Основы антропологии
Содержание
1. Современные и ископаемые приматы (до гоминид) и тенденции их эволюции
2. Проблемы происхождения человека (антропогенез)
3. Ранние стадии гоминид. Дриопитеки, рамапитеки, австралопитеки
4. Факторы и критерии гоминизации
5. Кроманьонский человек. Человек современного типа
6. Время и место формирования человека разумного
7. Индивидуальное развитие человека
8. Конституции человека
9. Эволюция мозга
10. Социальные факторы происхождения человека
11. Строение тела и характер
12. Видовое единство человека. Понятие о расе
13. Социальные корни расизма. Научная несостоятельность расистских теорий
14. Экология дифференциации человека
15. Этническая антропология
16. Классификация рас по Деникеру
17. Правила Бергмана, Аллена, Глогера, Томсона-Бакстона
18. Классификация рас по группам крови по Бойду
19. Природные факторы расообразования
20. Гипотеза адаптивных типов в расогенезе
21. Соотношение биологического и социального в становлении личности человека
22. Что изучает антропология?
23. Из каких частей состоит антропология живых современных форм?
24. Где впервые появился человек?
25. Сколько лет тому назад появился человек на Земле?
26. С появлением, какого доисторического человека стала развиваться культура?
27. Кто был предком неандертальца?
28. Кто составил переходную ступень от обезьяны к человеку?
29. Какой объем головного мозга был у китайского синантропа?
30. Кто впервые выделил четыре типа темперамента?
31. По какой формуле исчисляется индекс Пинье?
32. Сколько существует больших рас?
33. Где началось формирование больших рас на Земле?
34. В какой структуре клетки содержится генетическая программа человека?
35. Когда возник человек современного типа на Земле?
36. В какой части света был впервые обнаружен австралопитек?
37. В какие генетические процессы вовлекается ДНК человека?
38. Назовите природные факторы расообразования
39. В какой стране впервые был обнаружен неандерталец?
40. Назовите ответственные этапы в развитии человека
41. Какова была средняя продолжительность жизни первобытного человека?
1. Современные и ископаемые приматы (до гоминид) и тенденции их эволюции
Основные особенности приматов
- Пятипалая хватающая конечность
- Замена когтей ногтями
- Способность к вращательным движениям в локтевом суставе (пронации, супинации)
- Значительный объем движений плеча
- Увеличение головного мозга
- Всеядность
- Наличие 2-х генераций зубов: молочной и постоянной
- Высокий уровень развития ВНД
В настоящее время численность приматов быстро сокращается (69 видов и подвидов в Красной книге МСОП). Все перечисленные признаки отчетливее выражены у высших приматов по сравнению с низшими. Отряд приматов классифицируют на подотряд полуобезьян (prosimii) и обезьян (antropoidea).
40 млн лет назад обезьяны (antropoidea) в результате эволюции дали начало трем большим группам (надсемействам) живых существ:
Iгруппа: надсемейство geboidea, от которого появились в ходе эволюции обезьяны Нового Света (широконосые), эволюционировавшие в игрунков и капуцинов.
IIгруппа: надсемейство cercopithcoidea. Его представители дали начало обезьянам Старого Света (узконосым), которые эволюционировали в макак и павианов.
IIIгруппа: надсемейство hominoidea (гоминиды). От них произошли человекообразные обезьяны и человек. Этих обезьян (понгид) делят на гиббонов, орангутанов, горилл и шимпанзе.
По мнению Ч.Дарвина, общим предком человека, горилл и шимпанзе был дриопитек, возраст которого составляет около 25 млн лет. У истоков расхождения путей эволюции понгид и гоминид стоял один из дриопитеков, названный в честь индийского бога Рамы - рамапитек, челюсть которого была найдена Д.Льюисом в 1934 г. в Индии. Возраст рамапитека определен в 14 млн лет. Его рост составлял около 1 м, питался он зернами и корнями, мог частично передвигаться на задних конечностях, но большую часть времени проводил на деревьях.
Эволюция приматов до человека
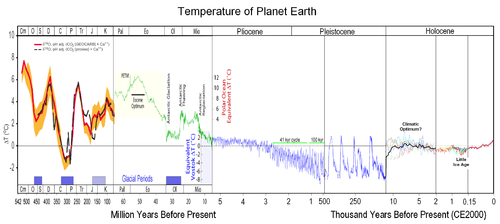
Изменения средней температуры Земли за последние 500 млн лет
Приматы — одна из старейших групп современных плацентарных млекопитающих. Эволюционная история приматов может быть прослежена примерно на 90 млн лет назад, когда Приматоморфы разделились на Приматов и Шерстокрылов. 85 млн. лет назад Сухоносые обезьяны отделились от Мокроносых. Около 80 млн. л.н. разошлись линии Долгопятообразных и Обезьянообразных, а Лемуры отделились от Лориобразных. Останки древнейших приматов известны лишь из палеоценовых и эоценовых отложений Северной Америки, Евразии и Африки (Плезиадаписы, Антрасимия, Нотарктус, Дарвиниус (Ида) и др). Только для Пургаториуса предполагается верхнемеловое время.
После глобального похолодания, когда около 30 млн лет назад, в раннем олигоцене, Антарктида начала покрываться льдом, приматы вымерли повсеместно кроме Африки, Америки и юга Азии. Одним из выживших был Грифопитек — ископаемая обезьяна, жившая на территории современной Германии и Турции около 16,5 млн лет назад, на 1,5 млн лет раньше, чем подобные виды появились в Африке. Возможно, первые человекообразные обезьяны также появились не в Африке, а в Евразии. С другой стороны, высказываются предположения, что предки гоминид мигрировали в Евразию из Африки около 17 млн лет назад, когда эти континенты некоторое время были соединены, прежде чем вновь разделились при расширении Средиземного моря. В начале миоцена климат снова стал тёплым, и они могли процветать в Евразии, после чего ареал обитания одного из них, дриопитека, распространился из Европы или западной части Азии в Африку[3]
.
Возможно, что уцелевшая после климатических изменений, тропическая популяция обезьян, хорошо представленная в слоях верхнего эоцена и нижнего олигоцена Файюмского оазиса в Египте (биретия, каранисия, сахарогалаго, протеопитек, египтопитек, катопитек и др.), дала начало всем ныне существующим приматам — лемурам Мадагаскара, лори Юго-Восточной Азии, африканским галаго, широконосым обезьянам Нового Света и узконосым обезьянам Старого Света (человекообразные обезьяны и мартышкообразные).
В раннем миоцене примитивные узконосые обезьяны Восточной Африки пережили длительный период видообразования. Среди выделяемых ныне видов и родов обезьян этой эпохи известны, в частности, камойяпитек, моротопитек, лимнопитек, проконсул, афропитек, кениапитек, чорорапитек, экваториус, отавипитек, ньянцапитек, викториапитек, дендропитек, все из Восточной Африки, а также ореопитек, который обитал в Италии около 9 млн лет назад и европейские — пиеролапитек, анойяпитек, дриопитек. При сравнении ДНК современных обезьян было показано, что гиббоны отделились от общего ствола гоминид ~ 18 млн лет назад, а орангутаны — около 14 млн лет назад. За исключением дендропитека останки ископаемых гиббонов науке неизвестны, и их происхождение остается неясным. Ископаемым прото-орангутаном считается сивапитек, обитавший в Азии около 12—10 млн лет назад и хорапитек из Таиланда.
Предполагается, что видами, близкими к общему предку горилл, шимпанзе и людей, были накалипитек из Кении и уранопитек с Балканского полуострова. По данным молекулярной биологии, около 7 — 8 млн лет назад сначала гориллы, а потом и шимпанзе отделились от предков людей. ДНК шимпанзе на 98,4 % идентична человеческой. Из-за влажного климата тропических лесов, в кислых почвах которого кости плохо сохраняются, а также отчасти из-за невнимания исследователей, сосредоточенных преимущественно на поиске предков человека, ископаемых горилл и шимпанзе до сих пор практически не найдено.
В середине миоцена климат снова стал холоднее и засушливее, что спровоцировало новое массовое вымирание фауны. Однако гоминины, как и многие другие виды (антилопы, гиены, собаки, свиньи, слоны, лошади) пережили климатические изменения и сумели к ним адаптироваться. Их дальнейшая эволюция привела к образованию множества новых видов, из которых самыми древними сейчас считаются Sahelanthropus tchadensis (7 млн лет назад) и Orrorin tugenensis (6 млн лет назад). За ними, в частности, последовали:
Ardipithecus (5.8-4.4 млн лет назад), с видами Ar. kadabba и Ar. ramidus;
Australopithecus (4-2 млн лет назад), с видами Au. anamensis, Au. afarensis, Au. africanus, Au. bahrelghazali и Au. garhi;
Paranthropus (3-1.2 млн лет назад), с видами P. aethiopicus, P. boisei и P. robustus;
Kenyanthropus (3 млн лет назад), вид Kenyanthropus platyops
Homo (от 2 млнлетназад), свидамиHomo habilis (или Australopithecus habilis), Homo rudolfensis, Homo ergaster, Homo georgicus, Homo antecessor, Homo cepranensis, Homo erectus, Homo heidelbergensis, Homo rhodesiensis, Homo neanderthalensis, Homo sapiens idaltu, Homo sapiens sapiens, Homo floresiensis.
2. Проблемы происхождения человека (антропогенез)
Происхождение человека или антропогенез.
Родиной европейской науки является древняя Греция. Возраст науки насчитывает 2000 лет и 12 тысяч научных дисциплин.
Антропология (от греч. Antropos (антрос) — человек, и logos (логос) — разум, знание) — наука о происхождении и эволюции человека. Термин введен Аристотелем.
Предмет включает в себя всезнание от анатомии до истории. Особенно длительная связь у антропологии отмечается с психологией и медициной.
Сам предмет имеет 2-а специфических метода: антропометрия и антропоскопия. Также широко используются смежные науки: физика, физиология, биология, генетика, медицина и ряд других наук.
Антропология состоит из 2-х разделов:
1 раздел: палеоантропология – антропология ископаемых форм;
2 раздел: антропология живых современных форм.
Первый раздел палеоантропология состоит их 4-х частей:
1) происхождение приматов;
2) происхождение человека - антропогенез;
3) происхождение человеческих рас – расогенез;
4) наука о болезнях ископаемого человека – палеопатология.
Второй раздел антропология живых современных форм состоит из 7-ми частей:
1) соматология (от греч. soma, родительный падеж somatos — тело и логия);
2) физическая антропология;
3) анатомическая антропология;
4) этническая антропология;
5) антропогенетика;
6) дерматоглифика (от греч. Derma (дерма), род. падеж dermatos кожа и glypho (глифика) вырезаю, гравирую, узор);
7) прикладная антропология.
Организм человека имеет много общих признаков, как с позвоночными, так и с млекопитающими. Например, позвоночник, 7 шейных позвонков, 2-е пары конечностей, замкнутая кровеносная система (2-а круга кровообращения: большой и малый круги), левая дуга аорты, 4-х камерное сердце, безъядерные эритроциты, волосяной покров, постоянная t тела, легочное дыхание, наличие диафрагмы, нервной системы, органов чувств и др.
Особенно велико сходство с приматами.
Научная классификация человека: по критериям зоологической систематики вид Homosapiens (Человек разумный) относится к царству животных (Animalia), типу хордовых (Chordata), классу млекопитающих (Mammalia), отряду приматов (Primates), семейству гоминид (Hominidae), вид разумный.
Впервые человека выделил из животных в отдельный ряд Карл Линней в 1758 году.
На латыни Homo sapiens (хомо сапиенс) - человек думающий (человек разумный).
Современная классификация отряда приматов включает около 200 видов, которые группируются в 50 родов.
Основные особенности приматов:
1. пятипалая хватательная конечность;
2. замена когтей – ногтями;
3. способность к вращательным движениям в локтевом суставе – супинация (ладонь кверху, ладонь вниз);
4. значительный объем движения плечом;
5. значительный размер головного мозга.
В среднем у современного человека мозг весит в кубических сантиметрах 1,5 кг. (1 кубический сантиметр = 1 грамму). Чем больше серого вещества (нейронов), тем умнее человек, а белое вещество – это проводящие пути. Левое полушарие отвечает за логику и интеллект, а правое полушарие – за гуманитарию.
