як кожна особина в популяції виконує свою, тільки їй властиву функцію. Ієрархія популяції проявляється у формі організації популяції: поодинокий спосіб життя, сімейний спосіб життя, зграя, стадо, колонія, прайд.
Як відмічав К.Лоренц,
батько етології (науки про поведінку), ієрархія популяції побудована на принципі домінантності та підпорядкованості. Всі особини популяції підпорядковані цьому принципу, а при вибуванні особини якоїсь ланки іде швидка перебудова ієрархічної структури популяції. Багато організмів мають дуже складну ієрархічну структуру (бджоли, мурашки і т.д.). Знання ієрархічної структури популяції дає людині потужний механізм управління популяцією.
Знання про структуру популяцій
є основою для раціонального природо-користуванпя. Контроль за віковими, статевими, ієрархічними показниками популяцій в природі дозволяє експлуатувати природній ресурс без істотних змін чисельності особин. Нажаль, слід відмітити, що в багатьох випадках аналіз структури популяції виду є поверховим і, як наслідок, іде перевикористання і послідуюче пригнічення популяції та реконструкція системи, складовим елементом якого і є популяція.
Дуже рідко біолог, еколог вивчає тільки динаміку однієї популяції. Практично вивчення однієї популяції веде до вивчення популяцій вищих і нижчих трофічних рівнів. Існує цілий ряд добре вивчених типів відносин між популяціями різних видів (міжвидові взаємовідносини). На певному трофічному рівні це може бути міжвидова конкуренція,
тобто конкуренція між особинами різних видів за доступні ресурси їжі і простір. Візьмемо, наприклад, систему водно - болотних територій. Міжвидова конкуренція між птахами тут проходить за трофічні можливості досить непомітно. Кулики, які добувають їжу по береговій лінії, не створюють конкуренції качиним, які добувають їжу іншим способом (цідять воду). Але, разом з тим ми можемо помітити конкуренцію серед самих качиних. Вид, кількість якого буде досить великою, і буде більш пластичним, тобто, він здатний використовувати широкий спектр їжі і буде конкурентом менш пластичному виду, який має більш вузький діапазон використання. Для водно-болотних угідь Паннонської (Середньодунайської) низовини цим видом є крижень, чисельність якого найбільша. Популяції інших видів знаходяться ніби в пригніченому стані і, порівняно з популяцією крижня, менш чисельні. Популяції, які знаходять на різних трофічних рівнях, також взаємодіють одна на одну. В даному випадку ми маємо справу з трофічною залежністю популяцій окремих видів. Як було сказано вище, в екосистемі між популяціями можуть виникнути конкурентні відносини за будь-який із широкого спектру ресурсів (їжу, простір, світло, і т.п.). Якщо два види будуть знаходитись на одному трофічному рівні, то між ними обов'язково буде присутня конкуренція за їжу. В процесі еволюції організми навіть одного трофічного рівня адаптуються, а точніше відбувається більш вузька спеціалізація кожного виду, за рахунок чого конкуренція зводиться до мінімуму. Ю. Одум, аналізуючи взаємодії двох популяцій, наводить 9 типів зв'язків:
нейтралізм -
популяції не діють одна на одну;
конкуренція взаємного
подавления
-
обидві популяції чинять тиск одна на одну;
конкуренція за ресурси
- популяції негативно впливають одна на одну, борючись за їжу;
аменсалізм -
одна популяція чинить тиск на іншу, при цьому сама не зазнає негативних змін;
паразитизм, хижацтво
- одна популяція негативно діє на іншу шляхом активно або пасивного нападу, при цьому залежить від стану другої;
коменсалізм
- одна популяція, об'єднуючись, бере з цього користь (наприкл., Риба-лоцман-акула);
пропюкооперація
- обидві популяції мають користь;
мутуиліім, симбіоз -
зв'язок двох популяцій, необхідний для існування
(наприкл., Рак-самітник-актинія, лишайник, мікориза).
Розглянемо деякі з них.
Взаємовідносини "хижак
– жертва".
Хижацтво - це спосіб добування їжі. Екологічна група хижаки мають цілий ряд морфо-анатомічних пристосувань для нападу, умертвіння та поїдання жертви. Взаємовідносини хижак-жертва найбільш доступна для спостереження і дослідження модель взаємовідносин. Знаходячись на вершині трофічного ланцюга, чисельність хижака знаходиться в певній залежності від чисельності жертви. Якщо прослідкувати криві росту чисельності хижака і його жертви, то помітним є зміщення вершини фаз максимальної чисельності хижака і його жертви.
