Оптимальность как результат эволюции
Содержание
Введение
1. Оптимальное пищедобывательное поведение
2. Явления компромисса в поведении животных
3. Функции затрат
4. Функциональные аспекты процесса принятия решения
5. Бюджет времени и бюджет энергии
Многие животные и растения кажутся идеально "сконструированными" для выполнения своих функций или достижения своих целей. Дарвин понимал, что возникновение столь явно совершенных адаптации можно объяснить с точки зрения естественного отбора. В настоящее время большинство биологов согласны с тем, что теория естественного отбора адекватно объясняет даже самые сложные адаптации, хотя некоторые исследователи все еще спорят с этим. Эта глава посвящена ряду проблем, которые возникают, если рассматривать поведение как результат эволюционного развития. Сначала мы обсудим закономерности оптимальной организации пищедобывательного поведения животного, т.е. тему, в которой весьма отчетливо выступают многие из указанных проблем, а затем рассмотрим вопрос о том, каким образом можно справиться с этими проблемами
Во времена Дарвина, чтобы опровергнуть теорию естественного отбора, часто прибегали к телеологическим аргументам. Ученые говорили, что с помощью такого пассивного и случайного механизма, каким представляется естественный отбор, нельзя объяснить очевидной целесообразности, присущей функциональной организации животных и растений. В своих аргументах телеологи использовали понятия цели или намерения для объяснения тех явлений, которые, казалось, невозможно было объяснить по-другому. В настоящее время телеологическую философию уже не считают пригодной при обсуждении проблем эволюции или закономерностей функционирования живого. Однако эта философия все еще играет значительную роль, когда речь идет о цели при обсуждении целенаправленного поведения отдельных организмов.
В последние годы в биологическом мышлении важную роль стало играть представление о запланированности, поскольку мы теперь понимаем, что естественный отбор - это формирующий агент. Современная эволюционная теория подразумевает, что животные стремятся приобрести такие свойства, которые гарантируют им, что в определенных стабильных условиях они не окажутся в худшем положении по сравнению с другими животными, с которыми они конкурируют. Иными словами, обитая достаточно долго в условиях стабильной среды, животные стремятся приобрести те свойства, которые в существующих условиях будут оптимальными. Это совсем не означает, что каждое животное прекрасно адаптировано ко всем условиям, поскольку все индивидуумы различаются между собой генетически и поскольку многих из них конкурирующие особи вытесняют из наиболее благоприятных местообитаний, к которым они были лучше всего приспособлены.
Многие животные способны выбирать из различных видов пищи ту, которая соответствует их физиологическим потребностям. Но для того чтобы получить пищу, животные должны расходовать энергию. Кроме того, им приходится тратить ценное время, расходовать такие физиологические "товары", как тепло и вода, а также подвергать себя опасности, рискуя быть съеденным хищником. Поэтому представляется очевидным, что способы добывания пищи неравноценны, если их рассматривать с точки зрения общей приспособленности животного.
Рассматривая процесс добывания пищи в этом аспекте, мы сначала должны выделить и охарактеризовать несколько основных эволюционных стратегий. Иногда полезно считать, что животные намеренно используют стратегию, направленную на увеличение будущего представительства своих генов, хотя по сути такая эволюционная стратегия является всего лишь пассивным результатом естественного отбора. Подобно тому, как мы можем говорить о стратегиях оборонительного и репродуктивного поведения, мы можем говорить и о стратегиях добывания пищи. Так, например, для некоторых видов животных характерна стратегия "сидеть и ждать", чтобы напасть на жертву из заранее выбранного укрытия. Одни виды охотятся, подкарауливая добычу, другие - преследуют ее. Приведем более конкретный пример. Весьма распространены случаи, когда для получения пищи один индивидуум извлекает пользу из пищедобывательных усилий другого индивидуума.
Такие взаимоотношения могут наблюдаться как между двумя особями различных видов, так и между особями одного и того же вида. В данном случае такие взаимоотношения в принципе мало чем отличаются от некоторых стратегий полового поведения, когда самцы более низкого ранга спаривались с самками украдкой от доминантов.
Барнард и Сибли исследовали альтернативные стратегии при добывании пищи в группе живущих в неволе домовых воробьев.
Оказалось, что в этом случае поставщики получают большую часть своей пищи за счет активного добывания мучных червей, предлагаемых экспериментатором. Что касается попрошаек, то они получают большую часть пищи, копируя поведение активно питающихся птиц или постоянно следуя за ними, вместо того чтобы самим активно разыскивать пищу. Иногда попрошайка выхватывал пищу у поставщика. Рассуждая теоретически, мы должны были бы ожидать, что "выигрыш" попрошаек должен увеличиваться с ростом числа поставщиков, поскольку увеличивается возможность паразитировать на них. Выигрыш поставщиков, напротив, должен уменьшаться с ростом числа попрошаек, поскольку у поставщиков становится меньше шансов извлечь выгоду из своих усилий, потраченных на добывание пищи.
Ситуация такого рода сама наталкивает на мысль проанализировать ее с позиций эволюционно стабильных стратегий, что и было сделано Барнардом и Сибли. Эти авторы обнаружили, что отдельные птицы не сменяют оппортунистически свою стратегию в соответствии со структурой стаи как целого. Попрошайки питаются лучше в том случае, когда поставщиков больше, однако при значительном превышении числа поставщиков питание попрошаек ухудшается. Вероятно, это происходит потому, что большое число поставщиков очень быстро уничтожает имеющуюся в наличии пищу.
Чтобы провести количественный анализ стратегии добывания пищи, необходимо допустить, что поведение животного предназначено для того, чтобы максимизировать некую величину. В том анализе, который провели Барнард и Сибли, в качестве выигрыша фигурировало число мучных червей, съеденных птицей за 10 мин. Иначе говоря, они предположили, что птицы максимизируют скорость потребления пищи. Совершенно очевидно, что такое допущение не вполне правомерно, поскольку можно было учесть и другие факторы, которые влияют на приспособленность животного в случае выбора той или иной стратегии. В частности, могут различаться "стоимости" этих двух стратегий. Однако, чтобы провести анализ такого типа, часто бывает удобно принять достаточно простой показатель приспособленности, например скорость получения пищи, или некую меру эффективности пищедобывания.
Когда хищник ловит свою жертву, ему приходится платить цену в виде времени, потраченного на то, чтобы поймать и съесть ее. Такое время часто называют временем "взаимодействия".
Эта цена с лихвой окупается за счет чистой энергетической стоимости добычи, т.е. валовой ценности минус энергия, затраченная на поимку жертвы и ее переваривание. Рентабельность
жертвы представляет собой чистую энергетическую стоимость, деленную на время взаимодействия. Когда животное имеет возможность выбирать добычу, то можно предположить, что оно выберет наиболее рентабельную жертву. Например, в одном исследовании ушастому окуню представлялась возможность охотиться в большом аквариуме на водяных блох - дафний.
Исследователи обнаружили, что при низкой плотности жертвы рыбы не проявляют никакого предпочтения по отношению к мелким, средним или крупным дафниям. Однако при изобилии дафний рыбы выбирают из них самых крупных и наиболее рентабельных, игнорируя мелких. Этого результата можно было бы ожидать, если исходить из модели рентабельности, показанной на рис.1.
Правда, это можно объяснить и по-другому, а именно: рыба хватает любую дафнию, которая в данный момент кажется ей самой крупной. Ближайшая к ней дафния может показаться ей по размерам больше, чем более крупная, но находящаяся дальше от нее. По мере возрастания плотности дафний возрастает и вероятность появления рядом крупных дафний. Выбирая каждый раз добычу, которая выглядит крупнее, рыба может вести себя таким образом, как это предсказывает модель рентабельности. Заметим, однако, что эта модель отражает, что именно должно произойти в данной ситуации, а модель "брать самую большую" описывает правило, которым рыба может реально пользоваться.
 |
Рис.1.