Высшее положение в отряде приматов занимает семейство гоминоидов, которое объединяет человека и человекообразных обезьян (гиббоны и крупные человекообразные обезьяны – орангутанг, гориллы, шимпанзе).
Человек является высшей ступенью эволюционного развития на Земле, общественным существом, отличительной чертой которого является – сознание.
Сознание сформировалось на основе общественно-трудовой деятельности.
Родиной человечества (метом, где появился человек), считается Африка. Т.к. установлено большое сходство с обезьянами: шимпанзе и горилла по молекулярным, биохимическим и биологическим параметрам. Например:
- по биохимии крови: группы А, В, О;
- по системе: ММ; по системе: РН (резус-фактор);
- по ферментам: изомилаза, плацентарная, щелочная фосфатаза;
- по строению гемоглобина;
- по составу аминокислот и строению ДНК.
Все это доказывает, что человек произошел от обезьян.
Самые древние, достоверные представители рода человеческого известны примерно 2-2,5 мл. лет тому назад. А появился человек современного вида 40-50 тысяч лет назад.
В результате кладогенеза (расщепление) возникло 3 линии гоминид.
Гоминизация – это процесс очеловечивания обезьяны, начало которому положило формирование первых специфических человеческих особенностей, а завершением было возникновение человека современного типа, человека разумного. Соматология (от греч. soma, родительный падеж somatos — тело и логия), отрасль морфологии человека, Соматология (от греч. soma, родительный падеж somatos — тело и логия), отрасль морфологии человека, Соматология (от греч. soma, родительный падеж somatos — тело и логия), отрасль морфологии человека, Это комплексный феномен, включающий развитие прямохождения, увеличение головного мозга, адаптацию (приспособление) к трудовой деятельности руки, изменение зубочелюстного аппарата (строение зубной системы), формирование членораздельной речи, мышления, сознания, интеллекта, перестройки онтогенеза, развитие социальной организации, материальной культуры и т.д. Все эти морфологические признаки в той или иной степени отражают и изменения в поведении. То есть трудовая деятельность, явившись решающим фактором гоменизации, входит в морфологический критерий гоминид опосредованно, через "отпечаток" её в анатомических структурах.
Существует много гипотез происхождения человека, но основных 3.
Первая теория появления человека: эволюционизм.
Считается, что все началось с Рамапитека линии (обезьяна в Индии) от него пошли 3 линии: Сивапитек, Дриопитек и Гиббон.
От Сивапитека пошел Орангутан. От Дриопитека пошли еще 3 линии: Австралопитек, Шимпанзе и Гориллы.
От Гориллы (Гомо) – далее закончилось человеком разумным.
От Австралопитека пошли: Древнейшие люди → Древние люди (Неандертальцы) + (Китайский Синантроп + Яванский Питикантроп) → Кроманьонский человек → Тип современного человека.
Тип современного человека разделился на 3 расы: Черная, Белая и Желтая.
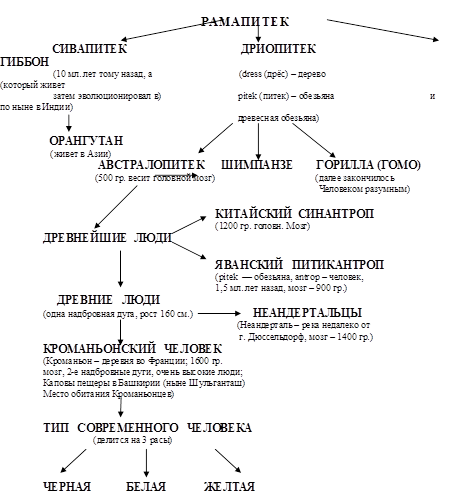
Сейчас из обезьяны не получается человек, потому, что разные генетические линии.
2 теория появления человека: Теория Креационизма - идея Божественного творения (создания) человека.
Креационизм постепенного творения (Progressive creationism): согласно этой концепции, Бог непрерывно направляет процесс изменения биологических видов и их появления. Представители данного направления принимают геологические и астрофизические данные и датировки, однако полностью отвергают теорию эволюции и видообразование путём естественного отбора.
3 теория появления человека: Теория Панспермии (космизм) – из Космоса (в результате прилета инопланетян).
В конце 20 и начале 21 вв. популярность приобрела концепция внеземного происхождения человека и, как кажется, она получает все больше научных доказательств. Миллионы людей отслеживают все случаи появления НЛО, а ряд ученых, современных фантастов, авторов многочисленных теорий пытаются подкрепить гипотетические предположения о внеземном происхождении человека разными аргументами. Например, такими:
1. Человеческое существо с самого своего рождения абсолютно не приспособлено к существованию в земных условиях. Если верить Дарвину, самые близкие нам родичи - приматы. Но у них все по-иному, не по-человечески. У шимпанзе срок беременности - 7, а у гориллы - 9,5 месяца, они рождают детей вполне доношенных, но весом всего 2 кг. При этом самка шимпанзе становится взрослой к 8 годам, самец - к 10.
2. Математиков анализ ситуации привел к мысли, что если бы сила тяжести на Земле составляла 0,6 от существующей, то человек, подобно кошке или собаке, мог бы падать сколько угодно, не причиняя себе заметных травм, не говоря уже о переломе костей! Может быть, поэтому человеческое дитя так страдает, плачет, капризничает, болеет? Ему невыносимо тяжело, больно, неуютно на этой чужеродной Земле, которая влияет на него своим тяготением. Значит, на планете-прародительнице, откуда предположительно когда-то переселились наши прапредки, было планетарное тяготение 0,6 от земного?
В мифологии западноафриканского племени догонов содержится недвусмысленное указание на прибытие их предков из другой звездной системы, расположенной близ Сириуса. Считается вполне вероятным, что сам Сириус находился когда-то вблизи Солнца, составляя с ним (что не противоречит законам небесной механики) двойную звездную систему.
Вполне возможно, что в какой-то период звезд было не две, а несколько. Впоследствии Сириус отклонился в своем движении от первоначального пути во Вселенной, и его занесло в тот участок нашей Галактики, где он в настоящее время и пребывает.
Первые расы появились 100 тыс. лет назад.
В Азии сформировались 2 расы: Восточная (желтая) и Юго-западная (белая и черная).
Развитие взглядов на происхождение человека
Одна из интереснейших и сложнейших тем, изучаемых в курсе общей биологии, – происхождение человека. Где, когда и как возник род человеческий? Как он расселился по Земле?
В прошлом веке в европейской культуре существовало два ответа: один дан в Библии, другой – в теории Чарлза Дарвина. И хотя происхождение человека от обезьяноподобного предка было одним из частных следствий дарвиновской теории эволюции, естественно, именно этот вопрос – создан ли человек Богом или произошел от обезьяны – привлек внимание широкой публики.
Наиболее ортодоксальные сторонники библейской версии считают, что каждый вид, в том числе и человек, был создан Богом. Область исследований, направленных на поиск научных доказательств этой версии, называется креационизмом. Современные креационисты подтверждают тексты Библии точными расчетами. В частности, они доказывают, что Ноев ковчег мог вместить всех тварей по паре – если учесть, что рыбам и другим водным животным место в ковчеге не нужно, а остальных позвоночных животных – около 20 тыс. видов. Если умножить это количество на два (в ковчег были взяты самец и самка), получится примерно 40 тыс. животных. Средней величины автофургон для перевозки овец вмещает 240 животных. Значит, нужно было бы 146 таких фургонов. А ковчег длиной 300, шириной 50 и высотой 30 локтей вместил бы 522 таких фургона. Значит, место для всех животных нашлось и еще осталось – для корма и людей. Тем более что Бог, как утверждает Томас Хайнц из Института креационных исследований, наверняка догадался бы взять маленьких и молодых животных, чтоб они и места занимали меньше, и размножались активнее.
Чарлз Дарвин не отрицал существования Бога, однако считал, что Бог создал лишь начальные виды, остальные же возникли под действием естественного отбора. Альфред Уоллес, который пришел к открытию принципа естественного отбора почти одновременно с Дарвином, в отличие от последнего утверждал, что между человеком и животными существует резкая грань в отношении психической деятельности. Он пришел к выводу, что мозг человека не может рассматриваться как результат естественного отбора. Уоллес провозгласил, что этот "мыслительный инструмент" возник в результате потребностей его обладателя, и предполагал "вмешательство высшего разумного существа".
Человек – это животное, интересующееся своим происхождением. Интерес к собственному происхождению свойствен человеку с глубокой древности. Чем дольше ученые изучают палеонтологическую летопись, тем яснее вырисовывается картина превращения обезьяны в человека. Современные представления гораздо сложнее излагавшейся в наших старых учебниках "единственно верной" теории, которую можно резюмировать словами Абрама Терца (Синявского): "Обезьяна встала на задние лапы и пошла прямым путем к коммунизму".
По пути гоминизации шли многие виды приматов, и Homo sapiens в момент своего появления был просто представителем одной из нескольких конкурирующих линий. То, что именно он достигнет успеха на арене эволюции, не было предопределено. Высыпая ведро песка, вы не можете предсказать, какая из песчинок окажется сверху. Однако ясно, что какая-нибудь из них займет место наверху, чуть-чуть выше других. То же происходит и в процессе эволюции: один из близких видов оказывается чуть-чуть успешнее, чем другие, но какой именно – заранее предсказать нельзя.
Сегодня большинство ученых придерживаются теории африканского происхождения человека и считают, что будущий победитель в эволюционной гонке возник на Юго-Востоке Африки около 200 тыс. лет назад и расселился оттуда по всей планете (рис. 1).
Раз человек вышел из Африки, то, казалось бы, само собой разумеется, что наши дальние африканские прародители были похожи на современных жителей этого континента. Однако некоторые исследователи считают, что первые люди, появившиеся в Африке, были ближе к монголоидам.
Монголоидная раса имеет ряд архаичных черт, в частности в строении зубов, которые характерны для неандертальцев и Homo erectus (Человека прямоходящего). Популяции монголоидного типа обладают высокой адаптивностью к различным условиям обитания, от арктической тундры до экваториальных влажных лесов, тогда как у детей негроидной расы в высоких широтах при недостатке витамина D быстро возникают заболевания костей, рахит, т.е. они специализированы к условиям высокой инсоляции. Если бы первые люди были подобны современным африканцам, то сомнительно, что они смогли бы успешно осуществить миграции по всему земному шару. Однако эта точка зрения оспаривается большинством антропологов.
Концепции африканского происхождения противопоставляется концепция мультирегионального происхождения, предполагающая, что наш предковый вид Homo erectus превратился в Homo sapiens в различных точках земного шара независимо.
Homo erectus появился в Африке около 1,8 млн лет назад. Он изготавливал каменные орудия, найденные палеонтологами, и, возможно, более совершенные орудия из бамбука. Однако от бамбука через миллионы лет не остается следов. За несколько сотен тысяч лет Homo erectus распространился сначала по среднему Востоку, затем в Европу и до Тихого океана.
Самый близкий родственник человека был открыт в 1856 г. в местечке Неадерталь возле Дюссельдорфа. Рабочие, нашедшие пещеру со странными черепами и большими костями, решили, что это останки пещерного медведя, и даже не предполагали, какие жаркие споры вызовет их находка. Эти кости, а также кости, найденные позже на севере Англии, на востоке Узбекистана и на юге Израиля, были останками предка человека, получившего название неандерталец, – примитивный человек, живший от 200 000 до 27 000 лет назад. Неандерталец делал примитивные орудия, раскрашивал тело узорами, имел религиозные представления и похоронные ритуалы.
Предполагается, что неандерталец эволюционировал из Homo erectus в Европе и вымер, не способный конкурировать с пришедшим из Африки Homo sapiens. Причиной вымирания могла быть и слишком высокая специализация – неандертальцы были приспособлены к жизни в условиях ледниковой Европы. При изменении условий такая специализация обернулась для них бедой.
Проблема происхождения человека представляет большой научный интерес и вызывает жаркие споры среди ученых. Существует пять основных концепций происхождения человека: креационизм, космическая, биологическая, трудовая и мутационная концепции. Социальные факторы антропогенеза (трудовая деятельность, общественный образ жизни, речь и мышление) приобрели большое значение в эволюции человека, что позволяет говорить об антропосоциогенезе. Это означает, что становление социальной формы движения не отменило действия биологических законов, а лишь изменило их проявление. Сравнивая человека и животных, мы можем говорить как о сходствах, так и о различиях. К основным сходствам можно отнести одинаковый внешний состав, строение и поведение, наличие у человека рудиментов и атавизмов. Самым же главным отличием можно считать понятийное мышление, речь, труд. Современные биологи и антропологи полагают, что процесс биологической эволюции человека как вида, т.е. его видообразования, прекратился со времени появления homo sapiens. Значит, эволюция на своременном этапе связана с социальной стороной, следовательно, будущее человека зависит от состояния нашей культуры и образования.
