КУРСОВАЯ РАБОТА НА ТЕМУ:
ОБМЕН ВЕЩЕСТВ У РЫБ
План
Вступление…………………………………………………………………..3
Структурные элементы питания рыб…………………………………..….4
Взаимосвязь обмена веществ рыб и химического состава воды……….11
Поддержание солевого баланса и система осмотической регуляции
у рыб………………………………………………………………………..16
Зависимость обмена веществ у рыб от температуры воды……………..18
Влияние растворенных в воде газов на обмен веществ у рыб……….....24
Выводы……………………………………………………………………..28
Список литературы………………………………………………………..29
Вступление
Обмен веществ, или метаболизм, — лежащий в основе жизни, закономерный порядок превращения веществ и энергии в живых системах, направленный на их сохранение и самовоспроизведение; совокупность всех химических реакций, протекающих в организме.
Обмен веществ складывался при самом возникновении жизни на Земле, поэтому в его основе лежит единый для всех организмов нашей планеты биохимический план. Однако в процессе развития живой материи изменения и совершенствование шли неодинаковыми путями у разных представителей животного и растительного мира. Поэтому организмы, принадлежащие к различным систематическим группам и стоящие на разных ступенях исторического развития, наряду с принципиальным сходством в основном порядке химических превращений, имеют существенные и характерные отличия.
Жизнедеятельность, обмен веществ рыб неразрывно связаны с водой. Их организм приспособился не только к водной среде, но и к ее физико-химическим характеристикам. Для того чтобы рыбы нормально развивались, их метаболизм должен быть настроен на составляющие среды. Хотя некоторые виды рыб умеют адаптироваться к непривычным для них параметрам воды, это отразится на них в будущем, а различия между соленой и пресной водой столь значительны, что вообще не могут быть преодолены [10,11].
1.
Структурные элементы питания рыб
Потребность рыбы в структурных элементах питания не остается постоянной. Она изменяется в зависимости от возраста, размера, половой зрелости рыб, гидрохимических свойств и температуры воды.
Белки (Протеины, Proteins) - природные высокомолекулярные органические соединения.
В процессах жизнедеятельности всех организмов белки выполняют структурную, регуляторную, каталитическую, защитную, транспортную, энергетическую и другие функции. В зависимости от формы белковой молекулы различают фибриллярные и глобулярные белки [1].
Белковые молекулы представляют собой длинные сложные цепочки, состоящие преимущественно из аминокислот (Рис.1.1). Основных аминокислот всего 20 штук. Каждый вид живых существ обладает собственным, только ему присущим набором разновидностей белковых молекул [3,17]. В процессе пищеварения белковые молекулы расщепляются именно на аминокислоты, которые всасываются в кровь и переносятся ею к клеткам организма как детали на сборочный конвейер. Часть аминокислот животные могут синтезировать самостоятельно. Главным образом этот синтез происходит в печени [9]. Однако некоторые из них могут быть получены только с пищей. Такие аминокислоты называют незаменимыми. У разных видов рыб их от 9 до 12.

R- атом водорода или какая-нибудь органическая группа
Рис. 1.1 Общая формула аминокислот
Рыбы, в т.ч. растительноядные, нуждаются в существенно большем (в 2-3 раза) содержании белка в корме, чем наземные животные. Это связано с особенностями их обмена веществ. Больше половины белка у рыб расходуется на энергетический обмен. Наземным животным с их громоздкой системой мочевыделения в этом случае грозило бы отравление, а рыбы достаточно легко справляются с этой проблемой, выводя аммиак через жабры [13,14].
Обычно оптимальной концентрацией белка в сухом веществе (т.е. без учета воды) корма для рыб считается 30-40%. Плотоядным рыбам нужно больше белка, чем растительноядным. Морским рыбам, в среднем, - больше чем пресноводным. Активно растущей молоди – больше чем взрослым рыбам.
Липиды (Lipids) – греч. lipos жир + eidos вид – класс жиров и жироподобных веществ (липоидов). С химической точки зрения представляют собой жирные кислоты и их производные [3].
Молекулы липидов состоят в основном из атомов углерода, водорода и кислорода. Кроме того, в небольшом количестве липиды, входящие в состав кормов, могут содержать и другие элементы - фосфор (фосфолипиды), азот.
Липиды играют весьма важную роль в жизнедеятельности организмов. Это: главные компоненты биомембран [5]; запасной, изолирующий и защищающий органы материал; наиболее калорийная часть пищи; важная составная часть диеты животных; переносчики ряда витаминов; регуляторы транспорта воды и солей; иммуномодуляторы; регуляторы активности некоторых ферментов; эндогормоны; передатчики биологических сигналов [3].
