СОДЕРЖАНИЕ
1. Цепи и циклы питания.
2. Экологическая пирамида.
3. Поток веществ и энергии.
Список использованной литературы
1. Цепи и циклы питания
Как уже отмечалось, между организмами биоценоза устанавливаются прочные пищевые взаимоотношения. В результате возникают цепи питания. Они и объединяют прямо или косвенно большую группу организмов в единый комплекс. Цепь питания обычно состоит из трех основных звеньев.
Первое звено образуют так называемые продуценты или производители. Это автотрофные зеленые растения, которые в процессе фотосинтеза создают органическое вещество — первичную биологическую продукцию — и аккумулируют солнечную энергию.
Второе звено представлено консументами, т.е. потребителями, — гетеротрофными организмами, питающимися растениями или другими гетеротрофами. Различают консументы первого порядка (фитофаги), второго порядка (плотоядные животные, питающиеся фитофагами), третьего порядка (хищники, питающиеся другими животными) и т. д.
Третье звено — это редуценты, или деструкторы, — разрушители органического вещества. К ним относятся микроорганизмы, грибы и организмы, питающиеся мертвым органическим веществом и минерализующие его до простых неорганических соединений.
В каждой цепи питания формируются определенные трофические уровни, характеризующиеся различной интенсивностью протекания потока веществ и энергии. Зеленые растения — созидатели органического вещества — образуют первый трофический уровень, фитофаги — второй, плотоядные животные — третий и т. д.
Все звенья цепи питания взаимосвязаны и взаимозависимы. Между ними от первого к последнему осуществляется передача вещества и энергии. Суть этого явления будет рассмотрена ниже. Сейчас важно обратить внимание на то, что при передаче энергии с одного трофического уровня на другой происходитее потеря. В результате цепь питания не может быть длинной, как это иногда изображают графически. Скорее всего она состоит из 4—6 звеньев (рис. 1). Однако такие цепи в чистом виде в природе обычно не встречаются, поскольку одни и те же виды могут быть одновременно в разных звеньях. Это происходит потому, что монофагов в природе чрезвычайно мало, чаще встречаются олигофаги и полифаги. Рассмотрим, к примеру, цепь питания, основным звеном которой является капуста. Следующим звеном в ней будут гусеницы капустной белянки, капустной моли, капустной совки, зайцы, т.е. все животные, питающиеся капустой. Следовательно, капуста здесь выступает основным звеном многих цепей, поскольку от последующего звена (гусеницы, зайцы и др.) могут тянуться еще цепи. Причем каждый организм, питающийся капустой, одновременно может быть составной частью не одной, а нескольких цепей. Так, заяц, поедая разные растения, входит как консумент первого порядка в большое количество цепей питания. Хищники также питаются различными растительноядными и плотоядными животными, а потому являются звеньями многих цепей.

Рис. 1. Упрощённая схема цепи питания
Подобные общие звенья связывают цепи питания в сложную систему. В результате в каждом биоценозе исторически формируются комплексы цепей питания, представляющие собой единое целое. Так создаются циклы, или сети, питания (рис. 2). Если принять во внимание, что практически каждый организм цепи питания выступает в роли хозяина по крайней мере одного, а чаще нескольких паразитов, составляющих в свою очередь звенья других цепей, то нетрудно вообразить всю сложность циклов питания биоценоза.
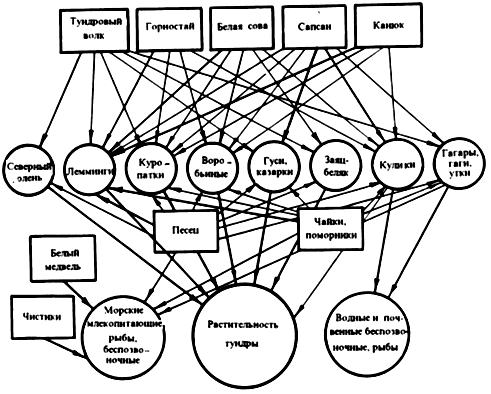
Рис. 2. Пищевые связи в биоценозе арктических тундр летом (по В.М. Сдобникову из Н.П. Наумова, 1963)
2. Экологическая пирамида
В любой цепи питания не вся пища используется на рост особи, т.е. на накопление биомассы. Часть ее расходуется на удовлетворение энергетических затрат организма: на дыхание, движение, размножение, поддержание температуры тела. При этом биомасса одного звена не может быть переработана последующим полностью. В противном случае исчезли бы ресурсы для развития живой материи. В каждом последующем звене пищевой цепи происходит уменьшение биомассы. Обычно, чем больше масса начального звена, тем больше она в последующих звеньях. Это касается не только биомассы, но и численности особей, и запаса энергии.