Происхождение человека Ответ на вопрос о том, как появился и формировался человек, иными словами, как шел антропогенез, был предметом ожесточенных дискуссий на протяжении веков и тысячелетий. Изучением процесса антропогенеза занимается антропология - общее учение о происхождении и эволюции человека, образовании человеческих рас и вариациях физического строения человека. С точки зрения естествознания необходимо, прежде всего, решать вопрос о происхождении и эволюции вида Homo sapiens, в определении его места в животном мире, выявлении его родства с другими приматами, а также выяснения отличий человека от них. В настоящее время на эти вопросы пытаются дать ответы пять основных концепций о происхождении человека: 1. Креационизм - человек сотворен Богом или мировым разумом. 2. Биологическая концепция - человек произошел от общих с обезьянами предков путем накопления биологических изменений. 3. Трудовая концепция - в появлении человека решающую роль сыграл труд, превративший обезьяноподобных предков в людей. 4. Мутационная концепция - приматы превратились в человека вследствие мутаций и иных аномалий в природе. 5. Космическая концепция - человек как потомок или творение инопланетян, в силу каких-то причин попавших на Землю. Креационизм в антропогенезе Эта концепция возникла исторически первой, так как религия появилась задолго до науки, и первые ответы на вопросы о происхождении и сущности человека были даны именно религией. В зависимости от уровня развития религии можно выделить несколько вариантов ответов на этот вопрос. Так, у бесписьменных народов, обычно обладающих развитыми тотемистическими мифами, рассказывается о том, как тотемный предок (обычно это или растение, или животное, или какой-то неодушевленный предмет), превратился в первого человека и дал начало их роду. например, у австралийцев, считающих себя потомками ящерицы, говорится, как их тотемный предок пришел с севера, и нашел там самозародившиеся беспомощные человеческие зародыши со склеенными пальцами и зубами, закрытыми ушами и глазами. Он каменным ножом отделил эти зародыши друг от друга, открыл им глаза и уши, научил добывать огонь, готовить пищу и т.д. В языческих религиях, основанных на вере во множество богов, олицетворяющих силы природы, человек считается творением этих богов. Так, древние шумеры считали, что первые люди были созданы богами из глины, чтобы те служили им. Иногда люди могли быть прямыми потомками богов. Например, древние греки возводили родословную многих своих героев к олимпийским богам, чаще всего к Зевсу. В монотеистических религиях, таких как христианство или ислам, существует лишь единый Бог, который считается творцом мира и человека. Так, в Библии, священной книге христиан, рассказывается, что на 6ой день творения Бог сотворил человека по образу и подобию своему, чтобы тот владел всей Землей. Он сделал Адама из праха земного и вдохнул в него дыхание жизни. Затем из ребра Адама он создал Еву. Коран рассказывает о том, как Аллах создал человека из эссенции глины, превратил ее в каплю, создал из нее сгусток крови, затем кусок мяса и кости. После этого он облек кости мясом и вырастил человека в другом творении. Подобные утверждения могут быть лишь предметом веры, но не научного доказательства. Тем более, что был обнаружен ряд фактов, не укладывающихся в религиозную картину мира. Среди них - обнаружение останков живых существ, имеющих сходство с человеком, но не являющихся людьми. Поэтому с момента возникновения науки ученые начали искать другие объяснения происхождению человека. Космическая концепция По степени доказательства эта точка зрения стоит ближе всего к концепции креационизма. Возникла она в 60е годы 20 в. под влиянием успехов космонавтики и растущей популярности научно-фантастической литературы. Сторонники этой концепции исходят из того, что человек на Земле появился в результате вмешательства инопланетян. Это могло быть либо осуществление заранее намеченной цели - распространение разума на подходящих планетах, либо результатом несчастного случая - аварии космического корабля инопланетян, заставившего их остаться на Земле, и их последующего одичания. Наиболее подробна космическая концепция была изложена Э. Фон Дэникеном в книге "Воспоминания о будущем" По его мнению, несколько млн. лет назад посланцы внеземной цивилизации посетили нашу планету и, руководясь своими целями, внесли изменения в генетический аппарат гоминид, вызвав, таким образом, появление человека. Позже они еще несколько раз посещали нашу планету, чтобы проконтролировать и помочь своим питомцам. Для доказательства этой точки зрения в книге приводится множество свидетельств палеоконтактов. Биологическая концепция Предпосылки, которые привели к формированию этой концепции, появились в эпоху Великих географических открытий, сделавших доступным для наблюдений и исследований практически весь мир. Начиная с 15 в., в мире господствовало мнение, что народы отличаются по уровню своего развития, потому что такими их создал Бог. но сначала 19в. в естествознание начинает проникать идея всеобщей связи и развития, которая в антропологии трансформировалась как представление о развитии человека, о том, что народы могут находиться на разных ступенях общественного прогресса. основная заслуга в этом принадлежит Тайлору, который в сер. 19в. разработал основные положения классического эволюционизма - теории, утверждающей единство человеческого рода, развивающегося по единым законам. Тогда же было высказано предположение, что разные народы движутся с разной скоростью, но путь, который им предстоит пройти, один. Параллельно с этими исследованиями появились и первые научные представления о происхождении человека, являющегося результатом животного мира Земли. Впервые эти идеи были высказаны К.Линнеем, который в своей "Системе природы" отнес человека к животному миру, отвел ему место рядом с человекообразными обезьянами. Во 2ой половине 18в. Ж. Бюффон и П. Кампер показали глубокое сходство в строении основных органов человека и животных, особенно высших обезьян, заложив основы научной приматологии. К 19в. в науке сложилось теоретические предпосылки появления теории антропосоциогенеза. Важнейшей из них стало проникновение идей эволюционизма в биологию. Решающий, подлинно революционный шаг был сделан Ч.Дарвином, который в 1871г. опубликовал свою книгу "Происхождение человека и половой отбор". В ней на громадном фактическом материале Дарвин обосновал два важных положения: - человек произошел от животных предков; - человек состоит в родстве с современными человекообразными обезьянами. Так возникла симиальная (обезьянья) концепция антропогенеза, согласно которой человек и современные антропоиды произошли от общего предка, жившего в отдаленную геологическую эпоху и представлявшего собой ископаемое африканское обезьяноподобное существо. Во 2ой половине 20 в. с помощью биохимии, физиологии, иммунологии и генетики удалось получить и другие доказательства родства человека с животным миром нашей планеты. Прямым доказательством родства человека и обезьян стали останки ископаемых существ - как общих предков человека и обезьян, так и промежуточных форм между обезьяньим предком и современным человеком. По последним данным, появление высших обезьян, давших начало обезьяньим предкам человека, произошло около 20 млн. лет назад. Судя по всему, в то время существовало около 20-30 видов антропоидов. Большинство из них вымерло, а переход к человеку осуществил лишь один вид. Итак, древнейший предок человека и высших обезьян - рамапитек, живущий на территории от Индии до Африки. Рамапитек -- останки ископаемой крупной обезьяны, по строению зубов оказавшейся промежуточной между современными человекообразными обезьянами и человеком. Жил около 14--8 млн. лет назад. Австралопитеки
Австралопитеки - самые древние, переходные от обезьян к человеку формы, найденные в Южной Африке и вымершие свыше 2 млн. лет назад. Эволюция их продолжалась 8 млн. лет. Австралопитеки ходили на двух ногах при выпрямленном положении тела, жили в пещерах, на безлесных пространствах. Их челюсти были слабо развиты. В качестве орудий для ударов использовали камни, палки, крупные кости конечностей копытных животных, для резки и рубки применяли нижние челюсти и лопатки тех же животных. Австралопитеки были всеядными. Концевые фаланги пальцев у австралопитеков короткие и плоские, как у современного человека. Человек умелый. Древнейшие люди (архантропы)
Древнейшие люди (архантропы) - ранняя стадия становления человека. Предками их были различные ветви вида "Человек умелый": представлены значительным числом форм. Внешне они были очень похожи на современного человека, хотя строение черепа имело такие примитивные черты, как низкий, покатый лоб, мощное развитие надбровного валика, отсутствие подбородочного выступа; масса мозга достигала 1000г, но форма его также была примитивной. Рост мужчины был 160см. По-видимому, им была свойственна примитивная речь. Речь, умение мыслить словами называют второй сигнальной системой и характерна она только для человека. Древнейшие люди освоили огонь. Они далеко расселились на территории Старого Света. Время их существования 1,5 млн. -- 200 тыс. лет. Древние люди (палеоантропы)
Древние люди (палеоантропы) - жили 200--400 тыс. лет назад. Представители этой стадии названы неандертальцами. Название этого вида связано с долиной Неандерталь в Германии (XIX в.), где впервые были найдены останки этих людей. По времени с эпохой неандертальцев совпала эпоха Великого оледенения. При большой массе мозга и черепа неандертальцы обладали еще примитивными чертами: низкий лоб и затылок, большой надглазничный валик, слабое развитие подбородочного выступа. Речь, по-видимому, была еще нечленораздельной. Этот вид с начала своего возникновения дал две ветви эволюции. Одна была представлена людьми звероподобными, очень крупными, но по строению бывшими ближе к древнейшим людям: они являлись тупиковой ветвью эволюции. Люди другой ветви были меньше ростом и менее развиты физически, но по морфологическим признакам были ближе к современному человеку. Они жили большими стаями, у них было разделение труда между мужчинами и женщинами. Речь была еще примитивна, но логическое мышление уже было развито. "Человек разумный" (неоантроп) -- существует 40 тыс. лет. Представителем неоантропов является кроманьонец. Первая находка была сделана на юге Франции близ местечка Кро-Маньон. Они характеризуются небывалым психическим развитием. Главной чертой было развитие интеллекта, способности к искусству. Они охраняли слабых, женщин, детей, стариков -- и этим обеспечили свое процветание. Важнейший вклад этих людей в историю человечества -- приручение ряда животных и развитие земледелия, выведение культурных растений. После этого эволюция человека вышла из-под определяющего влияния биологических факторов и приобрела социальный характер. Мутационная концепция антропогенеза Человек выделился из животного мира благодаря появлению способности к труду и сознательной деятельности. Как считает современная наука, возникновение таких необычных способностей у предчеловека должно было опираться на какие-то биологические предпосылки, создавшие возможность такого скачка. На этой основе возникли предпосылки о роли мутаций у предчеловека в наследственном аппарате. Причины появлений мутаций такого рода могли быть самыми разными. Одной из возможных причин стало влияние ближнего космоса и солнечной активности. Воздействие ближнего космоса шло по 2м направлениям: через изменение среды обитания древних обезьян, прежде всего ландшафта и климата, и через трансформацию самой жизни, проявлявшуюся через мутации, естественный отбор и борьбу за существование. Влияние солнечной активности на земные процессы и явления было доказано российским ученым А.Чижевским еще в 30е г.г. 20в. Современная наука согласна с ним в том, что периодические колебания магнитного поля Солнца влияют на нервную систему и сердечную деятельность и в прошлом могли привести к мутациям древних обезьян. В качестве еще одной причины мутаций ученые называют периодическую смену магнитных полюсов Земли. На протяжении последних 4 млн. лет северный и южный полюса менялись 4 раза. Но в период смены полюсов магнитосфера Земли, защищающая биосферу от космических лучей, намного ослабевает. Это приводит к увеличению ионизирующей радиации на 60% и в 2 раза увеличивает частоту мутаций в зародышевых клетках гоминид. Автор этой теории - Г.Н.Матюшкин. Дополнительным фактором, вызвавшим появление непосредственных предков человека, стала геологическая активность Земли и радиация снизу. Так, в Восточной Африке, признанном месте появления человека, около 20 млн. лет назад образовались трещины в земной коре, в результате чего на поверхности обнаружились залежи урановых руд. Они интенсивно влияли на приматов, живших в пещерах, расположенных вблизи, вызывая мутации разного рода. В 1960-е годы ученым удалось выяснить, что источником генетических мутаций может быть не только радиация, но и стресс, запредельные нервные нагрузки. Мутанта, ставшего предком человека, в обычных условиях ждала неминуемая гибель. Но он нашел замену всему тому, что у него отобрала природа. Он стал использовать искусственные орудия труда, жить в обществе, творить культуру. Иными словами, он стал человеком.