Жиры (Fats) – самая массовая разновидностей липидов корма (Рис.1.2). Кормовые жиры представлены в основном нейтральными жирами (триглицеридами). Это сравнительно простые соединения, которые в процессе пищеварения распадаются на составные части – глицерин и жирные кислоты. В составе триглицеридов содержится около 9% глицерина и жирные кислоты с разной длиной углеродной цепочки. Свойства триглицеридов зависят от длины и особенностей химической структуры, входящих в их состав жирных кислот [3].
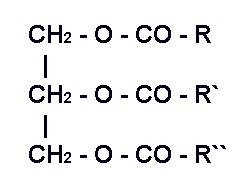
R, R’ и R’’ – углеводородные остатки (радикалы) жирных кислот, содержащие от 4 до 26 атомов углерода.
Рис.1.2 Общая формула жиров
Жиры являются основным источником энергии для большинства животных. Один грамм жира при полном окислении (оно идет в клетках с участием кислорода) дает 9,5 ккал (около 40 кДж) энергии. Это почти вдвое больше, чем можно получить из белков или углеводов. Кроме того, жировые запасы в организме практически не содержат воду, тогда как молекулы белков и углеводов всегда окружены молекулами воды. В результате один грамм жира дает почти в 6 раз больше энергии, чем один грамм животного крахмала – гликогена [16].
С другой стороны, жиры это не только высококалорийное «топливо». Они входят в состав клеточных компонентов, в том числе мембран, и служат основой синтеза важных для организма соединений. Жирорастворимые витамины (A, D, E и K) «хранятся» только в жирах и без них не усваиваются. При отсутствии в корме жира, нарушается деятельность центральной нервной системы, ослабляется иммунитет.
Несмотря на высокую «энергоемкость» жиров, получение из них энергии в организме – процесс медленный. Это связано с малой реакционной способностью жиров, особенно их углеводородных цепей. Углеводы, хотя и дают меньше энергии, чем жиры, зато позволяют получить ее намного быстрее. Жиры корма расщепляются в желудке и в кишечнике, после чего проникают через их стенки в кровеносные сосуды, откуда транспортируются в печень и жировые ткани, где происходит их накопление [2].
Жирные кислоты бывают насыщенными (предельными) и ненасыщенными (непредельными) [3]. Ненасыщенные жиры, также как незаменимые аминокислоты, не могут синтезироваться в организме рыб и должны поступать с кормом. Ткани высших позвоночных животных (в т.ч. с-х животных) содержат в основном насыщенные жиры, тогда как у рыб и растений они преимущественно ненасыщенные. Ненасыщенные жиры – мягкие и не застывают при пониженных температурах, характерных для рыб, которые имеют температуру окружающей среды. Жиры теплокровных животных – в основном твердые.
Если жир переходит из жидкого состояния в твердое, он теряет жирорастворимые витамины (A, D, E, K), которые затем быстро разрушаются. Жиры, находящиеся в твердом состоянии усваиваются в желудочно-кишечном тракте рыб гораздо хуже, чем жидкие. А «подогреть» пищу, как теплокровные животные, они не могут. Жиры, входящие в состав кормов для холодноводных рыб, должны застывать при более низкой температуре [6,7].
Углеводы (Сахара, Carbohydrates) - органические соединения, в состав которых входят углерод, кислород и водород. Имеют общую формулу Cn(H2O)m, за что и получили свое основное название.
В растениях углеводы - это первичные продукты фотосинтеза и основные исходные продукты биосинтеза других веществ. Они составляют существенную часть рациона многих животных; подвергаясь окислительным превращениям (биохимическому «сжиганию»), обеспечивают все живые клетки энергией; входят в состав клеточных оболочек и других структур [3].
Углеводы подразделяются на моносахариды (глюкоза, фруктоза…), дисахариды (сахароза, мальтоза…) и полисахариды (целлюлоза, она же – клетчатка, а также крахмал и его аналог животного происхождения – гликоген, …).
Ту часть полученных с кормом и усвоенных углеводов, которая сразу не была «сожжена» для получения энергии, водные животные преобразуют в жиры и гликоген, и накапливают в печени и мышцах. В случае необходимости гликоген легко превращается в глюкозу, а та, в свою очередь, может участвовать в энергетическом обмене.
Теплокровные животные тратят значительную часть полученных из пищи углеводов на поддержание повышенной температуры тела. Холоднокровные рыбы в этом не нуждаются, поэтому содержание углеводов не должно превышать 20-25% для молоди и 30-35% для взрослых рыб (данные для товарного рыбоводства). Считается, что рыбам не следует скармливать более 6 г углеводов на один килограмм их веса [8,17].