Таке зміщення максимальних точок легко пояснити: зростання кількості жертви веде до збільшення кількості хижака, так як є основою його арчового раціону. Через деякий час кількість жертви зменшується, що в свою чергу веде за загибелі хижака у зв'язку з відсутністю достатньої кількості харчового ресурсу. Важливо відмітити, що роль хижака в даному випадку не треба переоцінювати, але і не відкидати. В окремих випадках чисельність популяції залежить від іншого типу взаємовідносин: "хазяїн - паразит".
Паразитизм специфічний тип взаємовідносин між організмами. Стратегія паразита спрямована на якомога довше використання свого хазяїна, але при надмірному зараженні особини паразитами знижуються імунні властивості організму і особина гине. Отже, паразит сприяє загибелі особини, але не є причиною смерті. Якщо прослідкувати закономірності коливання чисельності хазяїна і паразита, то, як і у випадку з взаємовідносинами хижак-жертва, відмічається зміщення точок максимальних чисельностей паразита і хазяїна.
Розміри популяції можуть зростати в результаті імміграції (додаються особини ззовні) або за рахунок розмноження особин. На коливання чисельності суттєвий вплив мають кліматичні умови, які ми з вами розглядали в попередньому розділі (факторіальна екологія - температура, вологість і т.д.). Нерідко лімітуючим фактором, як ми розуміємо, виступають вороги, їжа і. т.п. Коливання чисельності проходить циклічно і їх можна назвати циклами. Але дослідження таких циклів вимагає довгого періоду часу і залежить від того коливання, популяції якого виду ми намагаємось встановити. Якщо врахувати період настання статевої зрілості, вагітність то ми побачимо, що у кожного виду вона різна. У маленької тваринки, такої як бурозубка, ці періоди будуть набагато коротшими від таких, як у копитних, слонів. Тобто, щоб прослідкувати цей процес, екологу необхідно володіти інформацією за той відрізок часу, за який проходить багаточисельна зміна генерацій (поколінь) і знати умови існування даної популяції. Набагато простіше цю інформацію можна здобути в лабораторних умовах, де в процесі постановки експерименту дослідник інколи штучно, а деколи і підсвідомо створює сприятливі умови існування (пацюк, дрозофіла і т.д.). Коливання чисельності графічно можна зобразити у вигляді синусоїди, для побудови якої необхідно проводити дослідження протягом довгого періоду часу. Дана синусоїда буде складатись із фрагментів, що можуть відхилятись від ідеальної кривої. Важливим моментом є той факт, що коливний процес буде відбуватись навколо уявної лінії, яка і буде ідеальним графічним вираженням чисельності популяції. Також слід відмітити, що коливання чисельності особин популяції можливе в певних межах, тобто, існує поняття мінімальної чисельності популяції. Якщо чисельність особин сягає показників нижчих від мінімальної чисельності, то вона зникає.
Розміри популяції можуть бути непостійними в результаті зміни плодючості, смертності, а нерідко і того і другого. При вивченні розмірів популяцій і їх змін завжди намагаються встановити ключовий фактор -
фактор, який відповідає за найбільшу частину змін, які відбуваються при зміні поколінь. Як правило, цей ключовий фактор впливає на смертність.
Слід розуміти, що коливання розмірів популяції проходить не хаотично. Насправді
с
ряд факторів, які втримують стан популяції в певних межах. Цими факторами є ті фактори, які знижують чисельність і сприяють смертності і найкраще діють при збільшенні щільності. Такими факторами можуть бути недостатня кількість їжі, збільшення кількості ворогів тощо.
5. Ріст чисельності популяції, криві росту та виживання
Якщо народжуваність у популяції буде перевищувати смертність, то дана популяція буде зростати. Яскравим прикладом такого явища є ріст народонаселення Землі. Було підраховано, що тільки за період XX ст. відбувся зріст народонаселення більш ніж удвічі. Тобто, внаслідок якісного стрибка людства, науково-технічного прогресу людство створило певні умови, які і призвели до такого різкого зросту.
Загальний хід зміни чисельності особин в популяції визначається рівнянням:
Nt
+1
= Nt
+ B-D + I - Е,
де N - чисельність особин в популяції, В - народжуваність, D -смертність, І - імміграція, Е - еміграція, t - час. Розміри популяції можуть зростати або за рахунок великої народжуваності, або за рахунок високої імміграції, або за рахунок поєднання обох цих факторів. Знижує розмір популяції смертність та еміграція особин за її межі.
Щоб ясно собі уявити закономірності росту популяції, доцільно
розглянути модель росту популяції дріжджів, які потрапили на свіжу
культурну речовину (рис. 1).