Трудовая концепция Трудовая концепция отвечает на вопрос о причине превращения обезьяноподобных предков в человека. По словам Ф.Энгельса, труд создал человека. Таким образом, важнейшими шагами в ходе эволюции человека стали прямохождение, а также развитие руки и мозга. Они привели его к очеловечиванию. Переход к прямохождению был вызван наступлением ледникового периода. Гоминидные предки не ушли к экватору, а стали приспосабливаться к новым суровым условиям. При этом предки австралопитеков передвигались в вертикальном положении, что освобождало их верхние конечности для защиты и охоты. Кроме того, поскольку перемещение на двух ногах сложнее, чем на четырех конечностях, стала развиваться психика. Это в свою очередь повлекло изменение анатомического строения мозга, его объем составлял 522см кубических. Итак прямохождение высвобождало передние конечности и постепенно превращало их в руку - орган трудовой деятельности. Изменение головы и глаз привело к возрастанию зрительной информации. Естественный отбор шел в направлении закрепления тех качеств, которые помогали приспособиться к новым условиям жизни. Так совершенствовалось прямохождение, устойчивость тела, развивались верхние конечности. В ходе отбора рука научилась использовать камни и дубины, кости и зубы убитых животных для защиты от нападения, для добычи пищи. Человеческие предки стали не только использовать, но и производить орудия труда. В своб очередь, труд обусловил зарождение и развитие социальных отношений, сознания, мышления, то есть окончательно превратил животное в человека. Сходства и отличия человека от животных Для рассмотрения проблемы антропогенеза необходимо уточнить, каково место человека в структуре живого. Отсюда вытекают основные сходства и отличия человека от животных. Они состоят в следующем. Сходство человека с животными определяется, во-первых, вещественным составом, строением и поведением организмов. Человек состоит из тех же белков и нуклеиновых кислот, что и животное, и многие структуры и функции нашего тела такие же, как и у животных. Чем выше на эволюционной шкале стоит животное, тем ближе его сходство с человеком. Во-вторых, человеческий зародыш в своем развитии проходит те же стадии, которые прошла эволюция животного. И, в-третьих, у человека имеются рудиментарные органы, которые выполняли важные функции у животных и сохранились у человека, хотя сейчас не нужны ему (например, аппендикс, копчик и др.). Появление в процессе эмбрионального развития человека хорды, жаберных щелей в полости глотки, дорсальной полой нервной трубки, двухсторонней симметрии в строении тела определяют принадлежность человека к типу хордовых. Развитие позвоночного столба, сердце на брюшной стороне тела, наличие двух пар конечностей -- к типу позвоночных. Теплокровность, развитие молочных желез, наличие волос на поверхности тела свидетельствуют о принадлежности человека к классу млекопитающих. Развитие детеныша внутри тела матери и питание плода через плаценту определяют принадлежность человека к подклассу плацентарных. Множество более частных признаков четко определяют положение человека в системе отряда приматов. Итак, с биологической точки зрения, человек - один из видов млекопитающих, относящихся к отряду приматов, подотряду высших. Однако и отличия от животных фундаментальны. К ним, прежде всего, относится разум. Самые высшие животные не обладают способностью к понятийному мышлению, т. е. к формированию отвлеченных, абстрактных представлений о предметах, в которых обобщены основные свойства конкретных вещей. Мышление животных, если о таковом можно говорить, всегда конкретно; мышление человека может быть абстрактным, отвлеченным, обобщающим понятийным, логичным. Благодаря способности к понятийному мышлению человек сознает, что он делает, и понимает мир. Вторым главным отличием является то, что человек обладает речью. У животных может быть очень развитая система общения с помощью сигналов, но только у человека есть вторая сигнальная система - общение с помощью слов. В естествознании предполагается, что речь произошла из звуков, произносимых при работе, которые потом становились общими в процессе совместного труда. Таким же путем в процессе общественного труда постепенно мог возникнуть разум. Способность к труду - еще одно фундаментальное отличие человека от животных. Только человек способен изготовлять, творить орудия труда. С этим связаны утверждения, что животные приспосабливаются к окружающей среде, а человек преобразует ее и что в конечном счете, труд создал человека. Со способностью к труду соотносятся еще два отличительных признака человека: прямохождение, которое освободило его руки и как следствие, развитие руки, особенно большого пальца на ней. Наконец, еще два характерных признака человека повлиявших на развитие культуры - использование огня и захоронение трупов. Таким образом, главные отличия человека от животных - понятийное мышление, речь, труд - стали теми путями, по которым шло обособление человека от природы. Биологическое и социальное в историческом развитии человека В настоящем разделе, теперь уже имея представление об антропогенезе, мы остановимся на рассмотрении собственного развития homo sapiens и его свойствах. И первый вопрос, который в связи с этим возникает, можно сформулировать так: а продолжается ли биологическая эволюция человека после возникновения 30--40 тыс. лет назад homo sapiens? Другими словами, как культурная организация жизни людей, включая способы производства, развитие трудовой деятельности, образ жизни и т.д., могла повлиять на биологическую эволюцию человека, и влияла ли она на нее вообще? Строго говоря, эволюция человека продолжается на всем протяжении его существования. Но она относится к социальной стороне жизни. Что же касается биологической эволюции, то с тех пор как человек выделился из животного мира, по крайней мере, она перестала играть решающую роль. Даже люди с ослабленным здоровьем благодаря успехам медицины могут принимать активное участие в жизни общества. Значение естественного отбора резко меняется в жизни человека и животных. Если у животных отбор - это главный фактор эволюции, то у человека его роль заключается в сохранении генофонда, в сдерживании мутаций, отрицательно влияющих на его здоровье. Естественный отбор у человека происходит главным образом на уровне зародышевых клеток. В основном дети рождаются из здоровых в генетическом отношении клеток. Об этом свидетельствует тот факт, что крупные генетические нарушения в половых клетках родителей в подавляющем большинстве случаев обусловливают гибель оплодотворенных яйцеклеток и зародышей на ранних стадиях развития. А меняются ли вместе с социальным обликом человека и его биологическая природа, физический облик, умственные способности? Становятся ли новые поколения людей более развитыми в физическом и умственном плане? Прежде всего, коснемся физического здоровья. Очевидно, что его состояние за историю homo sapiens существенно улучшилось. Комплексным показателем может служить увеличение средней продолжительности жизни населения. Под влиянием социальных условий она возросла с 20--22 лет в древности до 30 лет в XVIII в. К началу XX в. в странах Западной Европы средняя продолжительность жизни была примерно 56 лет. Сегодня в этих странах она достигла 75--78 лет. Теперь обратимся к вопросу о развитии умственных способностей. Один из создателей евгеники (теории о наследственном здоровье человека и путях его улучшения), английский психолог и антрополог Ф. Гальтон, был убежден в том, что интеллект современного человека снижается. По его мнению, представители низших классов обладают более низким коэффициентом интеллектуальности -- IQ. В то же время именно они имеют большее число детей. Причем статистические данные, свидетельствовавшие о более высоком репродуктивном уровне людей с низким IQ, были широко распространены до сравнительно недавнего времени. На основании этих данных Гальтоном и некоторыми другими учеными делался вывод о том, что человеческий вид будет все более наполняться "худшими породами" людей и, следовательно, уменьшать свой IQ. Однако в начале 60-х гг. вывод о tom)-что существует прямая зависимость между социальным положением, количеством детей и IQ, был признан ошибочным. И сегодня нельзя считать доказанным, что коэффициент интеллектуальности у людей обусловлен генетически. На сегодняшний день нет данных, позволяющих говорить об эволюции главного органа мышления - мозга. Косвенно о прекращении эволюции мозга свидетельствует тот факт, что его размеры у homo sapiens остаются неизменными на протяжении примерно 30--40 тыс. лет. А у наших предков они увеличивались постоянно, в течение всей эволюции. Так, у австралопитеков размер мозга составлял 500--600 куб. см, у питекантропов - до 900 куб. см, у синантропов -- до 1 000 куб. см. У современного человека средний размер мозга составляет 1 400 куб. см у мужчин и 1 270 -- у женщин. При этом у человека нет прямой зависимости между величиной мозга и индивидуальной одаренностью. Так, по имеющимся данным, довольно маленький размер мозга среди талантливых людей имел известный французский писатель А. Франс -- не более 1 000 куб. см. А самый большой объем мозга среди них имел И.С. Тургенев -- 2 012 куб. см. Это еще раз доказывает, что на основе различий в объеме мозга не следует делать каких-либо выводов об умственных способностях человека. Современные биологи и антропологи полагают, что процесс биологической эволюции человека как вида, т.е. его видообразования, прекратился со времени появления homo sapiens. Прежде всего, об этом свидетельствует тот факт, что в течение данного периода мозг человека не изменился, морфологическое изменение его завершилось. Для противоположной точки зрения, во всяком случае, нет достаточных оснований. Эволюция связана с социальной стороной, и будущее человека зависит от состояния нашей культуры. В основе эволюции лежит развитие интеллекта и целесообразной деятельности. Необходимо отметить также, что с возникновением человека и общества генетическая информация утрачивает свое главенствующее значение в его жизнедеятельности. Она заменяется социальной информацией. А развитие последней "определяется уже не столько естественным отбором наиболее умелых и одаренных, сколько социальными факторами, которым подчиняется и общебиологический процесс".
3. Ранние стадии гоминид. Дриопитеки, рамапитеки, австралопитеки
Эволюция: к австралопитеку.
Самые первые и наиболее примитивные млекопитающие появились в далекую геологическую эпоху, известную как мезозойская эра, по-видимому, в ее начальный период, называемый триасовым, ок. 200 млн. лет тому назад. В течение более 100 млн. лет эти очень маленькие существа не играли сколь-либо значительной роли; преобладающими животными на громадном отрезке времени были динозавры и другие рептилии. Вероятно, ко времени завершения мелового периода (ок. 75 млн. лет тому назад) на Земле появились наши отдаленные предки – первые приматы. По-видимому, это были мелкие зверьки, питавшиеся травой и плодами и приспособившиеся к древесному образу жизни. Затем мезозойская эра подошла к концу, динозавры вымерли, и началась кайнозойская эра. На протяжении первых двух ее эпох – палеоцена и эоцена, т.е. ок. 40–60 млн. лет тому назад, к семейству Paromomyidae, паромомиид (приматов) добавилось еще два крупных семейства приматов, вероятно древних родственников полуобезьян – лемуров и долгопятов. Точнее говоря, считается, что появление лемуров и долгопятов произошло в конце эоцена.
Эта ранняя стадия эволюции приматов характеризуется тремя важными изменениями: мозг стал значительно крупнее, морда сильно уменьшилась, а передние конечности становились все более приспособленными для хватательных движений, причем вместо когтей появились плоские ногти.
В позднем эоцене обозначилась также линия, ведущая к обезьянам, человекообразным обезьянам и человеку; ее начало связывают с появлением семейства Omomyidae, происходящего от наиболее древних приматов – паромомиид. Формируясь, линия высших приматов, или антропоидов, приобрела некоторые особенности, в частности значительно увеличилась подвижность передних конечностей и возросла зависимость от зрения.
Следующая геологическая эпоха – олигоцен, условно датируемая периодом примерно от 38 до 25 млн. лет тому назад, богата ископаемыми находками, но эти находки трудно связать с эволюцией человека. В это время линия высших приматов начинает разделяться на ветви, идущие в нескольких направлениях: к цебоидам (Ceboidea), или обезьянам Нового света; к церкопитекоидам (Cercopithecoidea), или обезьянам Старого света (мартышкообразным); и к гоминоидам (Hominoidea), т.е. к надсемейству, включающему современного человека, горилл, шимпанзе, орангутанов и гиббонов.