Хищные рыбы могут легко усваивать только низшие углеводы или гликоген. Растительноядные, с помощью особых ферментов, могут усваивать и высшие углеводы, например крахмал. Клетчатка также может усваиваться некоторыми видами рыб, видимо, при помощи микрофлоры желудочно-кишечного тракта. Однако для большинства видов содержание клетчатки не должно превышать 3%.
Виды рыб с высокой двигательной активностью (например, пелагические) должны получать больше углеводов, чем малоактивные. Быстро растущая молодь больше нуждается в углеводах, чем взрослые. При нересте и подготовке к нему потребность в углеводах также может возрастать [8].
Минеральные вещества - необходимы для нормальной жизнедеятельности рыб. Минеральные вещества в виде солей поступают в организм не только с пищей, но и из воды через жабры, слизистые покровы ротовой полости и кожу. Содержание в корме минеральных веществ становится менее важным, если рыба живет в соленой воде с высокой ионной активностью.
Витамины - незаменимые для жизни органические вещества разнообразной структуры, выполняющие функции биокатализаторов процессов, протекающих в живой клетке, и участвующие в обмене веществ.
В отличие от других незаменимых факторов питания (аминокислоты, жирные кислоты и др.) витамины не являются материалом для биосинтезов или источником энергии. Однако они участвуют практически во всех биохимических и физиологических процессах, составляющих в совокупности обмен веществ [3].
Биосинтез витаминов осуществляется в основном вне организма животного, поэтому животное должно получать витамины извне, с пищей.
Различают водорастворимые (C, B1, B2, B6, B12 и PP) и жирорастворимые витамины (A, D, E, K). Кроме того, выделяют группу витаминоподобных соединений.
К водорастворимым относятся: аскорбиновая кислота (витамин C), витамины группы B – тиамин (B1), рибофлавин (B2), B6, B12 (кобаламин), ниацин (витамин PP), фолацин, пантотеновая кислота, биотин.
К жирорастворимым относятся: витамины A, D (кальциферолы), E (токоферолы) и K.
Успешный перевод рыбы с естественной пищи на комбикорма стал возможным только после изучения потребности рыбы в витаминах [13].
2. Взаимосвязь обмена веществ и химического состава воды
Обмен веществ между организмом и окружающей средой тесно связан с химическим составом воды.
Синтез веществ, процессы дыхания, разложения сложных соединений проходят в клетках живых организмов. В процессе жизнедеятельности постоянно расходуются одни вещества и образуются другие. Часть вновь образовавшихся молекул остается в клетке, часть транспортируется в другие клетки или выводится в окружающую среду. Для обеспечения процесса жизнедеятельности необходим постоянный подвод исходных составляющих и отвод из клетки побочных продуктов, образовавшихся в ходе биохимических реакций [5].
Транспорт молекул осуществляется по специально организованным передающим тканям. Перед тем, как попасть в клетку или выйти из нее, все вещества должны пройти через клеточную мембрану, отделяющую клетку от внешней среды. Процессы обмена веществ на мембранах тесно связаны с химическим составом воды [5,13]. Содержание различных солей оказывает влияние на то, какие вещества и в каких количествах будут поступать в клетку или выходить из нее. Продукты, необходимые для жизнедеятельности организма, обычно транспортируются через мембрану в виде заряженных ионов. Транспорт может осуществляться активно — с использованием богатых энергией соединений или пассивно, за счет собственной кинетической энергии ионов. Пассивный транспорт — диффузия различных ионов через мембрану — осуществляется с разной скоростью. Относительная способность разных ионов диффундировать через мембрану определяет коэффициент проницаемости Р. Легче других проникает через мембраны ион К+, поэтому значение Р для К+ условно принимают за 1,0. Скорость проникновения ионов через мембрану зависит также от разности концентраций данного иона по обе стороны мембраны. Чем больше разность концентраций, тем больше ионов диффундирует в сторону меньшего их содержания. Кроме диффузии, идущей за счет разницы концентраций, существует активный транспорт ионов, при котором движение осуществляется за счет разности электрохимических потенциалов через специальные участки мембраны. Это движение может осуществляться и от меньшей концентрации к большей. Движущей силой процесса в этом случае является запас энергии в форме молекул АТФ [5,13].