Простая модель оптимального пищедобывательного поведения. Каждый тип жертвы характеризуется его рентабельностью E/h. Точками и штрихами вычерчены графики, крутизна которых отражает вероятные значения показателя для каждого типа жертвы. Если хищник поедает только более рентабельные жертвы, его поведение описывается сплошной линией. Ее крутизна меньше, чем крутизна пунктирной линии, поскольку хищнику приходится тратить время еще и на поиск более крупной жертвы. Если крутизна сплошной линии равна крутизне штриховой линии или меньше ее, то это означает, что для хищника имеет смысл поедать более мелкую добычу. Таким образом, по мере того как более крупные жертвы становятся все более редкими, сплошная линия поворачивается по часовой стрелке, и животное переключается на менее рентабельную добычу. |
Госс-Кастард исследовал пищедобывательное поведение травника,
болотной птицы, которая добывает пищу по берегам рек и на болотах. Он обнаружил, что когда эти птицы питаются исключительно червями полихетами,
они, как правило, не обращают внимания на мелких червей, а собирают тех, которые по размеру превышают некоторую величину. Эта величина зависит от того, как часто птицы находят более крупных червей, и не зависит от того, как часто им встречаются мелкие. Полученные Госс-Кастардом данные согласуются с представлениями о том, что стратегия добывания пищи у травника направлена на максимизацию энергетической рентабельности, т.е. травник выбирает червей, которые обеспечивают ему получение наибольшего количества энергии на единицу энергии, затраченной на добывание пищи.
Мелкие черви приносят не столь большую при быль из-за того, что у них низкая отдача чистой энергии в пересчете на время, затраченное на их добывание.

Рис.2.
Показатель риска быть съеденным травником для червей-полихет как функция от плотности Corophium
на данном участке территории. Каждая точка изображает усредненную величину показателя риска, полученную при исследовании определенного участка.
Если проводить анализ с точки зрения "ценности", эти результаты могут привести к заключению, что травник принимает решение о том, какую добычу использовать в пищу, на основе оценки энергетического баланса. Однако ГоссКастард обнаружил еще и следующее: если в распоряжении птицы кроме полихет были еще и бокоплавы Corophium,
то птицы предпочитали Corophium
. Можно было предположить, что в тех условиях, где обычно живет Corophium,
труднее отыскать полихет, но Госс-Кастард мог не принимать в расчет такую возможность, поскольку, как было им обнаружено, некоторые птицы предпочитают только червей, тогда как большая их часть поедает Corophium.
На основе своего предыдущего исследования Госс-Кастард высказал гипотезу о том, что когда травник поедает Corophium,
он достигает более высокой скорости поступления "чистой" энергии, чем тогда, когда питается полихетами. Однако когда он стал анализировать энергетическую ценность добычи и затраты энергии на получение добычи, оказалось, что, питаясь исключительно червями, птица должна была бы получить в 2-3 раза больше энергии в минуту, чем она получала, питаясь Corophium.
Совершенно ясно, что при наличии Corophium
пищедобывательное поведение травника определяет не одна энергия. По всей вероятности, кроме энергии Corophium
содержит еще что-то, что является важным для травника.
Таблица 1.
Сравнительные величины скорости получения травником энергии на трех исследуемых участках, где птицы поедали главным образом Corophium,
и скорости получения энергии, которой они могли бы достичь на тех же участках, поедая вместо этого только червей.
| Скорости поглощения энергии, кал • мин - 1
|
| Участок |
потенциальная |
фактическая |
| 9 |
234 |
88 |
| 10 |
224 |
70 |
| 11 |
185 |
93 |
Таблица 2.
Усилия, затраченные птицами для накопления 1 ккал на трех обследованных участках в случае, когда птицы питались главным образом Corophium,
как это и происходило в действительности, и в гипотетическом случае, когда они питались бы только Nereis.
| Участок |
Расстояние, на протяжении которого травник производил поиск, м |
Число клеваний и сделанных проб |
Время, затраченное на проглатывание добычи |
| Corophium
|
Nereis
|
Corophium
|
Nereis
|
Corophium
|
Nereis
|
| 9 |
103 |
42 |
165 |
62 |
48 |
| 10 |
150 |
44 |
671 |
167 |
121 |
49 |
| 11 |
106 |
56 |
543 |
198 |
79 |
48 |
Многие животные редко имеют возможность сконцентрироваться на одной какой-то деятельности, пренебрегая другими. Это особенно верно в отношении животных, которые могут подвергаться опасности со стороны хищников. Так, например, Барнард исследовал пищедобывательное поведение домовых воробьев на одной из английских ферм зимой. Воробьи кочевали стайками различных размеров и кормились в двух местах, которые отчетливо различались по степени риска подвергнуться нападению хищников: кошек и хищных птиц. Внутри коровника воробьи поедали зёрна ячменя, которые они отыскивали в соломенной подстилке. Их пищедобывательное поведение зависело главным образом от количества зерен и было относительно независимым от числа птиц в стае. Однако когда воробьи находились в открытом поле, они периодически прерывали еду и осматривались, нет ли поблизости хищников. В этом случае скорость потребления пищи в большей степени зависела от величины стайки и в меньшей - от количества зерен. В большой стае каждая птица может потратить больше времени на еду, поскольку ей не нужно быть столь бдительной, как в малой стае. Имеются данные, свидетельствующие о том, что в больших стаях птицы обнаруживают хищников гораздо быстрее, чем в небольших. Кроме того, находящиеся в открытом поле воробьи будут тратить меньше времени на осмотр пространства, если они питаются вблизи кустарников или живой изгороди, которые могут служить им укрытием от хищников. Весной на свежезасеянных ячменем полях они держатся обычно вблизи укрытий, хотя в открытом поле зерна больше.
Результаты этого исследования наводят на мысль о том, что в пищевом поведении воробьев встречается несколько вариантов своего рода компромисса, или баланса, между пищедобывательным поведением и поведением, направленным на защиту от хищников. Когда опасность встречи с хищником велика, птицы кормятся менее интенсивно.
 |
 |
| Рис.3.
Скорость потребления пищи у воробьев в зависимости от величины стаи, когда воробьи питаются в коровнике. |
Рис.4.
Скорость потребления пищи у воробьев в зависимости от величины стаи, когда воробьи питаются в открытом поле. |
Они каким-то образом выбирают для питания относительно безопасные, хотя и менее выгодные места, избегая более выгодных участков, где они подвергались бы большему риску. Барнард обнаружил, что расстояние до приближающегося хищника, при котором птицы взлетали, зависит от количества зерен в том месте, где они кормились в этот момент. Это, вероятно, обусловлено большей выгодностью питания по сравнению с риском нападения хищника, а с точки зрения механизмов этого поведения его можно, по-видимому, объяснить тем, что птицы больше поглощены едой в более богатых зернами местах и не так быстро замечают хищника. Вопрос о том, каким образом животные распределяют свое внимание при решении различных задач, приобретает особую важность, когда мы переходим к рассмотрению механизмов, лежащих в основе процесса принятия решения у животных.
Большинство конструкторских решений требует компромисса между теми преимуществами, которые можно извлечь из несовместимых друг с другом особенностей данной ситуации. Так, например, если рассмотреть конструкцию простого кровеносного сосуда, то окажется, что затраты энергии на трение при протекании крови через сосуд уменьшаются с увеличением радиуса сосуда, тогда как затраты энергии на поддержание тонуса сосудистой стенки возрастают. Такой тип прямого сравнения затрат и выгод называется компромиссом. Удачной обычно бывает такая конструкция, в которой используются балансовые зависимости, присущие данной ситуации. Отсюда следует, что выявление балансовых зависимостей в поведении животных может служить хорошей отправной точкой при анализе того, таким образом действует естественный отбор.
Зах выявил простой пример баланса в пищедобывательном поведении ворон, которые питаются брюхоногими моллюсками на западном побережье Канады. Во время отлива они ловят моллюсков, выбирая обычно самых крупных из них. Отыскав такого моллюска, ворона поднимается с ним в воздух над скалистым берегом, а затем бросает раковину вниз; раковина разбивается, и ее съедобное содержимое становится доступно птице. Бросая моллюсков различных размеров с разной высоты, Зах обнаружил, что число бросаний, необходимое для того, чтобы раковина разбилась, зависит от высоты, с которой бросают раковину.
 |
 |
| Рис.5.