Одна из находок эпохи олигоцена вызвала большие споры в научной среде. Это был парапитек (Parapithecus), найденный в Файюме (Египет). Многие антропологи полагают, что существо такого типа было древним предком современных гоминоидов, другие же полностью исключают саму принадлежность парапитека к приматам. Трудность проистекает из ограниченности имеющихся остатков этого существа – только нижняя челюсть – и из сложности определения его зубной системы (типа и числа зубов). По мнению некоторых антропологов, парапитек родствен амфипитеку (Amphipithecus), обнаруженному в Бирме, хотя его остатки могут датироваться эпохой эоцена. Вкратце, важность этой интерпретации состоит в следующем: если она правильна, то и человекообразные обезьяны, и человек могут восходить напрямую к первым приматам, минуя стадию церкопитекоидов (обезьян Старого света), поскольку у амфипитека и парапитека обнаруживаются антропоидные характеристики на тот момент, когда еще не произошло полного утверждения церкопитекоидов; если же нет, то скорее всего такая стадия существовала.
Вовлеченной в подобного же рода дискуссии оказалась и более поздняя находка, известная как ореопитек (Oreopithecus), сделанная более полувека назад, но ставшая центром новой полемики в 1958, когда были обнаружены новые ископаемые остатки. К сожалению, эта спорная находка дошла до нас в очень плохом состоянии, спрессованной в слое лигнита. Как полагают, ореопитек существовал в нижнем плиоцене ок. 10 млн. лет тому назад. На основании этого нового свидетельства эксперты по анатомии обезьян считают, что находка не принадлежит к церкопитекоидному типу. Если данное мнение подтвердится и ореопитек будет вполне надежно связан с более поздними гоминоидами, это станет дополнительным свидетельством в пользу того, что линия гоминоидов начала обособляться, минуя стадию подобия мартышкообразных.
Обращаясь вновь к эпохе олигоцена, упомянем ряд похожих находок из Европы и Африки. В их числе: проплиопитек (Propliopithecus) и прохилобат (Prohylobates) – оба из Египта, плиопитек (Pliopithecus) – из Франции, и лимнопитек (Limnopithecus) – из Кении. По мнению большинства специалистов, эти ископаемые не относятся к предкам человека, но ведут более или менее прямо к современным гиббонам. Однако, как полагают, они были схожи в важных элементах с каким-то другим современным им приматом – предположительно предком крупных человекообразных обезьян и человека. Эпоха олигоцена длилась ок. 13 млн. лет, и у нас нет возможности датировать ископаемые остатки точнее. Тем не менее очевидно, что олигоцен был периодом лучевого адаптивного расхождения антропоидов.
Из следующей эпохи миоцена до нас дошла чрезвычайно интересная и, по-видимому, связанная с предыдущей группа ископаемых остатков. Это дриопитеки (Dryopithecinae) –широко распространенная группа существ, по размерам сравнимых с шимпанзе и живших в разных частях Африки, Европы и Азии. По некоторым признакам, например по жевательной поверхности моляров, дриопитеков можно расположить на линии происхождения гоминид или, точнее, вблизи той точки, от которой началось расхождение человека и обезьян. Одно из этих ископаемых существ – рамапитек (Ramapithecus), найденный в Индии, – считается некоторыми учеными самым ранним из гоминид. Его возраст оценивают в 9–12 млн. лет.
Представление о рамапитеке как первом из гоминид было сильно поколеблено, когда позднее в Пакистане были обнаружены остатки сходного существа, названного Sivapithecus indicus. Фактически ряд антропологов в настоящее время использует название Sivapithecus для всех остатков Ramapithecus. Изучение остатков сивапитека показало, что он ближе к орангутану, чем к африканским видам человекообразных обезьян. (По существу, это подтверждает данные биомолекулярных исследований, также свидетельствующие о том, что орангутан – более отдаленный родственник человека, чем горилла и шимпанзе, и его отделение в ходе эволюции от группы "человек – шимпанзе – горилла" произошло в период от 10 до 11 млн. лет назад.)
Австралопитек. До недавнего времени самым большим белым пятном в ископаемой летописи эволюции человека оставался плиоцен – эпоха длительностью ок. 6 млн. лет, завершившаяся ок. 2 млн. лет назад. Для изучающих эволюцию человека плиоцен особенно важен, поскольку он предшествует непосредственно эпохе плейстоцена, в котором были впервые обнаружены ясные и неоспоримые ископаемые остатки гоминид, известных как австралопитековые (Australopithecinae).
Эти тонкокостные существа с небольшим мозгом являются самыми ранними из известных настоящих гоминид. Первая из находок австралопитековых, состоящая из части черепа ребенка, была сделана в 1924 в известковом карьере близ Таунса в Бечуаналенде (ныне Ботсвана) южноафриканским анатомом Р.Дартом. Дарт определил находку как новую форму гоминид и назвал ее Australopithecus africanus. Другие формы, названные парантропом (Paranthopus) и плезиантропом (Plesianthropus), были найдены позже и также в Африке. В 1959 в ущелье Олдовай в нынешней Танзании Луис Лики обнаружил одного из самых древних представителей этой группы – зинджантропа (Zinjanthropus).
К настоящему времени найдены остатки или значительные фрагменты скелета более ста особей австралопитековых. Они особенно интересны тем, что раскрывают относительные темпы эволюции различных частей тела гоминид. Австралопитековые имели рост в среднем 122–152 см и были прямоходящими, что следует из формы их длинных костей ног и рук и подтверждается формой таза и характером сочленения черепа с позвоночником. Но объем их черепной коробки был не больше, чем у современных шимпанзе и горилл, – ок. 500 куб. см. По современным представлениям, увеличение размеров черепной коробки и мозга произошло несколько позже начала прямохождения, увеличения подвижности рук, гибкости кисти и развития зубов.
Находки последних десятилетий проливают свет на происхождение австралопитековых. В 1974 в Эфиопии, в районе Афар, Д.Джохансон обнаружил при раскопках примитивный вид этих человекообезьян, первая представительница которого (обнаружены были кости женской особи) получила уменьшительное имя Люси. Научное название этого вида – A.afarensis, его возраст оценивается в 3–4 млн. лет, и большинство специалистов считает, что он был общим предком двух линий, разошедшихся в ходе эволюции: австралопитековых и гоминид. Линия австралопитековых, включавшая такие виды, как A.africanus, A.ethiopicus, A.boisei и A.robustus, угасла немногим более миллиона лет назад. Линия гоминид, к которой принадлежали исчезнувшие виды Homo habilis и H.erectus, привела к современному человеку – H.sapiens. Хотя A.afarensis находится в точке разветвления двух названных линий, он недостаточно похож на человекообразных обезьян, чтобы быть единственным связующим звеном между австралопитековыми и той древней "человекообразной обезьяной", от которой он произошел в ходе эволюции. "Обезьяна-прародитель" в действительности являлась не совсем человекообразной обезьяной, а скорее общим предком, как человекообразных обезьян, так и человека, и жила (что следует из результатов биомолекулярных исследований) 4–6 млн. лет тому назад. Следовательно, должны были существовать одна или несколько форм, которые еще более примитивны, чем A.afarensis.
В декабре 1992 такая примитивная форма была обнаружена в Эфиопии, неподалеку от места находки Люси, т.е. A.afarensis. Исследование этого примитивного вида, названного A.ramidus (в публикации 1994), показало, что его возраст 4,4 млн. лет; он имел по всем признакам значительное сходство с шимпанзе, но обладал и некоторым человекоподобием, например, относительно коротким основанием черепа и клыками той же формы, что и у гоминид. Неожиданным оказалось то, что A.ramidus был лесным жителем. Это удивительно, поскольку считается, что предок человека жил в районах открытой саванны, и именно условия открытой саванны стали важным, если не ключевым, фактором развития в ходе эволюции вертикального положения тела, т.е. хождения на двух ногах. Был ли A.ramidus существом двуногим, остается неизвестным.
Этапы эволюции приматов и человека
Эволюция Homosapiens – это эволюция всего – навсего одного из многих миллионов видов, живших на нашей планете.
Основной материал антропологам поставляют археологические раскопки.
Для разрешения проблемы происхождения и эволюции человека необходимо изучать костные остатки древнейших и древних людей в неразрывной связи с условиями жизни этих людей, со способами добывания средств к существованию и характером орудий производства. Антропология должна таким образом обращаться, таким образом, к результатам исследований геологов, палеонтологов, палеографов и археологов.
История человека складывалась как головоломка – по кусочкам. Сначала (в 17 в) они находили орудия труда древних людей, потом стали обращать внимание на фрагменты черепов и скелетов, лежавших на земле рядом с этими орудиями. Сначала (1865год ) был описан неандерталец, потом питекантропы, и только потом более древние формы. Еще в начале второй половины нашего века принято считать, что человек впервые появился в Азии около 8000 тыс. лет назад и что между обезьяной и человеком есть так называемое " недостающее звено", но последующие открытия в Африке убеждают нас в том, что возраст человека значительно больше, а его прародина - Африка.
На протяжении многих миллионов лет предки человека населяли ограниченную область на востоке Африки. Здесь, на озере Виктория, 18 млн. лет назад жил проконсул – наш общий с человекообразными обезьянами предок; здесь более 4 млн. лет назад возник афарский австралопитек – наш прямоходящий предок. Здесь провел всю свою историю первый представитель рода Человек – умелый человек, появившийся более 2 млн. лет назад, и здесь 1,6 млн. лет назад возник прямостоящий человек. Где начал свой путь наш вид – разумный человек – не известно, но где-то поблизости.
Важно и то, что африканские находки и вместе с достижениями генетики опровергают существование ранее представления о постепенном изменении организма человека вследствие трудовой деятельности. Новые открытия говорят о том, что прямохождение, увеличение размеров мозга и другие "человеческие" признаки появились за несколько миллионов лет до возникновения трудовой деятельности и о том, что человек появился не в результате постепенного поступательного развития, а результате некого скачка, при этом он длительно время существовал вместе со своими предками (австралопитековыми), которые потом вымерли.
В настоящее время выделяют следующие основные этапы в эволюции человека:
Дриопетек (рамапитек)
Австралопитек
Человек умелый
Человек прямоходящий
Неандертальский человек (палеантроп)
Неоантроп (это уже человек современного типа, Homosapiens).
Дриопитеки (Dryopithecinae )
Вымершие Dryopithecinae рассматриваются некоторыми учеными как одно из трех подсемейств, образующих семейство Hominidae (в два других подсемейства входят крупные человекообразные обезьяны, человек и вымершие "почти люди").
Косные останки дриопитеков найдены в Западной Европе, Южной Азии и Восточной Африке в слоях древностью 12-40 млн. лет. Дриопитеки рассматриваются как общая исходная предковая группа для современных человекообразных обезьян и человека. Группы этих обезьян лазали по дубам и субтропическим деревьям и раскачивались на их ветвях. Они, очевидно, питались плодами, поскольку их коренные зубы, покрытые тонким слоем эмали, не были приспособлены для пережевывания грубой пищи.
Впервые нижняя челюсть дриопитека была найдена в 1856 г. во Франции и описана Эдуардом Ларте. Наиболее древние остатки дриопитеков найдены в слоях нижнего миоцена возле оз.Виктория на острове Русинга в Восточной Африке (проконсул африканский – человекообразная обезьяна, жившая 18 млн. лет назад). Находки костных остатков проконсула, в том числе черепа, показывают промежуточное положение этой обезьяны между примитивными мартышкообразными и более поздними человекообразными. К среднемиоценовым слоям относятся многочисленные находки дриопитеков в центральной Европе, европейской части СССР, на Кавказе, Малой Азии, на севере Индостана и в Южном Китае. Известны и более поздние, но малочисленные находки дриопитеков из нижнего плиоцена.
Рамапитеки
В качестве предков человека некоторые указывают на рамапитека. Его рассматривали как самостоятельный род высокоразвитых гоминоидов.
Рамапитек - представитель ископаемых человекообразных обезьян, обитавших около от14 до 10 млн. лет назад на территории Юго-Восточной Африки, Передней и Южной Азии. Рамапитеки - некрупные обезьяны (ростом около метра и весом 18-20 кг) - вели преимущественно наземный образ жизни. Считается, что рамапитеки были той ветвью развития, которая привела к появлению узконосых обезьян (орангутангов).