Упрощенно структура живой клетки выглядит следующим образом: внутри клеточной стенки (сравнительно жесткого образования) располагается протопласт (живая часть клетки), в котором заключены все клеточные организмы, находящиеся в сложном растворе — цитоплазме [3]. Клеточная стенка имеет избирательную проницаемость для различных ионов, то есть различные вещества проникают сквозь мембрану с разными скоростями. Это определяется их различной растворимостью отдельных составляющих мембраны и различными скоростями перекачивания при активном транспорте. В результате образуется неравномерное распределение ряда веществ по обе стороны мембраны. Клетки растений активно накачивают калий, а близкий к нему натрий, наоборот, выталкивается в окружающую среду [3,5]. Из-за более высоких концентраций некоторых ионов внутри клетки создается осмотическое (диффузное) давление, характеризующее стремление раствора, отторгнутого мембраной, к снижению концентрации (разбавлению). Осмотическое давление может достичь десятков атмосфер. Это давление создает напряженное состояние клеточной оболочки. Напряжение мембраны зависит также от внешнего раствора. В зависимости от отношения осмотического давления внешнего раствора к давлению в клетке растворы подразделяются на три группы. Изотонические — в них разница давлений невелика (менее 0,5—1,0 атм); гипертонические — их давление выше, чем в клетке; противоположные им — гипотонические. Если клетка находится в гипертоническом растворе, то из нее происходит откачка воды, что приводит к уменьшению размера клетки и сжатию мембраны [3,5].Из гипотонических растворов вода поступает в клетки, что приводит к их набуханию (вплоть до разрыва мембраны) и потере части активных веществ.
Совокупность процессов регулирования осмотического давления жидкостей организма носит название осморегуляция [3]. Этот процесс обнаружен у большинства организмов. У пресноводных рыб вода вместе с содержащимися в ней солями активно поступает в клетки через поверхность тела и жабры и выводится из организма через почки. У солоноводных рыб попавшая в организм вода выводится через кожные покровы, a NaCl выводится главным образом через жабры за счет специальных желез [9,10]. Водные растения и пресноводные рыбы удовлетворяют потребность организма в ионах, поглощая их непосредственно из воды. Если она не содержит необходимые элементы, то при нормальном соотношении осмотических давлений происходит изменение содержания отдельных элементов, то есть изменение отношения ионов в организме. В ряде случаев это приводит к нарушению биохимических процессов [14].
В ходе экспериментов с пресноводными рыбами обнаружено, что они неплохо переносят изотонические растворы, полученные разбавлением морской воды, в то время как гипотонические растворы одной из солей — калия, магния, натрия или кальция — действовали смертельно. Был получен ряд токсичности ионов основных металлов:
Na+ < Ca2+, Mg2+ < K+
Опыты показали, что воздействие на рыб оказывает содержание Na+ в крови [4].
При повышении концентрации Na+ в воде соответственно увеличивается его содержание в крови, а содержание К+ уменьшается. При повышении концентрации калия происходит обогащение организма натрием. Так что калий оказывает косвенное токсическое воздействие. Обогащение рыб натрием дифференцировано в зависимости от пола рыбы. Кровь самок быстрее обогащается натрием (возможно за счет реакции яичников) [14,17].
При поглощении Na+ требуется большое количество энергии. При ассимиляции Na+ организмом происходит его замещение на NH4+. Аммоний может выделяться организмом из органических азотсодержащих соединений. Таким образом, повышенное потребление натрия приводит к нарушению белкового обмена [1]. У растений повышение концентрации натрия приводит к блокаде поступления ионов калия через мембраны клеток. Растение может испытывать калийное голодание даже при достаточно высоком абсолютном содержании калия [3].
Анионы также имеют различное воздействие на обитателей. Так, нитраты для рыб значительно более ядовиты, чем хлориды. Для растений наиболее токсичны хлорид-ионы Cl‾, затем следуют сульфат- и карбонат-ионы (SO42‾ и СO32‾).
Кроме осмотического давления и абсолютного содержания того или иного иона в воде большое физиологическое значение имеет соотношение ионов, растворенных в воде [5]. Большинство природных вод имеет приблизительно равное суммарное содержание одновалентных и двухвалентных ионов. К такому соотношению приспособлены процессы жизнедеятельности водных организмов. Конечно, в различных регионах земного шара состав воды различен, но организмы имеют возможность приспосабливаться к некоторым изменениям химического состава.
3. Поддержание солевого баланса и система осмотической регуляции у рыб
Жидкости, присутствующие в теле рыбы, содержат различные соли. Чтобы у рыбы эффективно происходил обмен веществ, концентрация этих солей должна оставаться в узком диапазоне. У пресноводных рыб в мышечной жидкости содержится большее количество солей, чем в окружающей воде. Для морских рыб наоборот - внутри у них меньше соли, чем в морской воде. Если бы рыбы были полностью водонепроницаемыми, они могли бы поддерживать свой внутренний водно-солевой баланс без затрат энергии [2,11].