Энергетические затраты на поддержание тока крови в зависимости от радиуса кровеносного сосуда. F -
расход энергии на трение; М
расход энергии на поддержание тонуса стенки сосуда; Т -
общие энергозатраты; р* -
оптимальный радиус сосуда, при котором величина Т минимальна |
Рис.6.
Число бросаний раковины, необходимых для того, чтобы она разбилась, в зависимости от высоты падения раковины. |
Воронам приходится расходовать энергию для того, чтобы взлететь на какую-то высоту и бросить моллюска. Поэтому Зах рассчитал общее количество взлетов, которое должна была сделать ворона, чтобы разбить раковину, бросая ее с данной высоты. Оказалось, что расход энергии на взлеты будет наименьшим, если раковину бросать с высоты около 5 м. Это хорошо видно на рис.7. Как и в конструкции кровеносного сосуда, здесь мы встречаемся с балансом - на этот раз между затратами в виде числа бросаний раковины, требующихся для того, чтобы ее разбить, и высотой бросания. Расчеты, проведенные на основе этого баланса, показали, что существует некоторая оптимальная высота бросания, аналогичная оптимальному радиусу кровеносного сосуда. Зах обнаружил, что высота, с которой вороны обычно бросают раковины, близко соответствует расчетной оптимальной высоте бросания, равной 5 м. Таким образом, создается впечатление, что в поведении ворон заложена какая-то программа, которая использует характерные особенности той ситуации, в которой им приходится добывать пищу.
Расчеты затрат и выгод, которые относятся к питающимся моллюсками воронам, так же как и расчеты конструкции кровеносных сосудов, очень упрощает то обстоятельство, что все балансируемые факторы можно измерить с точки зрения затрат энергии. Расчеты Заха дают основание заключить, что вороны выбирают раковины определенного размера и бросают их с определенной высоты, получая при этом наибольший выигрыш в чистой энергии. Однако не всегда возможно или даже желательно рассматривать оптимальный баланс только с энергетической точки зрения. Как мы уже видели в случае травника, энергия не всегда играет определяющую роль. Более того, животные далеко не всегда имеют возможность сконцентрировать всю свою активность исключительно на добывании пищи. Им приходится еще и остерегаться хищников, и охранять территорию от соперников и т.д. Поэтому необходимо рассматривать ситуацию в целом, не забывая, что добывание пищи - это всего лишь один из видов активности в поведенческом репертуаре животного и что затраты или приобретение энергии не единственный результат поведения.
У большой синицы песня играет важную роль в защите территории; кроме того, она используется, чтобы заявить о себе. Ранней весной птица поет в основном на рассвете, начиная свою песню в тот момент, когда солнце должно вот-вот показаться на горизонте. Такое хоровое пение ранним утром типично для многих птиц. Однако в этом есть что-то непонятное: ведь маленькие птички в большинстве своем теряют в течение морозной ночи до 10% массы тела, и поэтому, проснувшись, они должны испытывать сильный голод. Почему же тогда по утрам они прежде всего начинают петь, а не добывать пищу? По-видимому, для этого существуют две основные причины. Во-первых, защита территории особенно важна сразу после рассвета, поскольку именно в это время не имеющие территории птицы проверяют прочность "линии обороны", чтобы обнаружить какой-нибудь участок, освободившийся накануне в результате деятельности хищников.
 |
| Рис.7.
Суммарная высота взлета вороны минимальна при такой высоте, с которой обычно вороны бросают раковины. |
Во-вторых, выгоды от добывания пищи в это время невелики, потому что при слабой освещенности эта деятельность оказывается малоэффективной. Каселник в лабораторных экспериментах показал, что большие синицы не могут эффективно добывать пищу при освещенности ниже определенного уровня, а этот уровень достигается обычно через 40-80 мин после рассвета в зависимости от погоды. Таким образом, птицы, по всей вероятности, извлекают гораздо больше пользы от того, что сначала поют, а потом начинают кормиться, чем если бы они поступали наоборот.
Чтобы судить о том, является ли одна последовательность действий лучше, чем другая, нам нужно иметь какую-то общую "валюту", с помощью которой можно было бы сравнить эти последовательности. Нам также необходимо иметь определенный критерий, с помощью которого можно было бы измерять относительную ценность различных видов поведения. Каждый вид активности характеризуется затратами и выигрышами, связанными с ее реализацией. В этой главе мы рассмотрим вопрос о том, как можно точно оценить эти затраты и выигрыши, однако пока мы ограничиваемся общими рассуждениями. В конечном счете, какая разница, приводит ли данная конкретная активность к увеличению или снижению общей приспособленности животного?
 |
| Рис.8.
Зависимость освещенности мест, где кормится большая синица, от времени суток. Р
и S
обозначают соответственно уровни освещенности, при которых рентабельность пищевого поведения и эффективность поиска пищи достигают 95% своей максимальной величины. |
Большая синица, которая поет на рассвете, повышает свою приспособленность тем, что не допускает на участок конкурентов, но в то же время ее приспособленность уменьшается за счет того, что она откладывает на потом добывание пищи, особенно если она потеряла в весе за ночь. Такое повышение приспособленности удобно называть выигрышем,
а понижение - затратами.
Так, например, при добывании пищи животное выигрывает, приобретая энергию, но может потерпеть убыток, пренебрегая защитой своего участка или становясь более уязвимым для хищника. Одно из преимуществ оценки поведения животного с точки зрения затрат и выигрыша состоит в том, что оно позволяет нам сравнивать относительные ценности совершенно разных активностей. Различные виды пищедобывательного поведения можно сравнить с точки зрения приобретенных неэнергетических компонентов пищи и энергии, а различные типы защиты территории можно сравнивать по их эффективности в противостоянии пришельцам. Однако при сравнении пищедобывания и поведения защиты мы должны определять, намного ли каждая из этих активностей увеличивает или уменьшает общую приспособленность животного. Приспособленность - это единственная общая "валюта", с помощью которой мы можем проводить сравнения различных видов поведения. В конечном счете именно с помощью этой валюты природа производит такое сравнение, поскольку приспособленность определяется через действие естественного отбора.
Представляется важным не только иметь возможность определить, к каким последствиям могут приводить различные виды активности в целом, но еще и оценить их с точки зрения некоторого критерия. Так, например, вполне возможно, что при рассмотрении кандидатур на должность преподавателя университета мы захотим сравнить кандидатов по обычным показателям, таким, как педагогические способности и его потенциальные возможности как исследователя. Кроме того, нам нужен еще и критерий, с помощью которого мы могли бы решить, кому из кандидатов отдать предпочтение: хорошему преподавателю, но плохому исследователю или слабому преподавателю, но хорошему исследователю. Подобным же образом при оценке поведения животного нам необходим критерий оптимальности, который позволял бы судить о том, будет ли при определенных условиях один вид активности лучше, чем другой. Хотелось бы надеяться, что в конечном счете мы сможем определить критерий оптимальности с точки зрения общей приспособленности животного. Однако для того чтобы сделать это грамотно, потребовались бы обширные знания о состоянии животного и об условиях, в которых оно находится. В следующем разделе мы рассмотрим возможные подходы к получению таких знаний.
Мы уже видели, что животным часто приходится выбирать между альтернативными последовательностями действий, характеризующихся определенным балансом затрат и выгод. Наилучшим вариантом поведения животного будет тот, который обеспечивает наименьшие затраты или наибольшие выгоды. Подходя с эволюционных позиций, мы можем ожидать, что хорошо организованные животные должны вести себя так, чтобы максимизировать свою общую приспособленность. Таким образом, концепция затрат и выгод оказывается связанной с концепцией приспособленности.