Первую обезьяну такого типа нашел Дж. Льюис в 1932 г. В Северной Индии. Она описана в 1934 г. Под названием "короткомордый рамапитек". Предположительный геологический возраст – 10-12 млн. лет. Обнаруженные челюсти, два фрагмента верхней и нижней челюсти от двух разных особей рамапитеков были извлечены в Сиваликских холмах, отличались от челюстей классических дриоптеков укороченностью зубной дуги, имевшей, как полагали параболическую форму, округленную спереди, с уменьшенными клыками и резцами. Клыки не выступали из зубных рядов, а первый нижний предкоренной зуб не был вытянутым, как у понгид, а двубугорковым как у человека. Важно отметить, что в верхней челюсти между ячейкой клыка и первым премоляром отсутствует диастема для вхождения нижнего клыка при смыкании челюстей, а это свидетельствует скорее в пользу включения рамапитека в человеческую родословную, чем против его признания предком человека. Судя по строению длинных костей и позвонков, они могли иногда выпрямляться и передвигаться некоторое время на двух задних конечностях. Благодаря большой площади жевательной поверхности зубов рамапитеки были лучше приспособлены к питанию относительно жестким растительным кормом, в котором преобладали семена злаков, корни и побеги. Сбор семян трав требовал большой точности движения пальцев рук. Возможно, что, как и современные шимпанзе, рамапитеки от случая к случаю использовали камни и палки, защищаясь от хищников или добывая пищу.
Австралопитек - непосредственный предок человека
В 1924 г. Р.Дарт, профессор Иоганнесбургского университета, обнаружил в Южной Африке череп ископаемого гоминида, который по строению костей лицевого отдела черепа и зубов принадлежал существу, занимавшему среднее положение между обезьяной и человеком. Р.Дарт назвал его австралопитеком, или южной обезьяной (от лат. australis - "южный", pithecus - "обезьяна"). Возраст южноафриканского австралопитека составляет около 3,0—2,3 млн лет.
- Австралопитек (africanus), 1924 г. Южная Африка
- Австралопитек (afarensis) - "Люси", 1978 г. Восточная Африка. Возраст 3,9-3,4 млн лет
- Австралопитек (ramidis), 1994 г. Эфиопия. Возраст 4,4 млн лет
- Австралопитек (robustus), 1938 г. Южная Африка. Возраст 1,9—1,6 млн лет
- Австралопитек (boisei), 1959 г. Танзания, первое название зинджантроп
На основании изучения тазовых и бедренных костей австралопитеков считается, что они были двуногими существами, способными стоять и передвигаться подобно человеку. Жили они в пещерах, которые использовали в качестве убежищ, и изготавливали простейшие орудия. Австралопитеки были охотниками на мелких животных, но также использовали в пищу зерна, плоды диких деревьев, корни растений.
Австралопитек (Australopithecinae)
Австралопитеки - род высших двуногих человекообразных приматов, обитавших преимущественно в Восточной и Южной Африке от 4 до 1 млн. лет назад. Австралопитеки имели небольшое тело (длина в среднем 120-130 см), объем мозга колебался от 300 до 570 см3. По-видимому, среди древнейших австралопитеков были предки как поздних, более массивных австралопитеков, так и рода "человек".
Наша цивилизация начинается с австралопитеков. Встав на две ноги, они открыли новый способ выживания. Учёные спорят: "какое преимущество дало обезьяне хождение на двух ногах? Некоторые учёные считают, что австралопитеки изготовляли примитивные орудия, и для этого им и потребовалось прямохождение. Для изготовления орудий труда вовсе не нужно ходить на двух ногах. Четырьмя руками их сделать даже легче и значительно быстрее. Другие учёные считают, что хождение на двух ногах позволяло австралопитеку дальше видеть и как можно раньше обнаружить хищника. Это довольно сомнительное преимущество. Если обезьяна видела хищника, то и он её видел. Даже современный человек не может убежать ни от одного хищника, а он бегает гораздо быстрее австралопитека. Тот только недавно стал ходить на двух ногах и ходил на них довольно неуклюже, а бегал даже медленнее своего предка. Любой хищник мог легко его догнать. Хождение на двух ногах имеет и ряд других серьёзных недостатков.
Последние десятилетия Африканский континент дал значительное количество костных остатков плиоценовых гоминид, каковыми являются Австралопитеки. Самая яркая и отличительная черта австралопитеков – это прямохождение. От афарского австралоитека обрзовался целый кустик других видов австралипитеков. Последние из них вымерли лишь около 1 млн лет назад.
В ноябре 1974 г., в Хадаре Эфиопии (впадина Афар - Африка) был найден палеонтологом К. Джохансон, скелет молодой женской особи, Al-228- 1(гоминид), возрастом 3,5 - 4 млн лет, названного афарским австралопитеком или между собой палеонтологи назвали "Люси". Пока это самая полная находка австралопитека такой древности, сохранилось примерно 40 % всех Косте , среди них 52 крупные части и многочисленные мелкие фрагменты. Впервые оказалось возможным с достаточной степенью точности охарактеризовать некоторые черты внешнего облика австралопитеков.
Люси была ростом всего около метра. Точная реконструкция длины тела на основе расчетных формул для современных человеческих популяций осложняется из-за этих различий в пропорциях. Видимо, в наибольшей степени приближается к действительному определению роста Люси по длине ее бедра: всего 108 см. На черепе виден гребень для прикрепления мощной жевательной мускулатуры. Зубы у Люси – зубы всеядной обезьяны, причем клыки длиннее остальных зубов, а это значит, что движение нижней челюсти из стороны в сторону у нее ограничены (такие движения нужны для перетирания грубой растительной пищи). У всех потомков Люси, как рода австралопитек, так и рода человек, клыки укороченные, это ясно свидетельствует о том,что все эти виды прошли период приспособления к грубой растительной пищи. Люси могла защищаясь, не просто больно куснуть, но нанести глубокие рваные раны, а при необходимости накрепко вцепиться зубами. Словом она была вооружена не хуже, чем современные человекообразные обезьяны, для которых в бою зубы важнее рук.
Объем мозга у Люси и ее потомков был не больше, чем у современных человекообразных обезьян того же роста. Для того, чтобы скелет четвероногой обезьяны преобразовался в скелет Люси, эволюции требуется 5-7 млн лет. Значит, гоминиды начали свой путь по Земле на двух ногах 8-10 млн лет назад. А первые каменные орудия имеют возраст 2,5 млн лет. С идеей прямохождения ради изготовления орудий труда пришлось расстаться.
Когда изучили скелет Люси, выяснилось, что она ходила на двух ногах не кое-как, а полностью выпрямившись. "Конструкция" Люси не пригодна для хождения на четвереньках и лазанья по деревьям. "Она не просто умела ходить на двух ногах, но это был для нее единственный способ передвижения", - такой вывод сделал анатом К.О. Лавджой после тщательного изучения всего ископаемого материала. Более того, таз Люси, если сравнивать его с тазом современной женщины, был лучше приспособлен к прямохождению, потому что был уже. А чем таз уже, тем легче бегать. Но при узком тазе можно рожать детей только с небольшой головой!
Итак, человек не сам стал двуногим. Он произошел от двуногого животного.
Австралопитек африканский
Приспосабливаясь к питанию грубой растительной пищей, часть потомков Люси 2,8 – 3 млн лет назад дала начало новому виду более мощному, с укороченными клыками, - африканскому австралоптеку ("африканская южная обезьяна"). Этот вид прекратил свое существование около миллиона лет назад до наших дней. Большинство ископаемых находок обнаружены в пещере Стеркфонтейн в Южной Африке. Другие находки говорят о том, что африканус обитал также в Эфиопии, Кении и Танзании. По сравнительному соотношению размеров мозга и тела африканус занимает промежуточное положение между современными и человекообразными обезьянами и людьми.
Маленький, хрупко сложенный африканус, похожий на человекообразную обезьяну, а весил не больше 12-летней европейской девочки. Он ходил, выпрямившись, хотя мускулы его ног и отличались от наших. Руки были сравнительно длинные, а их большие и прочие пальцы, вероятно, менее умело обращались с предметами, чем это получается у нас. Нижняя часть его лица выступает вперед, но не столь значительно, как у человекообразных обезьян. На некоторых черепах заметны следы затылочного гребня, к которому были прикреплены сильные мышцы шеи. Мозг был не больше, чем у гориллы, но слепки свидетельствуют о том, что по строению мозг несколько отличался от мозга человекообразных обезьян.
Ученые расходятся во мнении относительно того, какой образ жизни вел африканус. Строение зубов и челюстей позволяет думать, что этот обезьяночеловек пережевывал растительную пищу, но, возможно, также обгладывал мясо животных, убитых хищниками. Специалисты оспаривают его способность к изготовлению орудий: большинство костяных "орудий", обнаруженных около ископаемых остатков африкануса, оказались просто остатками пищи гиен или других хищников.
Как считают некоторые специалисты, самой древней находкой африкануса является обломок челюсти, датируемый 5,5 миллиона лет, из Лотегама в Кении, а самый молодой экземпляр имеет возраст 700 тысяч лет. Но нельзя с полной уверенностью утверждать, что эти австралопитеки действительно относятся к виду африканус.
От него 2,3 лет назад произошли еще два вида, а от одного из них около 1,8 млн лет назад – еще один вид.
Австралопитеки массивные
Австралопитек могучий
Австралопитек могучий, которого в одно время называли парантропусом ("около – человек"), был более крупным и лучше развитым физически, нежели австралопитек африканский. По всей видимости, австралопитек могучий появился около 2,5 миллиона лет назад. Все останки представителей этого вида найдены в Южной Африке, в пещерах, куда их, вероятно, затащили хищные звери. Вымер этот вид около 1,5 миллиона лет назад. Австралопитек могучий был такого же роста, как многие европейские женщины - 1,6 м, но весил меньше, чем большинство из них, - около 50 кг. По сравнению с африканусом у него был более крупный и плоский череп, вмещавший более крупный мозг - около 550 см3, и более широкое лицо. К высокому черепному гребню были прикреплены мощные мышцы, приводившие в движение массивные челюсти. Передние зубы были такими же, как у африкануса, а коренные - более крупными. При этом коренные зубы у большинства известных нам экземпляров обычно сильно изношены, несмотря на то, что они были покрыты толстым слоем прочной эмали. Это может свидетельствовать о том, что животные питались твердой, жесткой пищей, в частности зернами злаков.
От него, возможно, произошел австралопитек Бойса. А строение черепа австралопитека могучего позволяет предположить, что он был также предком гориллы.
Австралопитек Бойса
Этот наиболее крупный и сильный из всех австралопитеков получил свое название в честь английского бизнесмена Чарльза Бойса, помогавшего финансировать палеонтологические исследования в Восточной Африке, в результате которых в 1959 г. был найден неизвестный ранее науке вид "южных обезьян". Первоначально Австралопитек Бойса именовался Zinjanthropus, что означает "восточно-африканский человек". Время его существования - от 2,5 до 1 миллиона лет назад.
Некоторые палеоантропологи считают, что австралопитек Бойса не является самостоятельным видом, а представляет собой лишь местную разновидность австралопитека могучего. Тем не менее, для него характерны многие отличительные признаки. Судя по реконструкциям, сделанным на основании изучения ископаемых костей черепа и конечностей, некоторые экземпляры этого вида достигали роста современного человека. Мозг у них был таких же размеров, как у австралопитека могучего, т. е. примерно втрое меньше нашего мозга. Эти существа ходили прямо. Своим мощным телосложением они напоминали гориллу. Как и у горилл, самцы были, по-видимому, значительно крупнее самок. Подобно горилле, австралопитек Бойса имел большой череп с надглазничными валиками и центральным костным гребнем, служившим для прикрепления мощных челюстных мышц. Но по сравнению с гориллой, гребень у австралопитека Бойса был меньше и более выдвинут вперед, лицо - более плоское, а клыки - менее развитые. Из-за огромных коренных и предкоренных зубов это животное получило прозвище "щелкунчик". Но биомеханические исследования показывают, что эти зубы не могли оказывать сильного давления на пищу, и были приспособлены для пережевывания не очень жесткого материала, например листьев.
Поскольку вместе с костями австралопитека Бойса, возраст которых составляет 1,8 миллиона лет, была найдена оббитая галька, можно предположить, что эти существа могли использовать камень в практических целях. Однако не исключено, что представители данного вида попросту пали жертвой своего современника - человека умелого, преуспевшего в применении каменных орудий.
Образ жизни австралопитеков
Все эти австралопитеки "решали" свои эволюционные задачи "силовым методом": зубы у них были крупные, челюсти мощные, могучая жевательная мускулатура прикреплялась к крепкому гребню на черепе. Сами они были очень большими, так что хищникам с ними было справиться не просто. Каменных орудий они не употребляли. Мозг за 2 млн лет не увеличился. Словом жили по принципу "сила есть - ума не надо". Они обитали в Африке рядом предками человека, но нашими предками они не являлись. Это наши двоюродные родственники, вымершие около миллиона лет назад.