Данное явление было изучено Ч. Элтоном и названо пирамидой чисел или пирамидой Элтона. Различают пирамиду численности (особей), пирамиду биомассы и пирамиду энергии.
Основание пирамиды образуют растения-продуценты. Над ними располагаются фитофаги. Следующее звено представлено консументами второго порядка. И так далее до вершины пирамиды, которую занимают наиболее крупные хищники. Высота пирамиды обычно соответствует длине пищевой цепи. И поскольку на верхние этажи пирамиды энергия доходит в очень малых количествах, цепь редко состоит более чем из 5—6 звеньев.
Ю. Одум сделал расчеты потока энергии от звена к звену в упрощенной теоретической экосистеме, сведя ее к одной примитивной цепи, функционирующей в течение года. Он рассуждал следующим образом. Допустим, имеется посев люцерны на площади в 4 га. На этом поле кормятся телята (предполагается, что они едят только люцерну), а телятиной питается 12-летний мальчик. Результаты расчетов, представленные в виде трех пирамид — численности, биомассы и энергии, свидетельствуют, что люцерна использует всего 0,24 % всей падающей на поле солнечной энергии, из которой 8 % приходится на телят; 0,7 % энергии, накопленной телятами, расходуется на развитие и рост ребенка с 12 до 13 лет. Несмотря на то что рассматриваемая схема искусственна, она все же дает четкое представление о масштабах снижения коэффициента полезного действия по мере перехода от основного звена в пирамиде кее вершине: из всей солнечной энергии, падающей на 4 га люцернового поля, лишь немногим больше миллионной частиее хватает на пропитание мальчика в течение года.
Из трех типов экологических пирамид пирамида энергии дает наиболее полное представление о функциональной организованности сообществ, потому что количество и масса организмов зависят не от количества фиксированной энергии в данный момент на предыдущем уровне, а от скорости продуцирования пищи. Пирамида энергии отражает картину скоростей прохождения массы пищи через пищевую цепь.
Правило пирамиды чисел универсально и объективно отражает круговорот веществ и поток энергии в биосфере. В масштабе всей биосферы это правило никогда не нарушается.
Правда, на незначительных участках могут быть некоторые отклонения от него. Это имеет место при вспышках массового размножения вредителей, когда полностью уничтожается растительность и на какой-то ограниченной территории временно разрушается цепь питания. В данном случае в движение приходит все сообщество животных и растений, связанных между собой пищевыми отношениями.
3. Поток веществ и энергии
Жизнь, возникнув на Земле, вот уже на протяжении миллиардов лет находится в постоянном развитии. Это происходит благодаря тому, что элементы живого вещества, поступающие из окружающей среды, пройдя через ряд организмов, снова возвращаются во внешнюю среду, а затем опять включаются в состав живого вещества. Таким образом, каждый элемент используется живой материей многократно. Именно круговоротом веществ и обусловлено неограниченное временем существование и постоянное развитие и совершенствование жизни на Земле. Этот так называемый биогенный круговорот веществ — важнейшая функция любого биогеоценоза. Его характер определяют изменения массы живых организмов (биомассы), структуры биогеоценоза, химизма среды. Однако биогенный круговорот веществ не следует понимать в абсолютном смысле. Как бы там ни было, эти вещества, переходя с одного трофического уровня на другой, высвобождаясь и вновь включаясь в состав живого вещества, частично исключаются из круговорота. В результате на Земле происходит накопление органических соединений в виде залежей полезных ископаемых (торф, уголь, нефть, газ, горючие сланцы). Но все это не отвергает общего правила. Существенно биомасса на Земле не накапливается, а удерживается на каком-то определенном уровне, поскольку она постоянно разрушается и вновь созидается из одного и того же строительного материала, т. е. в ее пределах протекает беспрерывный круговорот веществ.