Животные располагают большим разнообразием способов осуществления затрат и получения выгод, которые соответствуют давлениям естественного отбора. Естественный отбор действует на животное в целом; однако анализируя затраты и выигрыши от той или иной формы поведения, мы должны разделять затраты и выигрыши, которые могут быть приписаны каждому виду внутреннего состояния и поведения животного. Совокупность характеристик животного, представленная в таком виде, называется функцией затрат. Ее можно определить как характеристику мгновенного уровня риска, которому подвергается животное, когда оно находится в определенном внутреннем состоянии и осуществляет определенную деятельность в определенных условиях среды.
Предположим, например, что мы изучаем поведение серебристой чайки,
насиживающей яйца в гнезде. Обычно оба родителя насиживают яйца по очереди, и если только гнездо останется без внимания, его могут быстро разграбить соседи. Сидящая птица не оставляет гнезда до тех пор, пока ее не сменит супруг или не спугнет какой-либо хищник, например лисица или человек. Обычно птица покидает гнездо для добывания пищи и возвращается к нему через несколько часов. Однако иногда ее возвращение может задержаться из-за какой-либо неудачи, несчастного случая, а порой и из-за того, что ее может поймать экспериментатор. Что в таком случае должна делать сидящая на гнезде птица? С одной стороны, ее супруг может вернуться в любое время, но с другой - по мере течения времени у нее усиливается чувство голода. В конце концов эта птица будет вынуждена покинуть гнездо, чтобы добыть пищу. Поскольку серебристые чайки способны размножаться круглый год, с генетической точки зрения представляется невыгодным подвергать опасности жизнь особи из-за единичной кладки яиц. Вопрос в том, в какой именно момент птица должна решиться покинуть гнездо?
Если "наседка" улетает с гнезда слишком рано, то она подвергает опасности свою кладку без особой на то необходимости, потому что ее супруг, быть может, просто ненадолго задерживается. Если же птица покинет гнездо слишком поздно, то она необоснованно подвергает опасности свою жизнь, поскольку она может не найти пищу так быстро, как это бывает обычно. Для того чтобы точно определить, когда же птица должна улететь с
кладки, необходимо количественно оценить степени риска в каждой из этих альтернативных ситуаций. Такую оценку эффективно осуществляет природа, создавая животное, которое будет принимать самое подходящее в этой ситуации решение.
Чтобы изучить эту проблему, необходимо в полевых условиях измерить степень риска, которому подвергаются животные, занятые различными видами активности. Так, например, в случае насиживающих яйца серебристых чаек нам необходимо знать степень риска, которому подвергнутся яйца, если останутся в гнезде без присмотра, и степень риска, которому подвергнется птица, если будет продолжать сидеть на гнезде без пищи. Были предприняты попытки в самых различных экспериментах оценить затраты и выгоды, присущие поведению насиживания. Так, например, Дрент изучал поведение насиживания яиц у серебристых чаек на о. Схирмонниког.
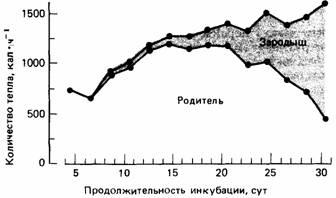
Рис.9.
Энергетическая цена инкубации яиц у серебристых чаек, вычисленная для всей кладки. Тепло, которое производит сам зародыш, изображено в виде затушеванной области. Таким образом, верхняя кривая отражает общее количество необходимой энергии, а нижняя - энергозатраты родителей.
Он обнаружил, что 20% яиц уничтожается хищниками, а 12% погибает от того, что не соблюдаются физические условия, необходимые для их инкубации. Если человек не тревожит птиц, они проводят на яйцах около 98% времени. Благодаря этому яйца защищены от переменчивых воздействий внешней среды, а птица имеет возможность регулировать температуру яиц. Главная проблема, с которой сталкивается птица, насиживающая яйца, - это регулирование температуры собственного тела. Холодной ночью ей приходится продуцировать тепла столько, чтобы его хватило для согревания себя и яиц, тогда как в полуденную жару ей приходится дышать очень часто, чтобы несколько охладить себя. Дрент присоединял датчики к гнезду и яйцам и с помощью следящей аппаратуры смог детально исследовать роль факторов, влияющих на развитие зародышей чаек. В самом начале инкубации температура яиц полностью зависит от родителей, но по мере развития зародыш начинает продуцировать метаболическое тепло. Однако зародыши не способны к терморегуляции, и поэтому, если оставить яйца открытыми, они будут постоянно охлаждаться. У зародыша нет компенсаторной теплопродукции, а вот только что вылупившийся птенец реагирует на снижение температуры воздуха повышением теплопродукции. Таким образом, птенец в какой-то степени обладает температурным гомеостазом.
В отсутствие родителей зародыш рискует погибнуть либо от перегревания, либо от переохлаждения в зависимости от того, какая стоит погода. Однако более важным фактором является общее количество тепла, которое получает зародыш к моменту его вылупления. Скорость развития зародыша зависит от интенсивности его метаболизма, на которую в свою очередь влияет температура внутри яйца. Если эта температура низкая, то зародыш развивается медленно. Во время инкубации яйцо постоянно теряет в массе за счет испарения воды.

Рис.10.
Потери воды яйцами серебристой чайки, измеренные по изменениям их массы.
По мере потери воды воздушное пространство внутри яйца увеличивается, однако слишком большая потеря воды приводит к обезвоживанию зародыша.
 |
Рис.11.
Сохранность яиц серебристой чайки, оставшихся на виду у хищников в отсутствие владельцев гнезда. |
Такая потеря воды приводит к тому, что в одном конце яйца образуется воздушное пространство, необходимое для дыхания зародыша в период, непосредственно предшествующий вылуплению. Зародыш не должен развиваться слишком быстро, иначе это воздушное пространство не сформируется как следует к тому моменту, когда птенец будет готов к вылуплению. Зародыш не должен развиваться и слишком медленно, иначе яйцо потеряет так много воды, что наступит дегидратация зародыша. Вылупление должно произойти в тот момент, когда яйцо потеряет строго определенное количество воды, а зародыш к этому времени будет достаточно развит, чтобы покинуть яйцо. Не имеет столь большого значения кратковременное охлаждение яиц, важно, чтобы за это время их температура не упала слишком низко. В теплый день яйца подстерегает другая опасность: они могут перегреться, если останутся долго открытыми, но обычно яйца, оставленные открытыми на час или два, с физиологической точки зрения практически не страдают. Таким образом, серебристые чайки во время насиживания яиц вполне могут пережить случайные тревоги, которые вынуждают их покидать гнезда. Однако если чайки будут потревожены не один раз, то средняя скорость притока тепла к яйцам может резко упасть ниже того уровня, который требуется для успешного вылупления.
У зародышей серебристой чайки в два раза больше шансов погибнуть от хищников, чем от нарушения их развития. Сидящим на кладке птицам приходится покидать свои гнезда при появлении некоторых хищников, а также человека. В отсутствие родителей яйца могут стать добычей соседних чаек, крыс, ежей, ворон или других хищников. Уровень риска, которому подвергаются яйца, может быть рассчитан на основе полевых наблюдений за судьбой оставшихся без присмотра яиц или в специальных экспериментах, когда родителей удаляют из гнезда.
Чайка, сидящая в гнезде и ожидающая возвращения своего партнера, не может покинуть гнездо, не подвергая опасности яйца. Как мы уже видели, риск, которому подвергаются яйца, можно количественно оценить как часть общей функции затрат. С одной стороны, имеются затраты как результат прекращения насиживания, поскольку возникает риск, что из яиц не вылупятся птенцы, потому что их съедят хищники или они погибнут от непогоды.

Рис.12.
Эти весы для взвешивания гнезда можно расположить в ямке, выкопанной под гнездом серебристой чайки. Затем гнездо укладывают на платформу весов.
Поведение птиц на гнезде при этом не изменяется, а изменения массы гнезда можно регистрировать с помощью соответствующего электронного устройства.
С другой стороны, имеются затраты, связанные с насиживанием, которые складываются из риска быть пойманным хищником и из риска погибнуть от истощения.