Однако на самом деле рыбы "протекают" - в том смысле, что вода и соли могут проникать через тонкие эпителиальные поверхности, особенно через жабры. Вода проникает через жабры пресноводных рыб в процессе осмоса, а соли выходят через жабры путем естественной диффузии. Поэтому рыбы должны затрачивать энергию, чтобы противостоять этим силам. Именно это они и делают в процессе, который получил название осмотической регуляции [13,14].
Осмотическая регуляция у пресноводных рыб осуществляется путем сочетания физиологических процессов, которые происходят главным образом в почках и жабрах. Функция почек состоит в том, чтобы выводить из тела избыток воды [9]. Это достигается благодаря специальным трубчатым структурам внутри ткани почек, которые отфильтровывают воду из крови и выводят ее в мочевой пузырь, откуда она испускается в виде мочи. Мочевой пузырь имеется не у всех групп рыб и его не следует путать с плавательным пузырем. При одинаковом весе тела пресноводные рыбы производят примерно в 10 раз больше мочи, чем морские (и, соответственно, примерно в 10- 20 раз больше, чем наземные животные).
Помимо того, что рыбы вынуждены справляться с избыточным притоком воды в организм, они еще должны сохранять соли, присутствующие в их теле. Необходимое пропорциональное количество солей в моче поглощается почками еще до испускания мочи. Кроме того, есть еще специальные клетки в жабрах - хлоридовые клетки, которые также помогают поддерживать солевое соотношение путем активного поглощения солей (в виде ионов) непосредственно из воды. Эта система поглощения солей, требующая затрат энергии, называется "ионным насосом". Этот процесс работает в обоих направлениях, и нежелательные ионы (такие, как ионы аммиака NH4+) обмениваются на полезные ионы (например, ионы натрия Na+) [5,14,17].
4. Зависимость обмена веществ у рыб от температуры воды
Температура водной среды - самый значительный природный фактор, который прямо воздействует на уровень обмена веществ пойкилотермных животных, к которым относятся и рыбы [12].
Всех рыб по диапазону температур, при котором возможна их нормальная жизнедеятельность, разделяют на теплолюбивых (плотва, сазан (карп), карась, линь, растительноядные виды (толстолобики, белый амур), осетровые и прочие) и холодолюбивых (ручьевая форель, сиги, лосось, налим и др.) [10,11,16].
Обмен веществ у первых представителей наиболее эффективен при высокой температуре. Они наиболее интенсивно питаются и активны при температуре +17-28°С, при понижении температуры воды до +17°С их пищевая активность ослабевает (а зимой у многих видов вообще прекращается). Предзимье и всю зиму они проводят в малоподвижном состоянии в глубоких местах водоема [12].
Для холодолюбивых рыб оптимальные температуры +8-1б°С. Зимой они активно питаются, а их нерест происходит в осенне-зимний период. Известно, что к похолоданию и снижению температуры воды рыба "привыкает", перестраивая свой метаболизм только за 17-20 суток. При снижении температуры воды с +12°С до +4°С у хариуса, например, величины энергозатрат уменьшаются на 20% [12].
С понижением температуры воды увеличивается растворимость кислорода, поэтому зимой насыщенность воды кислородом достаточно высока.
При длительном понижении температуры воды рыбы должны располагать не только достаточным запасом жира как энергетического материала, но и в течение этого периода сохранить нормальный обмен веществ [6,12].
Способность приспосабливаться к меняющимся условиям среды - одна из важнейших особенностей живых существ. Их распространение, численность и биоразнообразие в значительной мере определяются эффективностью адаптационных механизмов. Именно они позволяют организмам существовать в условиях, часто малопригодных для жизни, а иногда несовместимых, на первый взгляд, с нею [12,13].
Механизмы температурных адаптаций [12]:
Биологические антифризы. Устойчивость к низким температурам холоднокровных животных полярных областей определяется несколькими механизмами. У обитателей этих районов в биологических жидкостях (крови и лимфе) присутствуют биологические антифризы - пептиды и гликопротеины, препятствующие замерзанию воды в клетках. Кроме того, устойчивость к холоду обеспечивают многоатомные спирты - глицерин и сорбит. Если пептиды и гликопротеины останавливают рост микрокристаллов льда в клетках, то глицерин заменяет воду, которая при понижении температуры выводится во внеклеточную среду, где лед менее опасен. Глицерин, в частности, способствует стабилизации структуры белков и биологических мембран. У большинства морских животных в снижении температуры замерзания жидкостей участвуют присутствующие в крови глюкоза, аминокислоты и соли (прежде всего NaCl). Чем больше этих молекул в растворе, тем меньше у воды возможностей формировать кристаллы льда. Такой механизм называется коллигативным (заместительным).