Биогенный круговорот веществ принял определенный характер с появлением зеленых растений, осуществляющих процессы фотосинтеза. Рассмотрим это на примере круговорота кислорода — продукта фотосинтеза растений. Практически весь молекулярный кислород земной атмосферы возник и поддерживается на определенном уровне благодаря деятельности зеленых растений. В большом количестве он расходуется организмами в процессе дыхания. Но, кроме того, обладая высокой химической активностью, кислород непрерывно вступает в соединения почти со всеми элементами земной коры. Если бы зеленые растения не выделяли такого огромного количества кислорода, он бы в конце концов полностью исчез из атмосферы, и тогда преобразился бы весь облик Земли: исчезли бы почти все организмы, прекратились бы все окислительные процессы — планета наша стала бы безжизненной. Однако это ей не угрожает именно потому, что в природе происходит нескончаемый круговорот веществ. Подсчитано, что весь кислород, содержащийся в атмосфере, оборачивается через организмы (связываясь при дыхании и высвобождаясь при фотосинтезе) за 2000 лет, углекислота атмосферы совершает круговорот в обратном направлении за 300 лет, а все воды на Земле разлагаются и воссоздаются путем фотосинтеза и дыхания за 2 000 000 лет.
Однако для столь грандиозного биологического круговорота веществ необходима энергия. Источником ее является солнечная радиация, аккумулируемая зелеными растениями-автотрофами. Солнечная энергия также регулярно циркулирует в биогеоценозе. Но в отличие от круговорота веществ, который протекает по замкнутому кругу, переходя в цепях питания с одного трофического уровня на другой, энергия постоянно расходуется. До 30 % ее рассеивается в атмосфере или отражается облаками и поверхностью Земли, до 20 % поглощается в верхних слоях атмосферы (водяные пары, капельки воды, пылевые частицы), приблизительно 50 % достигает суши и поверхности океана и поглощается в форме теплоты. Лишь ничтожная часть, всего около 0,1—0,2 % энергии, получаемой Землей от Солнца, улавливается зелеными растениями и обеспечивает весь биологический круговорот веществ в биосфере.
Более половины энергии, связанной при фотосинтезе, тут же расходуется на дыхание растений, а остальная поступает в пищевые цепи.
Суммарно только около 1 % лучистой энергии Солнца, которая падает на растение, превращается в потенциальную энергию химических связей синтезированных органических веществ. Более половины этой энергии расходуется на жизнь самих растений, а остальная поступает в пищевые цепи и может быть использована гетеротрофными организмами при питании. Когда животное съедает растение, большая часть энергии, которая содержится в пище, используется на различные процессы жизнедеятельности, превращаясь при этом в теплоту и рассеиваясь в пространстве. Только 5—20 % энергии пищи переходит во вновь созданное живое вещество тела животного. Если растительноядное животное съедается хищником, то вновь теряется большая часть заключенной в пище энергии. В результате таких огромных потерь полезной энергии цепи питания не могут быть очень длинными.
Таким образом, энергия Солнца, утилизированная зелеными растениями, превращается в потенциальную энергию химических связей органических соединений, из которых строится тело самих растений. В организме растительноядного животного эти органические вещества окисляются с выделением такого количества энергии, которое было затрачено на их синтез растением. Часть ее используется для жизни животного, а остальная, согласно второму закону термодинамики (переход энергии из одной формы в другую сопровождается снижением количества полезной энергии), превращается в теплоту и рассеивается в пространстве, т.е. уходит из биоценоза (энтропия).
Поток энергии в экосистеме может быть проиллюстрирован схемой простой цепи питания (рис. 3). Солнечная энергия, полученная растением, лишь частично используется в процессе фотосинтеза углеводов.