Указанные аспекты функции затрат также могут быть исследованы экспериментально. Например, массу тела насиживающей птицы можно измерить с помощью специальных весов, помещенных в яму, выкопанную в земле под гнездом. Количество жира у птицы можно вычислить исходя из соотношения ее массы и размеров костей скелета, которые измеряются у пойманной птицы перед началом эксперимента в процессе ее мечения. Количество пищи, которое серебристая чайка получила за время кормежки, можно оценить по изменению ее массы, измеренной на тех же весах, помещенных под гнездом, перед и после ее путешествия за пищей. Качество пищи можно оценить путем анализа фекалий, собранных вокруг гнезда, и по результатам наблюдений за птицей во время добывания ею пищи. Серебристые чайки могут летать за пищей на расстояния в несколько миль. Поэтому при проведении наблюдений за их пищедобывательной активностью некоторых птиц снабжали радиопередатчиками и прослеживали их маршрут с помощью радиоприемника с направленной антенной. Таким образом, используя различную аппаратуру, иногда удается получить достаточно точные оценки затрат и выигрышей животных, ведущих обычный образ жизни в естественных условиях обитания.
Вообще говоря, функция затрат состоит из двух главных частей. Первую часть составляют затраты, связанные с активностью, осуществляемой в данное время, а вторую - затраты, связанные с тем, что другие активности в это время не осуществляются. Например, чайка, сидящая на яйцах, несет затраты, связанные с их насиживанием. В них входят физиологические затраты, связанные с поддержанием определенной температуры яиц, и поведенческие затраты, связанные с риском быть пойманным хищником во время насиживания. Кроме того, некоторые затраты обусловлены тем, что насиживание несовместимо с другими видами поведения, обычно связанными с иными мотивациями животного. Так, например, во время насиживания у птиц неизбежно усиливается чувство голода. А голод влечет за собой риск гибели от истощения, поскольку в тот момент, когда животное решит приступить к добыванию пищи, оно может не найти ее. Как мы увидим далее, такой анализ поведения обязательно требует знания особенностей экологии данного животного. Стараясь понять, каким образом животные учитывают затраты и выигрыши, обусловленные особенностями среды их обитания, иногда бывает полезно провести аналогию между экономическими расчетами человека и подобным поведением у животных.

Рис.13.
Серебристая чайка с прикрепленным к
ней радиопередатчиком.
В экономике существуют два основных принципа принятия решения. Во-первых, решение должно быть рациональным, а во-вторых, оно должно исходить из какой-то оценки всех "за" и "против". Принятие решения будет в принципе рациональным, если оно внутренне непротиворечиво, т.е. обладает свойством, обычно называемым транзитивностью выбора.
Предположим, что кто-то должен выбирать между вариантами А, В и С. Если А предпочтительнее В, а В предпочтительнее С, то кажется разумным ожидать, что А будет предпочтительнее С. Тогда мы можем записать логический порядок предпочтения А-В-С.

Рис.14.
Радиоприемное устройство, смонтированное на крыше лендровера, которое используется для прослеживания серебристых чаек на острове Уолни в Камбрии.
В этом случае говорят, что соотношение между А, В и С является транзитивным.
Если же А предпочтительнее В и В предпочтительнее С, однако А не предпочтительнее С, то тогда решение выбрать С, а не А считается иррациональным, а соотношение между А, В и С называется нетранзитивным.
Экономисты основывают свою теорию на концепции "личности с рациональным экономическим мышлением", которая подразумевает, что взаимосвязи между всеми предпочтениями этой личности являются транзитивными.
Рациональность не обязательно подразумевает участие рассудочной деятельности. Человек может принять какое-то решение на основе рассуждения, но может принять рациональное решение и чисто автоматически, как будто он создан или запрограммирован таким образом, чтобы поступить именно так. Люди принимают и нерациональные решения. Фактически невозможно доказать, что человек совершает транзитивный выбор, поскольку для этого понадобились бы повторные эксперименты с осуществлением выбора в идентичных условиях. Но это нереально, поскольку условия никогда в точности не повторяются, хотя бы уже потому, что воспоминание об уже однажды сделанном выборе накладывает отпечаток на условия, в которых предстоит сделать следующий выбор. Экономисты вынуждены принимать условия транзитивности выбора как рабочую гипотезу, как предположение, на котором основана элементарная теория принятия решения.
Транзитивность выбора подразумевает, что в процессе принятия решения некоторая величина должна максимизироваться. Чтобы в этом убедиться, рассмотрим следующую ситуацию. Предположим, что варианты А, В и С можно каким-то образом оценить количественно. Если А оценивается больше, чем В, то можно записать, что А > В. Человек выбирает А, а не В, руководствуясь некоторым принципом максимизации. Если мы знаем, что В больше С, то мы можем написать В > С. Если человек выбирает С, а не А, то это выглядит так, как будто С оценивается выше, чем А. Однако нам известно, что А > В > С, а отсюда следует, что С оценивается ниже, чем А. Таким образом, если человек выбирает С, а не А, то это означает, что он оказывает предпочтение варианту С, несмотря на то что С оценивается ниже, чем А. Человек, который совершает выбор на такой нетранзитивной основе, не может выбирать вариант, имеющий наивысшую оценку; о таком человеке нельзя сказать, что он использует принцип максимизации. И наоборот, если предпочтение человека является транзитивным, мы можем сделать вывод, что он или она пользуются принципом максимизации.
Величина, которую в ситуации выбора максимизирует личность с рациональным экономическим мышлением, называется полезностью. Я
могу получить некоторое количество полезности, если куплю что-то для своей коллекции фарфора, некоторое количество полезности от занятий спортом и еще какое-то от чтения книг. Затрачивая свое время и деньги на эти вещи, я выбираю то, что максимизирует количество получаемого мною удовлетворения или полезность. Я не отдаю себе отчета в том, что я максимизирую полезность; однако если я поступаю рационально, мое поведение выглядит именно таким образом. Следовательно, полезность - это воображаемая мера
психологической ценности различных благ, досуга и т.д. Воображаемая - потому что на самом деле мы не знаем, влияет ли она на поведение выбора, который осуществляют люди. Мы только предполагаем, что они ведут себя таким образом, как будто максимизируют полезность.
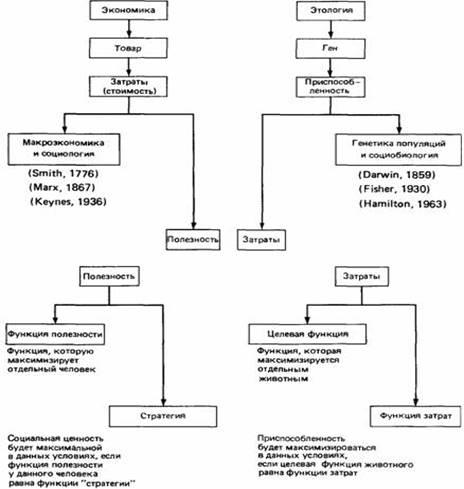
Рис.15.
Параллель между понятиями экономики и этологии. Вверху.
Общая схема, на которой указаны фамилии исследователей, внесших наибольший вклад в развитие этих областей науки, и даты публикации их основных работ. Внизу.
Детализированная схема в приложении к отдельному человеку или животному.
Что касается поведения животных, то здесь эквивалентом полезности выступает выигрыш - величина, противоположная затратам. Подобно тому как полезность представляет собой воображаемую меру ценности поведения, затраты являются воображаемой мерой изменения приспособленности, связанного с поведением животного и его внутренним состоянием.
Мы не знаем, включается ли учет затрат непосредственно в процесс принятия решения у животных. Мы только можем проверить гипотезу о том, что животные ведут себя таким образом, как будто они максимизируют выигрыш. Компоненты затрат включают в себя факторы, связанные с поведением, осуществляющимся в какое-то конкретное время, и с риском, связанным с внутренним состоянием животного. Так, например, чайки, насиживающие яйца, несут физиологические затраты за счет того, что обеспечивают теплом яйца, и они же несут затраты в связи с тем, что у них возрастает чувство голода, когда они сидят на гнезде.
 |
 |
| Рис. 19.