Высокомолекулярные антифризы адсорбируются на образовавшихся мельчайших (и поэтому не опасных) внутриклеточных кристаллах льда, не давая им расти. Это очень эффективные криопротекторы: так, гликопротеины тормозят замерзание жидкостей в 200 - 300 раз сильнее, чем вещества, обладающие коллигативным механизмом действия. Высокомолекулярные антифризы составляют значительную часть от веса всех жидкостей, и в их присутствии температура замерзания раствора ниже, чем температура таяния. У антарктических рыб антифризы постоянно присутствуют в жидкостях тела (3.5% от веса всех биологических жидкостей), что определяется генотипическими механизмами [12]. У других организмов они образуются в зависимости от сезонных колебаний температуры окружающей среды (фенотипическая адаптация). Синтез антифризов регулируется как экзогенно (при участии температуры среды и длительности светового дня), так и эндогенно (контролируется гипофизом).
Агрегатное состояние мембранных липидов. Интенсивность обмена веществ связана с клеточными мембранами, состояние которых зависит от мембранных липидов [5,13]. Степень их вязкости определяет работу мембранных ферментов, транспорт ионов, синаптическую передачу и другие процессы. Липидный бислой обычно находится в жидкокристаллическом состоянии. Но при понижении температуры мембраны “затвердевают”, и большинство мембранных процессов замедляется. Необходимая степень вязкости мембран при низких температурах поддерживается за счет изменения соотношения липидных компонентов мембран - насыщенных и ненасыщенных жирных кислот в сторону последних (они делают липидный бислой более рыхлым). Соотношение этих двух типов жирных кислот может меняться в ходе синтеза и включения в молекулы фосфолипидов, а также за счет изменения числа двойных связей. Кроме того, насыщенные жирные кислоты могут превращаться в ненасыщенные при помощи фермента десатуразы, который активируется изменением температуры. У рыб, обитающих в холодных водах, ненасыщенных жирных кислот больше, чем у жителей умеренных и тропических широт. Это позволяет метаболическим системам животных Арктики и Антарктики нормально функционировать [5,12,14].
Аминокислотные замены. Серьезная проблема для обмена веществ у холоднокровных животных при низких температурах - снижение конформационной гибкости белков, существенно затрудняющее их функции. Поддержание определенной гибкости молекул достигается благодаря мутациям, вызывающим аминокислотные замены в белках животных, которые обитают в высоких широтах. Характер таких замен детально изучен на примере фермента лактатдегидрогеназы [12]. При сравнении аминокислотных последовательностей этого белка, выделенного из скелетных мышц нототениевых рыб Антарктики и Южной Америки, было установлено, что замены локализованы в одной из α-спиралей, формирующих активный центр фермента. У большинства рыб умеренных широт в начале этой α-спирали находится остаток пролина, создающий определенную “жесткость” данному участку. У нототениевых рыб Антарктики пролин заменен на аланин, что обеспечило более гибкую структуру активному центру лактатдегидрогеназы, столь необходимую при пониженной температуре. Это только один из многих примеров генотипических адаптаций.
Компенсация энергетического обмена. Еще одно важное приспособление к холоду связано с компенсацией уровня энергетического метаболизма у холоднокровных. Так, в мышцах у полярных рыб концентрация митохондрий выше, чем у тех же видов или их близких родственников из умеренных или тропических широт [14]. Как следствие этих различий, интенсивность дыхания полярных организмов выше по сравнению с обитателями умеренного пояса при расчете на одинаковую температуру, обычно 20°С. Другой тип компенсации метаболизма - более низкий энергетический барьер ферментативных реакций в тканях полярных животных, чем обитателей умеренного пояса [17].
При сезонных или более кратковременных (несколько недель) колебаниях температуры адаптационные механизмы имеют фенотипическую природу, они формируются на протяжении одного поколения, обратимы и “не записаны” в геноме. В этих случаях холоднокровные животные вынуждены приспосабливать свой метаболизм к новому температурному режиму [12].
Один из механизмов таких фенотипических адаптаций - синтез индуцированных температурой изоформ тех или иных ферментов, которые больше приспособлены к новым условиям. Например, в мозге радужной форели, находящейся в течение трех недель при низкой температуре, синтезируется специфическая “холодовая” изоформа ацетилхолинэстеразы. Образование индуцированных температурой изоформ эстераз показано также у других видов рыб. Однако такая индукция достаточно редкое событие.