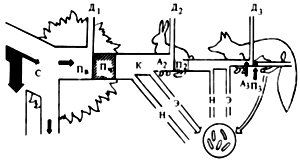
Рис. 3. Поток энергии через три уровня простой пищевой цепи (по П. Дювиньо, 1973)
Фиксированная в углеводах энергия представляет собой валовую продукцию биогеоценоза (Пв). Углеводы идут на построение протоплазмы и рост растений, причем часть их энергии затрачивается на дыхание (Д1). В результате чистая продукция (Пч) определяется по формуле
Пч = Пв - Д1.
Таким образом, поток энергии, проходящий через уровень продуцентов, т.е. валовую продукцию, можно представить так:
Пв = Пч + Д1
Определенный объем созданных продуцентами веществ служит кормом (К) фитофагов, остальные в конце концов отмирают и перерабатываются редуцентами (Н). Корм, ассимилированный фитофагами (А2), лишь частично используется для образования их биомассы (П2).
В основном он растрачивается на обеспечение энергией процессов дыхания (Д2) и в некоторой степени выводится из организма в виде выделений и экскрементов (Э). Поток энергии, проходящий через второй трофический уровень, выражается следующим образом:
А2 = П2 + Д2.
Консументы второго порядка (хищники) не истребляют всю биомассу своих жертв, но и из того количества ее, которое они уничтожают, лишь часть используется на создание биомассы их собственного трофического уровня. Остальная же часть в основном затрачивается на энергию дыхания, а также выделяется с экскретами и экскрементами. Поток энергии, проходящий через уровень консументов второго порядка (плотоядные), выражается формулой:
Аз = Пз + Дз.
Анализируемая схема наглядно показывает, что поток энергии, который выражается количеством ассимилированного по цепи питания вещества, на каждом трофическом уровне уменьшается, т.е. Пч>П2>Пз и т. д.
Таким образом, поскольку определенное количество вещества может быть использовано каждым биоценозом неоднократно, а порция энергии — лишь один раз, в экосистеме осуществляется не «круговорот веществ и энергии», как иногда указывается, а каскадный перенос (поток) энергии (рис. 4) и круговорот веществ, т.е. применение понятия «круговорот» правомерно только по отношению к веществам.
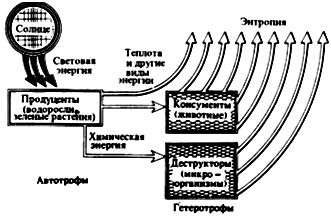
Рис. 4. Поток энергии в биосфере (по Ф. Рамаду, 1981)
Этот процесс протекает в природе с определенной скоростью. Поэтому биологическую продуктивность можно выразить продукцией за сезон, за год, за несколько пет или за любую другую единицу времени. Для наземных и донных организмов она определяется количеством биомассы на единицу площади, а для планктонных и почвенных — на единицу объема.
Следовательно, биологическая продуктивность представляет собой количество воспроизведенной биомассы на 1 м2
площади (или на 1 м3
объема) в единицу времени и выражается чаще всего в граммах углерода или сухого органического вещества. Биологическую продуктивность нельзя смешивать с биомассой. Допустим, за год планктонные водоросли на единицу площади синтезируют столько же органического вещества, сколько и высокопродуктивные леса, однако биомасса последних в сотни тысяч раз больше.
Биомасса того или иного биоценоза не дает четкого представления о его продуктивности. Это связано с тем, что скорость образования биомассы (продуктивность) в разных биоценозах неодинакова. Поэтому биоценозы различаются не только биомассой, но и продуктивностью, т. е. скоростью создания определенного количества биомассы. Луговые степи дают больший годовой прирост биомассы, чем хвойные леса. При средней фитомассе 23 т/га годовая продукция их (оставляет 10 т/га, тогда как у хвойных лесов при фитомассе 200 т/га она равна 6 т/га. Популяции мелких млекопитающих по сравнению с крупными обладают большой скоростью роста и размножения и дают более высокую продукцию при равной биомассе.