Те же линии уровня функции полезностей, взятые из рис.18 и наложенные на рис.16. Штриховой линией представлено наилучшее возможное сочетание червей и ракообразных. |
Рис. 20.
Различные бюджетные линии соответствуют различным количествам энергии, которыми изначально располагает животное. Обратите внимание на то, что оптимальный вариант поведения остается одним и тем же. |
 |
Рис.21.
Изменение положения бюджетной линии при изменениях цены товара X.
Обратите внимание на изменения оптимального предпочтения. |
Различные затраты можно объединить в виде функции затрат, которая оценивает каждый аспект состояния и поведения животного с точки зрения связанных с ними затрат. В экономике эквивалентная функция называется функцией полезности
. Здесь, может быть, имеет смысл проанализировать конкретный пример поведения животного затрат на определенные виды активности или от цены на товары. В тех примерах, которые мы разбирали до сих пор, энергетические затраты служили общей мерой при оценке возможных альтернативных видов активности. Однако такое наблюдается не всегда. Поэтому было бы ошибкой рассматривать энергию как общую валюту, используемую в процессе принятия решения, хотя в некоторых случаях энергия и может служить своего рода общим "знаменателем". Энергия будет служить общей мерой для всех альтернатив только в том случае, если наличный запас энергии в разбираемых обстоятельствах будет действовать как ограничивающий фактор. Пример из экономики, вероятно, поможет нам лучше понять эти различия.
Предположим, что человек зашел в универсам, чтобы приобрести различные товары. Как мы уже видели, каждый предмет, который может быть куплен, характеризуется некоторой функцией полезности, а полезность определенных товаров будет зависеть от предпочтения и опыта покупателя. Такие факторы, как аппетитность, товарный вид и состав продукта, будут влиять на полезность пищевых товаров. Понятие полезности дает возможность сравнивать самые различные товары. Так, например, человек может получить одну и ту же полезность и от пакета с растительной смесью, и от бутылки содовой воды. Полезность, таким образом, представляет собой валюту, с помощью которой можно оценить различные предметы. Однако другой валютой, общей для всех предметов, являются деньги. Эта валюта приобретает особое значение, если покупатель может истратить лишь определенную сумму, что ограничивает количество его покупок. Однако, хотя каждый продукт в универсаме и имеет свою цену, деньги не всегда выступают в роли реального ограничителя. Если покупатель очень торопится, он может потратить денег больше, чем в том случае, когда он располагает временем. Время, таким образом, становится ограничителем, который "берет за горло", и может случиться так, что покупатель гораздо быстрее произведет выбор среди бутылок с содовой водой, чем среди различных пакетов с растительной смесью. Время, необходимое для выбора каждого предмета, будет определять наклон линии временного бюджета, и может оказаться, что выбор, сделанный покупателем в условиях дефицита времени, будет отличаться от того выбора, который бы он сделал при ограниченных деньгах.
Из приведенного выше примера мы должны понять, что существует много возможных ограничений, которые оказывают влияние на поведение выбора у животных. В зависимости от обстоятельств эти ограничители могут меняться, но только тот ограничитель, который в данный момент "берет за горло", имеет существенное значение. Именно поэтому время и энергию нельзя путать с полезностью или затратами. Проводя аналогию между животным и моделью потребителя в экономике, можно считать, что затраты эквивалентны полезности, энергия - деньгам, но время эквивалентно времени. Животное может заработать энергию за счет добывания пищи и может потратить ее на различные другие активности. В добавление к основному постоянному уровню метаболических расходов это животное может сберечь энергию путем запасания пищи или депонирования жира, а может и потратить ее на различные виды активности, включая и добывание пищи. Если цена активности высока, энергетический бюджет животного испытывает сильные ограничения. Когда же эта цена уменьшается, животное ощущает, что его реальный энергетический доход увеличивается, а бюджетные ограничения ослабевают.
Если на приобретение различных товаров и на осуществление какой-то деятельности мы располагаем ограниченной суммой денег, то мы часто распределяем эту сумму в соответствии с нашими нуждами. Обычно мы учитываем все наши расходы на какой-то определенный период времени - день, неделю или год - и называем это бюджетом. Можно составить бюджет предстоящих расходов, а можно - тех, которые уже были произведены. В одном случае мы ассигнуем определенные суммы денег на определенные цели, а в другом - мы подводим итог расходам, которые уже произвели. В любом случае ведение бюджета предполагает определенную дисциплину в расходовании денег.
Мы уже видели, что деньги в потребительской экономике - это аналог энергии в поведении животных, поэтому естественно спросить, а есть ли у животных энергетический бюджет. Однако мы видели также, что энергия - это только один тип ограничителя, который воздействует на поведение животного. Другим важным ограничителем является время, и мы также можем задаться вопросом, приложимо ли понятие "бюджет времени" к поведению животных. Бюджет времени или энергии - это не просто подсчет того, как животное тратит время и энергию. Если бы животное растрачивало время и энергию совершенно беспорядочно, то это означало бы, что оно не имеет никакого бюджета. Однако мы можем ожидать, что естественный отбор сформировал животных такими, что они должны тратить имеющиеся у них энергию и время с максимальной пользой. Поэтому разумно ожидать, что животные используют время и энергию как ценные ресурсы и, следовательно, имеют соответствующие бюджеты.
Хейнрих сравнивал пищедобывательную деятельность шмеля с поведением покупателя:
"Задача, с которой сталкивается шмель, начиная поиск пищи на лугу, где много различных растений, нисколько не отличается от задачи, с которой сталкивается неопытный покупатель, толкающий тележку вдоль торгового зала универсама. Прямо или косвенно они оба стараются получить за свои деньги как можно больше ценностей. Никто из них наперед не знает, что именно содержится в упаковках, которые стоят на полках или находятся на лугу. Но они научаются этому на опыте".
Цветки обеспечивают шмелей энергией, необходимой для выращивания потомства, при этом шмели должны тратить значительное количество энергии на добывание пищи. Шмели способны жить в холодных климатических условиях благодаря их удивительной способности к терморегуляции. Они могут поддерживать высокую температуру тела при низкой температуре окружающего воздуха, что дает им возможность оставаться активными, даже если для этого потребуются высокие энергетические затраты. Они могут сберегать энергию, значительно уменьшая уровень активности и таким образом сохраняя тепло. Когда пищевые ресурсы цветков оказываются бедными, шмелям тем не менее удается извлекать выгоду за счет медленной пищедобывательной деятельности. Когда пищи очень много, шмели повышают температуру тела и интенсифицируют пищедобывательную активность. Следовательно, они рассчитывают свои энергозатраты в соответствии с имеющимися условиями.
Собирая пищу, шмель проводит большую часть времени в полете. Перелетая с цветка на цветок, шмели стараются свести до минимума продолжительность и дальность полета. За час они пролетают от 11 до 20 км и проводят внутри гнезда всего 2-4 мин в паузах между полетами за пищей. Простые расчеты показывают, что для добывающего пищу шмеля бюджет времени более важен, чем бюджет энергии. Предположим, например, что у одного шмеля цветки находятся близко от гнезда и он может постоянно "пастись" на них, тогда как другой шмель летает за взятком за три километра от гнезда. Если второй шмель пролетает по 15 км в час, то он должен тратить на каждый полет 24 мин. Питаясь на кипрее, оба шмеля смогут собрать полный зобик нектара примерно за 10 мин. Шмель, который совершает дальние путешествия, добывает 30 мг сахара за 34 мин, но при этом около 3 мг сахара расходуется во время каждого полета на метаболические процессы. Таким образом, полеты туда и обратно отнимают у этого шмеля примерно две трети времени и только одну десятую часть энергии. За это же самое время шмель, питающийся вблизи гнезда, получил бы 102 мг сахара. Таким образом, имеет смысл тратить время на дальние перелеты лишь в том случае, если растения, растущие далеко от гнезда, в 3,4 раза ценнее, чем растения, растущие поблизости.