Другой механизм связан с изменением функциональных свойств ферментов без изменения их изоформ. Было обнаружено, что у рыб при адаптациях к низким и высоким температурам в течение нескольких недель меняются функциональные свойства ферментов. На примере лактатдегидрогеназы из скелетных мышц вьюна показано, что величина константы Михаэлиса (КМ), по которой можно судить о сродстве фермента с субстратом, зависит от ряда факторов, в том числе и от температуры: при оптимальной - КМ минимальна (т.е. фермент-субстратное сродство максимально), значит, фермент функционирует наиболее эффективно [12]. У рыб, адаптированных к холодной воде, фермент-субстратное сродство максимально при низкой температуре. У рыб, помещенных в теплую воду, в течение двух-трех недель оптимум фермента постепенно смещается в сторону высокой температуры. Это говорит о том, что за данный срок фермент перестраивается на работу в новых условиях [14].
5. Влияние растворенных в воде газов на обмен веществ
Вода как среда обитания рыб содержит растворенные газы, особенно кислород, азот и в небольшом количестве углекислый газ.
Все рыбы дышат растворенным в воде кислородом, поэтому содержание его в воде имеет для них решающее значение. Лишь немногие рыбы частично приспособились к дыханию атмосферным кислородом. К содержанию кислорода в воде рыбы относятся неодинаково. Как правило, пелагические рыбы, речные и холодолюбивые, более требовательны к содержанию кислорода, чем донные, озерные и теплолюбивые [7,10].
По количеству кислорода, необходимого для нормального развития рыб, их делят на несколько групп:
рыбы, нуждающиеся в очень большом содержании кислорода в воде (7—11 см3/л); при снижении его до 5 см3/л дыхание рыб делается практически невозможным (форели, сиги);
рыбы, которым также необходима высокая степень содержания кислорода в воде, однако существование их возможно и при содержании его 5—7 см3/л (хариусы);
рыбы, менее требовательные к содержанию кислорода и легко переносящие уменьшение количества его до 4 cмз/л (окунь, карп, плотва, щука);
рыбы, довольствующиеся еще меньшим содержанием кислорода и могущие жить в водоемах, где его количество уменьшается до 0,5—2,0 см3/л (линь, сазан, карась) [2,17].
Морские рыбы, как правило, более чувствительны к недостатку кислорода, чем пресноводные, и задыхаются при уменьшении его содержания до 60—70% нормального.
Количество растворенного в воде кислорода зависит от температуры, солености, ледового покрова, развития растительности, процессов распада органического вещества и др.
При повышении температуры и солености растворимость кислорода в воде уменьшается. Так, при 0°С и солености 0‰ в воде может раствориться 10,29 см3/л, а при 30° С — только 5,57 cм3/л кислорода [17].
Потребление кислорода рыбами зависит от их вида, возраста, подвижности, плотности посадки, физиологического состояния, а также температуры и солености воды.
При повышении температуры воды обмен веществ возрастает и потребность в кислороде (в см3 на 1 кг массы за 1 ч) увеличивается.
На потребление кислорода рыбами оказывает влияние и соленость воды. У пресноводных рыб, например, при небольшом увеличении солености обмен веществ возрастает, а при значительном замедляется, и потребление кислорода уменьшается.
У некоторых рыб чувствительность к кислороду обостряется на определенных стадиях развития [14].
Обычно молодь рыб более требовательна к содержанию кислорода, чем старшие возрастные группы. Так, личинки плотвы на 8-й день после вылупления гибнут в воде с содержанием кислорода ниже 3,45 см3/л, в то время как 49-дневные мальки выдерживают уменьшение его до 1 см3/л, а взрослые рыбы — даже до 0,6 см3/л.
Чем рыба подвижнее, тем она больше потребляет кислорода.
Интенсивность обмена веществ и потребление кислорода одиночными рыбами и в скоплениях неодинаковы [10,13]. При высокой плотности населения рыб потребление кислорода ими снижается. У рыб, залегающих на зимовку в ямы (карповые), потребление кислорода по сравнению с одиночными рыбами значительно уменьшается. У гольяна, помещенного в водоем, где до этого содержались рыбы, потребление кислорода снижается до такого же уровня, как и при групповом содержании.
Потребление кислорода изменяется в зависимости от физиологического состояния рыбы. Перед нерестом у некоторых рыб потребление кислорода повышается на 25—50% первоначального.
При плохом кислородном режиме интенсивность питания низкая и не увеличивается даже при обилии корма [13].
Для рыб неблагоприятен не только недостаток кислорода в воде, но и избыток его, причем в обоих случаях замедляются окислительные процессы. При быстром повышении содержания кислорода у рыб появляются беспокойство, одышка, кислородный наркоз, и они погибают от удушья.
Избыток кислорода в воде по сравнению с оптимальным режимом в период эмбрионального развития снижает функцию кроветворных органов, что вызывает анемию у рыб [6].