Таким образом, чтобы оценить значение вида (группы видов) для круговорота веществ и в отношении его биологической продуктивности в биоценозе или в биогеоценозе в целом, нужно знать не только его биомассу, но и относительную скорость прироста или время ее полного возобновления.
Продукция каждой популяции за определенное время представляет собой сумму прироста всех особей, включая отделившиеся от организма образования и устраненные (элиминация) по разным причинам особи (смерть, миграция).
В том случае, когда все особи доживают до конца изучаемого периода, продукция равна приросту биомассы. В природе это исключено, и продукция популяции обычно рассчитывается по формуле:
Р = (В2 – В1) + Е,
где Р — продукция; В1 и В2 — соответственно начальная и конечная биомасса; Е — элиминация.
Это так называемая чистая продукция. Валовая продукция включает в себя прирост (чистая продукция) и затраты на энергетический обмен.
Необходимо различать первичную продукцию, т.е. продукцию автотрофных организмов, и первичную продуктивность, т.е. скорость, с которой автотрофные организмы (продуценты) в процессе фотосинтеза связывают энергию и запасаютее в форме органического вещества.
Подсчитано, что солнечная энергия, достигающая поверхности Земли в течение года, исчисляется в 5-1020
ккал (21 •1020
кДж). Это составляет 9 млрд. ккал (37,8-109
кДж) на гектар. Один гектар леса в средних широтах продуцирует до 6 т древесины и 4 т листьев, сжигание которых дает 46 млн. ккал (193,2-106
кДж). Значит, эффективность первичной продуктивности леса, т. е. эффективность использования растениями солнечной энергии для создания органического вещества, составляет всего около 0,5% (46х100:9). Конечно, цифры эти чрезвычайно относительны, поскольку эффективность первичной продуктивности зависит от возраста леса, количества деревьев, погодных условий и многих других факторов. Но тем не менее они дают представление о коэффициенте полезного действия биоценоза.
Консументы образуют свою биомассу. Для обозначения биомассы и скоростиее образования консументами применяются термины «вторичная продукция», т. е. продукция гетеротрофных организмов, и «вторичная продуктивность», т. е. скорость образования продукции гетеротрофами. Как уже отмечалось, поток энергии от продуцентов к консументам сопровождается потерямиее.
Дело в том, что значительная часть съеденного гетеротрофами корма расходуется на теплопродукцию, на выработку энергии, необходимой для их жизнедеятельности, и лишь небольшое количество его (1,3—2%) используется на создание вторичной продукции. Например, для получения 1 кг говядины требуется от 70 до 90 кг свежей травы.
При этом необходимо учитывать, что все виды, дающие вторичную продукцию, возникают на основе утилизации вещества и энергии первичной продукции. Но так как при переходе с одного трофического уровня на другой энергия частично затрачивается на нужды энергетического обмена и рассеивается, то продукция каждого последующего трофического уровня меньше продукции предыдущего. Например. продукция (ее выход) фитофагов всегда больше, чем у живущих за их счет хищников.
Большое значение в механизме биологического продуцирования имеют гетеротрофные организмы, утилизирующие поступающее со всех трофических уровней мертвое органическое вещество, частично минерализуя его, частично превращая в вещество микробных тел. Последнее служит важным источником питания многих водных и почвенных животных.
Кроме первичной и вторичной продукции биоценозов, различают промежуточную и конечную продукции. Промежуточная продукция отличается тем, что после потребления другими членами биогеоценоза возвращается в круговорот веществ этой же системы. Конечная продукция исключается из данного биогеоценоза, т. е. выводится за его пределы. Это, к примеру, продукция, получаемая человеком в процессе возделывания сельскохозяйственных культур, разведения домашних животных, охоты, промысла и т. д.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Радкевич В. А . Экология: Учебник. -4-е изд., стер. –Мн.: Выш. шк., 1998. -159 с.: ил.
2. Одум Ю. Экология. –М., 1986. –Т. 1. 328 с.
3. Чернова Н.М., Былова А.М. Экология. –М., 1988. -272 с.
|