Пищедобывательное поведение шмелей изменяется в зависимости от количества нектара. Чем больше нектара в каждом цветке, тем больше они обследуют других цветков в ближайших окрестностях. Хейнрих провел эксперименты, в которых закрывал тонкой прозрачной тканью некоторые участки клеверного поля, а другие участки оставлял открытыми. Шмели выбирали нектар из закрытого клевера, а в прикрытых цветках нектар накапливался. Когда ткань с цветков снимали и шмели могли посещать и богатые, и бедные нектаром участки клеверного поля, оказалось, что на богатом нектаром участке клеверника шмели обследовали около 12 цветков в головке клевера и совершали короткие перелеты между головками. На участке клеверника с низким содержанием нектара шмели проверяли примерно два цветочка в каждой головке и совершали более длинные перелеты между головками клевера. Именно таким путем шмели концентрировали свою пищедобывательную активность на самых выгодных участках клеверника.
Энергетические затраты на добывание пищи в некоторой степени зависят от температуры окружающей среды. Когда шмели "пасутся" на кипрее, они задерживаются на каждом цветке всего лишь 1-2 с, собирая язычком крошечные капли нектара. Однако посещение других цветков длится до нескольких минут. Во время пребывания на цветке шмели не дают охладиться своему летательному аппарату и сохраняют его "полетную" температуру, что обеспечивает им готовность взлететь без всякой задержки. Во время длительного пребывания на цветке они поддерживают температуру грудной части тела около 32° С за счет дрожания грудных мышц; однако шмели не тратят энергии на согревание брюшка, которое не входит в состав двигательного аппарата, обеспечивающего полет.
 |
| Рис.22.
Изменения пищедобывательного поведения шмеля при сборе нектара на двух площадках, засеянных белым клевером. На одной площадке,
где побывало много шмелей, на одну головку цветка приходится всего 0,003 мг сахара; другая площадка была закрыта легкой прозрачной тканью, вследствие чего запасы нектара достигли там уровня 0,01 мг сахара на одну головку цветка. Как видно из диаграмм, если запасы нектара малы, шмели совершают длинные перелеты с одной головки клевера на другую. |
 |
| Рис.23.
Соотношение между временем, которое шмели тратят на перелеты, и временем их пребывания на цветках определяется в основном тем, какой вид растений они посещают. Температура воздуха мало влияет на соотношение. Серыми полосками обозначены результаты, полученные при 20°С. а зелеными при 30°С. |
При температуре окружающей среды около 25°С шмелю нет необходимости согревать грудной отдел во время периодов покоя; если же температура падает ниже этой величины, то шмелю приходится "платить все дороже" для того, чтобы эффективно добывать пищу.
В цветках разных видов растений образуется различное количество нектара, поэтому можно рассчитать относительные затраты на добывание пищи на различных растениях при разной температуре воздуха. Шмели собирают пищу на "прибыльных" цветках рододендрона при самых различных температурах воздуха, тогда как на кальмии и черешне при низкой температуре они "пасутся" не столь интенсивно, поскольку эти растения не могут возместить им дополнительную энергию, требующуюся для терморегуляции.
На примере пищедобывательной активности шмелей видно, каким образом время и энергия ограничивают ее эффективность. Когда шмелю приходится путешествовать на некоторое расстояние, чтобы найти медоносные растения, время становится лимитирующим фактором, и для шмеля выгодно потратить энергию, чтобы сберечь время. Когда же шмель питается на относительно бедных нектаром цветках или добывает пищу при низкой температуре, он может потратить больше времени ради того, чтобы сберечь энергию.
 |
| Рис.А.
Температура тела шмелей при различной температуре воздуха в условиях, когда шмели питаются на "рентабельных" цветках. Б. Расчетные затраты на добывание пищи при различной температуре воздуха у рабочего шмеля массой 0.2 г. который поддерживает температуру груди на уровне 30°С и проводит половину времени в полете, а половину - на цветках. |
Взаимоотношения между временем и энергией часто играют важную роль в поведении животного. Ему приходится распределять эти ресурсы для каждого конкретного типа активности и для всех конкретных условий, с которыми оно сталкивается в данном случае. Кроме того, ему приходится принимать в расчет затраты на переход от одного типа активности к другому. Когда животное переключается с одного поведения на другое, это может потребовать определенных затрат.
Например, питающийся на кукурузном поле голубь может захотеть пить, поскольку в данном случае пищу его составляют сухие зерна, а чтобы получить воду, ему необходимо пролететь более километра. Помимо физиологических затрат на перелет от места кормежки до места, где находится вода, птица будет тратить время на этот перелет и при этом не будет получать никакого выигрыша ни от приема пищи, ни от питья. Более того, во время этого перелета голубь может подвергнуться определенному риску при встрече с хищными птицами или вооруженными фермерами. Таким образом, затраты на переключение с одного вида активности на другую представляют собой уменьшение приспособленности, которое происходит в тот период времени, когда животное переходит от одной активности к другой и не получает пользы ни от одной из них. Эти затраты могут включать в себя потери ценного времени, расход энергии или риск от встречи с хищниками. Теоретически можно показать, что затраты на переключение
следует рассчитывать таким образом, как будто они составляют часть затрат на активность, которую собирается осуществить животное.
Следует различать мгновенные затраты,
которые возникают на каждом отрезке соответствующего периода времени, и совокупные затраты
за весь рассматриваемый период в целом. Весьма важным представляется то, что переключения с одного типа поведения на другое имеют смысл в том случае, если совокупные затраты вскоре после этого переключения станут меньше, чем они были бы, если бы животное вовсе не меняло своего поведения. Мною было экспериментально показано, что если затраты на смену активности высоки, то голуби переходят от пищевого поведения к питьевому менее часто, чем они делали бы это в других условиях.
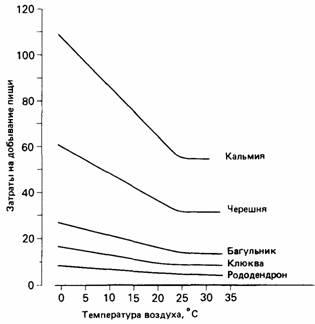 |
Рис.25.
Расчетные затраты на добывание пищи у шмелиной матки при посещении различных видов растений. |
Вместе со Стефаном Ларкином мы получили данные о том, что птицы действительно включают затраты на смену активностей в состав затрат на то поведение, которое они собираются осуществить. Они не сменят своей активности до тех пор, пока не рассчитают этих затрат. Ларкин обнаружил, что временная картина пищевого и питьевого поведения у голубей изменялась вполне предсказуемым образом, если затраты на переключение с одного вида поведения на другой увеличивались, т.е. возрастал расход времени или энергии.
Помимо ежеминутного расчета затрат животные производят и более глобальную оценку бюджета своего времени и своей энергии. В целом можно ожидать, что поведение, основное назначение которого способствовать выживанию особи, будет иметь приоритет перед поведением, которое обеспечивает другие виды приспособленности, таким, например, как территориальное, половое или родительское. Однако виды животных значительно различаются между собой. Некоторые типы поведения существенны для животного всегда, тогда как другие имеют значение только в тех случаях, когда исчерпываются возможности регуляторных физиологических механизмов. Так, для животных некоторых видов питьевое поведение - это каждодневная необходимость, тогда как животные других видов могут обходиться вовсе без него.
Каждый вид активности имеет какое-то значение с точки зрения приспособленности животного, и животным приходится распределять по приоритету различные виды активности как в целом, так и в своей ежеминутной деятельности. Вместе с Хаустоном мы попытались решить эту проблему.
 |
Рис.26.
Затраты на переключение с одной активности на другую. А. Мгновенные затраты на осуществление активности А выше, чем на осуществление активности В, но для переключения с активности А на активность В животное должно осуществить переходное поведение С, мгновенные затраты на реализацию которого еще более высоки. Б. В момент времени Т совокупные затраты животного на последовательность действий А - В - С становятся равными совокупным затратам, которые понесло бы животное, если бы продолжало осуществлять активность А. |
Для этого мы оценивали, каковы будут затраты животного, если оно воздерживается от каждого вида активности, входящего в его естественный поведенческий репертуар. Например, если животное не питается, его затраты будут высокими, однако если оно воздерживается от поведения чистки шерсти, затраты могут быть относительно низкими. Животное, испытывающее пищевую мотивацию и сильную мотивацию чистки шерсти, но имеющее слишком мало времени для удовлетворения обеих этих мотиваций, понесет меньшие потери с точки зрения своей общей приспособленности, потратив имеющееся у него время на питание.