Переход от жизни в лесу к жизни в саваннах повлек за собой изменение характера питания. Первоначальное употребление исключительно растительной пищи сменилось всеядностью, причем мясо приобретало всевозрастающее значение. В условиях савнн значительно возросшие источники получения животного белка наряду с нерегулярными потреблением фруктов должны были всячески содействовать характерной для ранних гомонид склонности к охоте. Другая вероятная причина особого значения животной пищи могла заключатся в необходимости пополнения сокращающихся растительных ресурсов природной среды, которая становилась все более и более засушливой.
Базовая стоянка является одной из наиболее важных, характерных черт образа жизни ранних гомонид, поскольку она представляет собой место длительного, хотя и временного обитания. Существование базовых стоянок, вероятно, может быть объяснено продлением периода несамостоятельности молодых членов коллектива и большей их зависимостью от взрослых, что придает особое значение отношениям матери и ребенка. Исходя из сроков прорезывания зубов у австралопитековых, эта зависимость длилась примерно столько же, сколько и у современного человека. Исследования приматов показывают, что подобное удлинение периода, предшествующего достижению полной зрелости, тесно связано с усвоением навыков и образа поведения, необходимых для самостоятельной жизни. Молодой шимпанзе достигает самостоятельности между семью и восемью годами, и передача более сложных навыков, которыми владели ранние гоминиды, должна была потребовать еще более длительного времени. Возникновение базовых стоянок, таким образом, может рассматриваться как следствие ограничения подвижности коллектива, обусловленное длительным периодом обучения.
Все эти древние стоянки находились у воды, что, возможно, отражает неспособность человека жить вдали от постоянных водных источников и человек, и животные, бесспорно, отдавали предпочтение именно таким районам. Вероятно, не существовало и соответствующих средств для хранения и транспортировки воды, а стоянки, расположенные у воды, обеспечивали не только наилучшие возможности для охлты, но и постоянное снабжение водой для утоления жажды, которая непременно возникала в результате употребления в пищу сырого мяса. Возможно, именно этими причинами руководствовались при выборе места для стоянки, что в то же время могло быть связано густым растительным покровом, типичным для расположенных близ воды участков в тропиках. Это не только давало изобилие растительной пищи, но и позволяло укрываться на деревьях в целях защиты от крупных хищников. Очевидно также, что использование палок и камней должно было служить одним из средств самозащиты. С другой стороны, уменьшение размеров клыков – одна из характерных особенностей расположения зубов у австролопитеков – совпало с более широким и более эффективным применением орудий, явилось его прямым результатом.
В настоящее время не существует надежных методов, с помощью которых можно было бы определить численность коллектива гоминид, однако размеры участка, занимаемого базовой стоянкой, дают основания полагать, что эти коллективы невелики. Вместе с тем вполне вероятно, что действительная их численность фактически непрерывно менялась. Было подсчитано, что ископаемые останки австралопитековых из пещеры Сварткранс принадлежат 50-70 особям. Стаи, по всей видимости, состояли из членов двух-трех сходных по составу семей, включавших матерей с детенышами и, возможно, трех-четырех взрослых самцов. Все члены стаи находились во взаимозависимости и делили между собой добычу, полученную охотой и собирательством. В самом деле, раздел пищи является основной характеристикой человеческого общества и одним из важнейших различий в образе жизни человека и млекопитающих. По мере увеличения наших знаний об образе жизни африканских человекообразных обезьян становится все более очевидным, что австралопитековые не были агрессивными "вооруженными убийцами". В основе агрессивности современного человека и многих животных лежит необходимость защиты своей территории, но для африканских человекообразных обезьян энергичная деятельность подобного рода, судя по наблюдениям, нехарактерна.
4. Факторы и критерии гоминизации
Гоминизация – это процесс очеловечивания обезьяны, начало которому положило формирование первых специфических человеческих особенностей, а завершением было возникновение человека современного типа, человека разумного. Соматология (от греч. soma, родительный падеж somatos — тело и логия), отрасль морфологии человека, Соматология (от греч. soma, родительный падеж somatos — тело и логия), отрасль морфологии человека, Соматология (от греч. soma, родительный падеж somatos — тело и логия), отрасль морфологии человека, Это комплексный феномен, включающий развитие прямохождения, увеличение головного мозга, адаптацию (приспособление) к трудовой деятельности руки, изменение зубочелюстного аппарата (строение зубной системы), формирование членораздельной речи, мышления, сознания, интеллекта, перестройки онтогенеза, развитие социальной организации, материальной культуры и т.д. Все эти морфологические признаки в той или иной степени отражают и изменения в поведении. То есть трудовая деятельность, явившись решающим фактором гоменизации, входит в морфологический критерий гоминид опосредованно, через "отпечаток" её в анатомических структурах.
Факторы, способствующие гоминизации приматов:
1. Развитие прямохождения
2. Увеличение головного мозга
3. Адаптация руки к трудовой деятельности
4. Изменение зубочелюстного аппарата
5. Формирование членораздельной речи, мышления, сознания
6. Разаитие социальной организации
Самые древние достоверные представители homo известны примерно с 2,0-2,5 млн лет назад. В результате кладогенеза (расщепления) возникло 2-3 линии гоминид.
Человек умелый (Homohabilis)
Homohabilis(человек умелый), по общему мнению, представлял собой первый известный вид нашего рода Homo. Homohabilis представлял собой двуногое существо ростом 120—140см. У этого вида сравнительно более крупный мозг, чем у австралопитека; таким образом, мозговая коробка была больше, а лицо меньше и менее выдвинуто вперед. Верхняя и нижняя челюсти были меньше, чем у австралопитека бойсеи (зинджа), но почти не отличались от челюстей питекантропа и современного человека, зубной ряд имел открытую форму, похожую на латинскую букву U. Руки были короче по сравнению с ногами. Кисть руки человека "умелого" была способна к силовому захвату большой мощности; об этом свидетельствовали широкие ногтевые фаланги и массивные трубчатые кости кисти. Форма тазовых костей позволяла как ходить на двух ногах, так и рожать детей с крупными головами. Морфологически Homohabilis тесно примыкает к австропитековым.
Первоначально Homohabilis назвали "презинджантроп" (то есть предшественник зинджантропа), поскольку останки его обнаружили в немного более глубоком горизонте. Однако в дальнейшем выяснилось, что презинджантроп был современником зинджантропа.
На месте нынешнего Олдувайского ущелья, у озера (или водного потока) существовавших около 2 млн лет назад, в самом низу — слой (I) толщиной до 40 м, были обнаружены останки австралопитековых — зинджантропа и человека "умелого" и вместе с ним самые древние орудия человека, названные олдувайскими. Возраст этих находок по калий-аргону — 1,75-1,85 млн лет. Примитивное "праорудие" получило название "чоппер", то есть "ударник", а сама эта древнейшая каменная культура стала именоваться галечной, или олдувайской, — по имени ущелья.
Заслужить высокое звание Homo habilis (человек умелый) - значило овладеть рядом совершенно необходимых умений, приобрести сумму знаний. Во мраке пещер одним из первых понадобилось умение сохранять и добывать огонь. Постоянные очаги палеолитических убежищ использовались для обработки продуктов питания и создания комфортной среды обитания. Но одновременно они были своеобразным социокультурным центром, символом бытового, производственного и идеологического единства древних микроколлективов. Вероятно, отсюда берут начало многие культы и обряды, в том числе и неугасимый огонь на захоронениях павших в сражениях XX века...
До последнего времени археологи считали, что огонь начал служить человеку примерно 0,5 млн. лет назад, когда Homo erectus или ранний Homo sapiens поселились в холодных областях Европы и Азии. Обожженные кости возрастом 0,7- 1,0 млн. лет считались следствием действия естественного огня (молния, лесной пожар). Но в 1988 г. в Сварткапской пещере (Южная Африка) обнаружено 270 костей антилопы, зебры, кабана, одного из видов гоминид. Их возраст - 1,0- 1,8 млн. лет. Находки костей в разных слоях указывают на многократное разведение огня именно умелым человеком.
Использование огня сразу выделило человека среди других живых существ на Земле. Оно дало ему новые орудия труда и охоты, позволило использовать жареную, а затем и вареную пищу, что привело к настоящей революции в его развитии, оздоровило условия пребывания в пещерных жилищах (дым отгонял комаров, гнус, мошку).
Ранний человек был медлительнее и слабее, чем крупные хищники, и не имел такого естественного оружия, как клыки и когти. И все-таки ранние гоминиды научились компенсировать эти недостатки. Они стали придавать кускам камня, костей и дерева форму, позволяющую резать, скрести и копать. В отличие от клыков и когтей, такие орудия можно было собирать, хранить или по своему желанию менять одно на другое. Со временем эти орудия дали человеку беспрецедентную власть над окружающей средой.
Первыми орудиями, вероятно, были осколки костей, острые палки и подносы из коры для сбора пищи. Такие, в основном непрочные, орудия не сохранились. Но камень оказался долговечнее. Мы знаем, что ранние гоминиды в Эфиопии сознательно расщепляли небольшие камни, возможно, для того чтобы получить твердые, острые края для резки мяса. Господствовали три основных вида орудий: камни-многогранники для нанесения удара, чопперы и ножи из отщепов. Их производство не содержит никакого секрета; это орудия малых размеров, для выделки которых не существовало каких-либо установленных норм. Однако рубящие края орудий из отщепов явно свидетельствуют о наличии зачаточных знаний по обработке камня. Подобные коллекции орудий могут относиться к самому началу эпохи обработки камня.
Таким образом, орудия олдувайской культуры представляют собой основной инвентарь, необходимый для того, чтобы обеспечить снабжение разнообразной растительной и животной пищей, которая доставлялась на базовую стоянку. Очевидные свидетельства подобного образа жизни дают нам костные останки животных, обнаруживаемые на стоянках. Отдельные животные вместе с тем могли служить добычей лишь при наличии навыков коллективной охоты, и новейшие исследования охотничьих привычек шимпанзе показали, что к настоящему времени навык коллективной охоты стал у них неотъемлемой частью образа жизни.
На тесную связь между эволюцией мозга и развитием технических навыков было обращено особое внимание. Более эффективное приспособление к охотничье-собирательскому образу жизни явилось следствием развития технических навыков, что, в свою очередь, оказалось возможным в результате увеличения частей мозга, связанных с умением передвигаться и способностью к общению. Таким образом, культурная эволюция и увеличение размеров мозга находятся по отношению друг к другу в прямой зависимости. Сравнительная быстрота, с которой австралопитек африканский превратился в Homohabilis (в то время как австралопитек массивный оставался биологически законсервированным, пока не вымер в результате борьбы за существование), оказалась возможной лишь благодаря умелому изготовлению и использованию орудий.
Человек прямоходящий (Homoerectus) (архантроп)
Homo erectus (люди прямоходящие) жили в период с 2 миллионов до 400 тысяч лет назад. По одной версии, они появились в Африке, но постепенно расселились по всему Старому Свету.
Первые окаменелые останки Homo erectus были найдены Эженом Дюбуа в 1891 г на Яве, который ввел для их обозначения родовое название Pithecantropus.
С тех пор было найдено много других останков, но тем не менее они остались фрагментарными, что не позволило антропологам определиться, какие физиологические вариации нормальны для этого вида. Большинство находок по-своему уникально, но достаточно сложно судить, был ли вид Homo erectus разнообразным по внешнему виду, или же существовало несколько видов. В настоящее время почти любая подобная находка автоматически причисляется к Homo erectus, что расстраивает некоторых специалистов.
Homo erectus (человек прямоходящий) имел более крупные мозг и тело, чем его вероятный предок Homo habilis, и во многих отношениях уже был похож на своего крепко сложенного непосредственного потомка - современного человека.
Его череп, хотя и имел самые толстые стенки по сравнению с черепами всех прочих представителей человеческой трибы, сохранил архаические черты. Череп Homo erectus был длинным и низко посаженным, с костистой выпуклостью сзади, со скошенным лбом, толстыми надглазничными валиками, c более плоской, чем у нас, лицевой частью, с крупными выдвинутыми вперед челюстями, более массивными, чем у нас, зубами (но все-таки немного меньшими, чем у Homo habilis); подбородок отсутствовал.