Снижение содержания кислорода может привести к летним и зимним заморам.
Зимние заморы обычно характерны для проточных и слабопроточных водоемов — озер, прудов, болот, богатых органическими веществами, на окисление которых расходуется большое количество кислорода, а поступление его из воздуха становится невозможным из-за ледового покрова. Зимние заморы возникают и на некоторых реках, длительное время покрытых льдом [15].
Летние заморы чаще всего происходят в заросших прудах и озерах ночью или в период массового развития в них водорослей, особенно часто ночью, когда происходит усиленное потребление кислорода растениями.
Растворенные в воде углекислый газ и сероводород отрицательно влияют на жизнедеятельность рыб [15,16].
Углекислый газ образуется в результате дыхания животных и растений, при разложении органических веществ. Наличие большого количества его является показателем загрязнения водоема. Даже при небольшом содержании углекислого газа в воде кровь теряет способность усваивать кислород, и рыба погибает от удушья, поэтому такие водоемы непригодны для рыб [4,15].
Сероводород появляется в водоеме при недостатке кислорода. Сероводород может образовываться и на дне пресных стоячих водоемов, и некоторые малоподвижные рыбы, такие, как линь, карп, карась и др., могут переносить небольшую концентрацию сероводорода.
Следует отметить, что иногда в некоторых участках рек с быстрым течением, особенно под водосливом гидроэлектростанций, вода перенасыщается газами воздуха, что может вызвать у рыб газопузырьковую болезнь.
Активная реакция среды (рН), имеющая важное значение для жизни рыб, зависит от соотношения растворенных в воде кислорода и свободной углекислоты и закономерно изменяется в зависимости от суточного и сезонного хода фотосинтеза.
В пресных водоемах избыток углекислого газа вызывает увеличение кислотности воды, в то время как в морской, содержащей в большом количестве бикарбонаты, избыток этого газа связывается, и рН более постоянна [16].
Для каждого вида рыб характерны определенные значения активной реакции среды. При изменении этих величин обмен веществ нарушается, так как снижается способность организма поглощать кислород. Оптимальная величина рН для рыб обычно составляет от 7 до 8 [2].
Выводы
В процессе жизнедеятельности потребность организма рыб в тех или иных веществах неодинакова и зависит от возраста, размера, половой зрелости рыб, гидрохимических свойств и температуры воды.
Процессы обмена веществ на мембранах клеток тесно связаны с химическим составом воды. Содержание различных солей оказывает влияние на то, какие вещества и в каких количествах будут поступать в клетку или выходить из нее.
Жидкости, присутствующие в теле рыбы, содержат различные соли. Чтобы у рыбы эффективно происходил обмен веществ, концентрация этих солей должна оставаться в узком диапазоне.
Совокупность процессов регулирования осмотического давления жидкостей организма носит название осморегуляция.
Температура водной среды и растворенные в воде газы - весомые факторы, влияющие на интенсивность обмена веществ у рыб.
Список литературы
Александров В.Я. Реактивность клеток и белки. - Л.: 1985 г.;
Баклашова Т. А. Ихтиология. – М.: 1980 – 122 стр.;
Гудин Т., Мерсер Э. Введение в биохимию растений. - М.: Мир, 1986- 312 с.
Карпевич А. Ф. Теория и практика акклиматизации водных организмов. - М: 1975 г.;
Клячко О.С. и др. // Биофизика. - 1993. -Т.28. - С.596 - 601.;
Никольский Г. В. Частная ихтиология. - 3 изд., М.:1971 г.;
Моисеев П. А., Азизова Н.А., Куранова И. И. Ихтиология. - М.: 1981 г.;
Моисеев П. А., Вавилкин А. С., Куранова И. И. Ихтиология и рыбоводство. - М.: 1975 г.;
Наумов Н.П., Kapташев Н. Н. Зоология позвоночных. - М.: 1979 г.;
Никольский Г. В. Экология рыб, 3 изд. - М.: 1974 г.;
Одум Ю. Экология: В 2 т. - М.: Мир, 1986 г.;
Озернюк Н.Д. Температурные адаптации. - М.: 2000 г.
Строганов Н. С. Экологическая физиология рыб. – М.: 1962.- Т.1.
Хочачка П., Сомеро Дж. Биохимическая адаптация. - М.: 1988 г.
Худолей В.В., Мизгирев И.В. Экологически опасные факторы. - СПб, 1996.
Шилов И.А. Экология. - М.: Высшая школа, 2003. -512 с.
Шустов С.Б., Шустова Л.В. Химические основы экологии.-М.: Просвещение, 1995.
|