Предположим, что в течение обычного дня своей жизни животное осуществляет различные виды полезного поведения таким образом, как это показано на рис.27. Если условия, в которых живет животное, день ото дня практически не меняются, оно распределяет разные виды своего поведения в соответствии с имеющимся у него временем. А теперь представим себе, что внешняя среда изменилась таким образом, что животному требуется гораздо больше времени, чтобы получить обычное количество пищи. Оно может прореагировать на изменившиеся условия жизни по-разному: может тратить на питание то же самое время, что и раньше, но довольствоваться меньшим количеством пищи; будет стремиться получить обычное количество пищи; выбирать компромиссный вариант между этими двумя крайностями. Если животное будет тратить много времени на получение обычного количества пищи, то у него останется меньше времени на все другие виды активности, которые надо будет "сжать", чтобы уместить в оставшееся время. Мы обнаружили, что степень "сопротивления" любой активности такому сжатию может быть выражена одним параметром, который мы назвали упругостью
этой активности. В том случае, когда животное питается в течение обычного периода времени, но получает меньше пищи, упругость пищевого поведения относительно низка, поскольку оно не "сжимает" другие виды активности, несмотря на усилившийся голод животного. В том случае, когда животное добивается получения обычного количества пищи, упругость пищевого поведения относительно высока, поскольку ему удается потеснить другие виды активности в пределах общего времени, которым располагает животное; при этом само пищевое поведение ни в чем не страдает.
Упругость поведения -
этомера того, до какой степени может быть сжат во времени любой вид активности животного под влиянием других ее видов. Когда время является ограничителем бюджета, у животного наблюдается тенденция игнорировать те виды поведения, которые имеют низкую упругость. Действительно, если при ограничении времени какая-либо активность полностью исчезает из поведенческого репертуара животного, то мы можем назвать ее роскошью или праздной активностью.
Непосредственные измерения упругости поведения почти не проводились, поскольку осуществить их практически очень трудно - для этого нужно в течение нескольких дней хронометрировать поведение животного в ситуации, когда экспериментатор может изменять количество времени, имеющегося в распоряжении животного. Такого рода исследование провел Крофт.
 |
| Рис.27.
Диаграмма, показывающая, каким образом животное может распределять свое время на протяжении суток. |
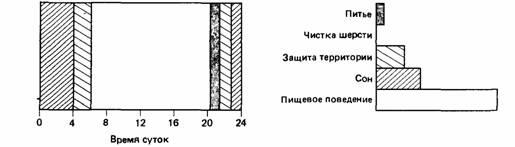 |
| Рис.28.
Диаграмма, показывающая, каким образом то же самое животное может перестроить свой распорядок дня в том случае, когда для получения того же количества пищи придется затрачивать больше времени. |
Он изучил, как влияет длина светового дня на поведение и гормональный баланс самок канареечного вьюрка.
Изменяя длину светового дня, Крофт тем самым изменял время, которым располагали канареечные вьюрки для осуществления разных видов активности, поскольку в темноте они не активны. Крофт обнаружил, что птицы тратят на питание одно и то же время как в короткие, так и в длинные дни, но в длинные дни они питаются более эффективно. В течение длинного дня птицы тратят больше энергии на различные другие виды активности, так что этот результат не столь уж неожидан.
 |
| Рис.29.
Примеры кривых спроса, используемых в экономике. Спрос на кофе является неэластичным, а спрос на рыбу эластичным. |
 |
| Рис.30.
Средняя частота реакций в группах сиамских бойцовых рыбок,
которые проплывали через специальный туннель, чтобы получить пищевое подкрепление или возможность осуществить демонстрацию угрозы по отношению к своему изображению в зеркале.
По оси абсцисс число реакций, требующихся для получения подкрепления. |
Птицы, содержащиеся на режиме длинного дня, тратят больше времени на сон и больше времени не заняты активной деятельностью, чем птицы, для которых установлен короткий день. Поскольку птицы имеют возможность спать ночью, представляется наиболее вероятным, что в условиях длинного дня птицы заполняют сном свободное время. Эту точку зрения подтверждает тот факт, что во время строительства гнезда птица меньше времени отводит на сон в дневное время.
Крофт рассчитал, сколько времени остается у птицы на строительство гнезда за вычетом времени, необходимого для питания и чистки перьев (в расчет принималось и время перелетов с одного места на друroe). Как видно из табл.3, в длинные дни у птиц имеется достаточно времени для постройки гнезда, тогда как в короткие дни его очень мало. Однако интерпретация этих наблюдений осложняется тем обстоятельством, что длинные световые дни стимулируют выработку соответствующих гормонов, и поэтому в этих условиях мотивация строительства у птиц выше, чем в условиях короткого дня. Гнездостроителъное поведение птиц в условиях длинного дня более эффективно, чем в условиях короткого дня, и они тратят меньше времени на излишние повторения собирающих и собственно гнездостроительных движений. Таким образом, хотя понятие упругости поведения с
теоретической точки зрения не связано с понятием мотивации
, при изучении временного бюджета животного трудно отделить влияние упругости поведения от влияния мотивации на это поведение.
Однако упругость можно измерить косвенным путем и посредством функции спроса,
которая используется экономистами для выражения связи между ценой тех или иных благ и их потреблением. Например, когда повышается цена на кофе, люди продолжают покупать его примерно столько же, сколько и раньше, - разве что чуть-чуть меньше. Однако стоит подскочить цене на фрукты, как спрос на них падает. Когда же повышается цена на свежую рыбу, спрос на нее снижается очень сильно. По всей вероятности, люди готовы платить большую цену, лишь бы не нарушить своей привычки пить кофе. Спрос на кофе называют неэластичным
. Однако, когда повышается цена на свежую рыбу, люди стараются покупать ее меньше или переключаются на другие продукты, например мясо или рыбные консервы. Спрос на рыбу называют эластичным.
В поведении животных наблюдается явление, в точности аналогичное описанному. Если животное тратит определенное количество энергии на какую-то деятельность, то, как правило, оно будет меньше заниматься этой деятельностью, если энергетические траты на нее возрастут.
Таблица 3.
Расчетные величины среднего времени, которым располагают для строительства гнезда овариэктомированные, но получающие эстроген птицы в условиях короткого и длинного светового дня, по сравнению со временем, которое необходимо для постройки гнезда
| Измеряемый показатель |
Световой день 8 ч; темнота 16 ч |
Световой день 14 ч; темнота 10 ч |
| Имеющееся время для строительства гнезда |
1,1 ч |
7,5 ч |
| Необходимое для строительства гнезда время |
2,3 ч |
2,3 ч |
Результаты многочисленных исследований показали, что функция спроса у животных подчиняется такому же общему закону, что и функция спроса у человека. Например, Ли и Поупер обнаружили, что спрос на пищу был эластичным у крыс, которым приходилось совершать определенную работу для получения пищевого подкрепления. Если от крыс требовалось совершить больше работы, то крысы продолжали работать, получая примерно такое же количество пищи. Однако эти исследователи обнаружили также и то, что эластичность спроса на пищевые шарики увеличивалась, если они были сделаны из сахарозы.
Параллели между явлениями спроса у животных и человека - это тема, которая вызывает сейчас большой интерес. Эластичность функций спроса дает представление об относительной важности тех или иных благ, на которые человек тратит свои деньги. Существует тесная взаимосвязь между эластичностью спроса и упругостью поведения. Таким образом, функции спроса можно использовать как непрямую меру упругости. Если активность А имеет более высокую упругость, чем активность В, то функция спроса для активности А будет характеризоваться относительной неэластичностью, а для активности В - эластичностью.
|