Рис. 1 - Модель росту чисельності популяції дріжджів
В такому новому та сприятливому середовищі умови для росту популяції оптимальні і буде спостерігатись експоненціальний ріст популяції. Після попадання у свіже поживне середовище ріст буде йти поступово, досягаючи максимальної чисельності. Затримка росту популяції на початкових етапах пов'язана із адаптацією до нових умов середовища. Намальована нами крива - це експоненціальна або логарифмічна крива. В наступних етапах життєдіяльності популяції настає період, коли експоненціальний розвиток неможливий. Таке може відбуватись з різних причин зменшення ресурсів живлення, накопичення продуктів метаболізму тощо. Як наслідок - процес росту популяції поступово уповільнюється і крива росту набуває S- подібну форму.
Існує і інший тип росту чисельності популяції, коли експоненціальний ріст продовжується до раптового зменшення кількості організмів (рис. 2).
Дане явище може відбуватись за рахунок різкого зменшення ресурсу, території тощо. Такий тип кривої росту дістав назву J - подібної кривої. Слід відмітити, що в обох випадках експоненціальний ріст відмічається на початкових етапах росту.
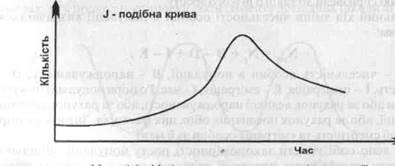
Рис. 2 - Модель росту чисельності популяції
Отже, ми розглянули дві моделі росту популяції. Разом з тим, слід відмітити, що побудова таких кривих можлива тільки за умови більш менш стабільного існування екосистеми. Тобто, там, де чинники системи не діють як лімітуючи на ріст популяції. Змальовані нами моделі в чистому вигляді, як правило, в природі не існують. Якщо з деякими схожостями ми можемо зустрітись в природі при розселенні і освоєнні видом нових територій, що наглядно можна проілюструвати розселенням горлиці кільчастої в Центральній Європі, то на територіях, де види інтродуценти вже ввійшли до складу екосистем, такого спостерігатись не буде. Разом з тим, дані моделі дають нам змогу розуміти закономірності росту чисельності популяцій, прогнозувати поведінку виду в нових умовах, управляти і корегувати чисельність "корисних"
і "шкідливих"
видів.
Одним із основних факторів, які впливають на розміри популяції, є процент особин, які гинуть до досягнення ними статевої зрілості. Для того щоб чисельність популяції залишалась сталою, в середньому тільки два потомки кожної пари повинні доживати до репродуктивного віку. Щоб отримати криву виживання, нам необхідно почати з певної популяції новонароджених особин і потім відмічати кількість особин, що вижили в залежності від часу.
Будуючи криві виживання для окремих видів, ми можемо визначати смертність для особин різного віку і, таким чином, вияснити, в якому віці даний вид найбільш вразливий. Якщо ми встановимо причини смерті, можемо зрозуміти як регулюється величина популяції.
Криву виживання можна отримати, якщо розпочати з певної популяції новонароджених особин і на майбутнє відмічати число або відсоток особин, які залишаються жити в залежності від часу. Більшість тварин та рослин старіють, яке в першу чергу проявляється у зменшенні кількості особин після досягнення репродуктивного періоду, (рис. 3). Причинами цього явища є велика кількість факторів, але, як правило, в пост-репродуктивний період організм поступово втрачає свою захисну здатність. Крива А - характерна для видів, де смертність є більш менш сталою одиницею у всі періоди розвитку. Для більшості безхребетних є типовою така крива. Щось подібне було отримано для „популяції" чашок в кафетерії.
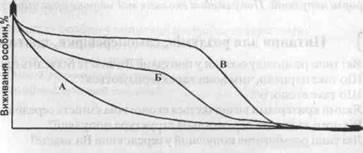
Рис. 3 - Типи кривих виживання
Крива Б - характерна для популяцій організмів з високими показниками смертності в ранній до-репродуктивний період. Така крива характерна для муфлонів, гірських кіз. Крива В - близька до ідеальної кривої, так як бачимо, що смертність довгий період часу уступає віку, а старіння є головним фактором смертності. Прикладом може бути популяція людей на нашій планеті. Велика кількість людей помирає внаслідок старіння, але середній вік не перевищує 75 років. Невелике відхилення на початкових фазах пов'язано із дитячою (до-репродуктивною) смертністю.
Завершуючи розгляд питань динаміки чисельності популяції, слід відмітити, що процес коливання чисельності неперервний і може змінюватись в часі, як наслідок адаптаційних змін. Зникнення цього явища можливе тільки у зв'язку із зникненням виду. Питання динаміки популяцій є основою для розуміння більш широких питань, таких як
динаміка угруповання, екосистеми, біосфери в цілому.
|