Сильные мышцы сзади шеи были присоединены к заднему черепному бугру и поддерживали голову с тяжелой лицевой частью, не давая ей провисать вперед. Объем мозга составлял в среднем 880-1100 см3 (мнения специалистов расходятся), что больше, чем у человека умелого, хотя и меньше, чем у современного человека.
Некоторые взрослые особи, вероятно, достигали роста 1,8 м и весили по крайней мере столько же, сколько и мы. Homo erectus жил от 1,6 миллиона до 200 тысяч лет назад, а возможно, и в течение более длительного периода. Появившись впервые, вероятно, в Африке, отдельные группы распространились потом в Европу, Восточную Азию (сюда относится пекинский человек, или Sinanthropus) и Юго-Восточную Азию (яванский человек, или Pithecanthropus). По всей видимости, темпы эволюции отдельных изолированных популяций были различны.
Более высокоразвитый интеллект и более совершенная техника изготовления орудий помогли этому охотнику раннего каменного века колонизовать новые места обитания - заселить небольшими группами Африку, Европу и Азию (главным образом Южную), а также продвинули человека прямоходящего далеко вперед по сравнению с предшествовавшими ему гоминидами, предоставив этому виду возможность существовать в новых природных и климатических условиях.
В Европе 400 тысяч лет назад у отдельных особей появились черты, встречающиеся у ранних представителей нашего вида Homo sapiens. 200 тысяч лет назад Homo erectus, вероятно, уже вымер; возможно, он стал жертвой конкуренции со стороны своих собственных потомков.
У Homoerectusобнаруживаются как "примитивные", так и более "прогрессивные" признаки и можно рассматривать его морфологический статус как промежуточный между Australopithecusи Homosapiens.
Основные примитивные признаки Н. erectusтаковы:
1. Кости черепа очень толстые.
2. Сильно развитый надглазничный валик, не прерывающийся на всем протяжении.
3. Очень покатый лоб.
4. Мощный затылочный гребень. <...>
10. Массивная нижняя челюсть. .
11. Отсутствие выдающегося вперед подбородка.
12. Образование нескольких подбородочных отверстий.
13. Крупные верхние резцы.
Прогрессивные признаки Н. erectusследующие:
1. Объем мозговой коробки превосходит минимальные величины для Н. sapiens; выпуклый черепной свод.
2. Относительные размеры лицевого черепа меньше, чем у австралопитека.
3. Зубная дуга параболической формы.
4. Морфология зубов ближе к Homosapiens, чем к австралопитеку.
5. Начальная стадия развития подбородка (у пекинского человека).
6. Кости конечностей по размерам и пропорциям аналогичны костям Homosapiens.
Антропологи не могут точно сказать, где именно произошел переход от австралопитека к Н. erectus. Принято считать, что предки гоминид обитали в Африке, и что именно на этом континенте происходила начальная стадия их эволюции. В пользу этого говорят находки ряда последовательных форм австралопитеков на Африканском материке и близкое родство человека и современных человекообразных обезьян, обитающих в Африке. С появлением Н. erectusгоминиды значительно расширили область своего обитания в Старом Свете, постепенно заселив более удаленные от экватора области и страны умеренного пояса.
Образ жизни
Скопления костей и каменных орудий находят на тех местах, где семейные группы или более крупные объединения H.erectusразбивали свои стоянки. Большинство из стоянок служило пристанищем лишь в течение нескольких дней, пока люди планировали охоту, свежевали туши убитых животных и делили между собой их мясо, собирали съедобные растения. Но человек прямоходящий нередко сооружал и укрытия, которые известны по находкам в прохладных северных районах. Самые крупные хижины этой сезонной стоянки на морском берегу могли вмещать до 20 человек.
Огонь был, по-видимому, знаком людям еще до появления Homoerectus. Но можно утверждать, что именно Homoerectusпервым начал систематически использовать огонь для обогрева, приготовления пиши, защиты от хищников и для охоты на диких животных.
Для человечества все эти достижения означали важные изменения — культурное развитие теперь приобретало большее значение, чем биологическая эволюция.
На ряде стоянок человека прямоходящего найдены серьезные доказательства того, что эти предприимчивые гоминиды не просто собирали растения и срезали мясо с туш убитых хищниками животных, но и сами активно охотились на крупную дичь, объединяясь для этого в группы с тем, чтобы спланировать и осуществить совместное преследование или засаду. Находки, обнаруженные на трех континентах, дают представление о приемах охоты и о животных, которые были ее объектом.
Самые первые ручные рубила появились примерно в то же время, что и Homoerectus. Поскольку для изготовления подобных однотипных орудий требовалась немалая сообразительность, можно заключить, что их вероятным изобретателем был именно этот высокоразвитый гоминид. Но все данные указывают на то, что эволюция предметов материальной культуры характеризовалась чрезвычайно замедленными темпами.
Предметы материальной культуры, дают основания предполагать, что если человек среднего плейстоцена в действительности обладал способностью говорить, то язык этот, несомненно, отличался меньшей выразительностью по сравнению с языком, на котором разговариваем мы"
Человек разумный (Homosapiens)
Из всех видов гоминид, когда-либо существовавших на Земле, Homo sapiens — единственный, чью эволюционную судьбу можно рассматривать в перспективе, поскольку только этот вид дожил до настоящего времени, широко расселившись по земному шару. 300 тысяч лет назад Homoerectusуже начал превращаться в человека вполне современного типа. На протяжении развития рода Homo sapiens более высокоорганизованные виды гоминид, расы и народы, находившиеся на так называемой оси эволюционной магистрали, вытесняли тех, чья организация была ниже или даже упрощалась в процессе адаптации к специфическим средам обитания, таким, например, как горы или тропические леса.
Homosapiens — вид Homo; его характерные признаки: большой объем мозговой коробки — в среднем больше 1000 см3
; надглазничные гребни развиты в различной степени; для лицевого скелета характерен ортогнатизм или слабый прогнатизм. Клыки относительно малы и не перекрываются уже после начальной стадии стирания; скелет конечностей приспособлен к прямохождению.
При рассмотрении ископаемых останков Homosapiensудобно выделить, с одной стороны, формы, близкие к современному человеку, и формы, которые отличаются от современного человека в большей степени, чем различаются представители различных рас живущих ныне людей. Предшественником современного человека в Западной Европе был неандерталец, который один населял эту область во время первого вюрмского оледенения (70—40 тыс. лет назад).
Основные гипотезы эволюции человека в период становления Homosapiens
В течение второй половины XIX в. большинство сторонников эволюционной теории не сомневалось в том, что предками современного вида были европейские неандертальцы. Эта точка зрения отстаивалась многими антропологами и позднее. Важнейшими аргументами в пользу теории "неандертальской фазы" служили следующие факты. Во-первых, все сколько-нибудь достоверно датированные останки древних людей располагаются в определенной последовательности: неандертальцы всегда залегают в слоях более ранних, чем кости неоантропов. Во-вторых, останки неандертальцев, как правило, были обнаружены вместе с мустьерскими орудиями, а люди современного строения связаны с орудиями позднего палеолита. В-третьих, скелеты неандертальцев, главным образом, их черепа гораздо более примитивны по своему строению, чем современные, и, отвлекаясь от некоторых особенностей, в общем могли занять место промежуточного звена между человеком и его гипотетическим животным предком. В-четвертых, некоторые ранние формы людей позднего палеолита обладают отдельными чертами, которые сближают их с неандертальцами и тем самым свидетельствуют об их прямой родственной связи. В-пятых, среди европейских неандертальцев встречаются индивиды с более прогрессивными чертами.
Темпы эволюции были разными в различных местах, что зависело от таких факторов, как мутации, все более широкое использование орудий для тяжелых работ, выполнявшихся раньше при помощи зубов или мышц, миграция и изоляция некоторых популяций. Даже эволюция различных частей человеческого тела проходила неравномерно, что приводило, например, к появлению черепов, сочетавших в себе характерные современные и архаичные черты. В настоящее время большинство специалистов согласны с тем, что все формы Homosapiensпроизошли от Homoerectus. Основные изменения касались размеров мозга, а значит, и размеров черепа.
Неандертальский человек (палеоантроп)
Homosapiensneanderthalensisполучил свое имя по ископаемым остаткам, найденным в долине Неандерталь близ Дюссельдорфа, Германия. Так называемый классический неандерталец из Европы имел крупную удлиненную голову; мозг его был больше нашего, а стенки черепа толще, чем у нас, но тоньше, чем у Homoerectus. На человека прямоходящего неандерталец был еще несколько похож своими мощными надглазничными валиками и покатым лбом. У неандертальца был четко выраженный, похожий на шишку затылочный бугор с большим основанием, к которому прикреплялись шейные мышцы. Широкая лицевая часть сильно выдвинута вперед и скошена назад по бокам, что придавало скуловым костям "обтекаемую" форму. Классические неандертальцы были невысоки, чрезвычайно мускулисты и коренасты, с большими суставами ног и рук. Пропорциями тела они похожи на эскимосов, чье плотное сложение помогает им сохранять тепло в условиях холодного климата. Неандертальский подвид произошел от архаичной формы Homo sapiens, возможно, еще 200 тысяч лет назад. Физические данные и передовые технические приемы неандертальцев сделали некоторых из этих людей среднего палеолита, возможно, первыми гоминидами, способными выносить тяготы зимы в условиях холодного климата. Кроме того, ритуальное поведение неандертальцев свидетельствует о достижении ими более высокого уровня развития человеческих эмоций и самосознания.
Несмотря на все это, около 30 тысяч лет назад эта группа, очевидно, вымерла. Некоторые ученые предполагают, что неандертальцы были уничтожены новым, вполне современным типом человека, который сформировался где-то в другом месте. Иные объяснения состоят в том, что неандертальцы скрестились с нашим подвидом или сами эволюционировали, превратившись в него. По некоторым из них видно, что даже особи, проживавшие совместно в одной группе, значительно отличались друг от друга по строению черепа и челюстей.
Возраст находок колеблется, возможно, от 250 до 30 тысяч лет, но наиболее "зрелые" неандертальцы относятся к первой половине последнего (вюрмского) оледенения: 70—30 тысяч лет назад.
Неандертальцы обитали также в Юго-Западной Азии и, возможно, в Африке, но у некоторых из них отсутствовали те грубоватые черты, которые были характерны для классической европейской формы. Вероятно, у европейских неандертальцев такие черты возникли в результате приспособления к сильным холодам ледникового периода. Некоторые из азиатских и африканских неандертальцев имели более прямые и тонкие конечности, менее мощные надглазничные валики и укороченные, менее массивные черепа. Наряду с надглазничными валиками и выдвинутым вперед лицевым отделом некоторые черепа имели высокий лоб и высокую округлую черепную коробку. За пределами Европы и Юго-Западной Азии не было найдено типично неандертальских остатков. Примерно 40 тысяч лет назад последние неандертальцы Юго-Западной Азии, очевидно, существовали одновременно с людьми, имевшими вполне современный облик.
Орудия неандертальцев называются мустьерскими по находкам в пещере Ле-Мустье во Франции. Они явились шагом вперед по сравнению с более ранними культурами рубил и чопперов. Мустьерские орудия явились дальнейшим развитием старой ашельской технологии и культуры рубил — чопперов, они изготовлялись от 100 до 35 тысяч лет назад в Европе. Северной Африке и Юго-Западной Азии.
Европейские неандертальцы смогли пережить суровые зимы ледникового периода только благодаря тому, что создавали для себя теплый микроклимат с помощью одежды и обогреваемых жилищ. В некоторых местах естественными укрытиями от холода являлись пещеры.
Захоронения, ритуалы и зачатки искусства говорят о том, что неандертальцы в большей степени обладали самосознанием, общественными интересами и в целом были более способны к абстрактному мышлению, чем их предок Homoerectus.
Неандертальцы явились первыми людьми, которые систематически хоронили своих мертвых. Имеются несомненные доказательства того, что погребение умерших было обрядом. Скелеты лежали в ямах, вырытых в полу пещер. Многие были уложены в позу спящего человека и снабжены предметами, которые, как считалось, должны были сопутствовать умершему в его могиле: от каменных орудий и кусков жареного мяса до каменных подушек, подстилки из лесного хвоща… Они проявляли также и сострадание друг к другу. Но все же, подобно своим предшественникам, эти люди, по-видимому, иногда ели друг друга, раскалывая черепа и кости конечностей, чтобы извлечь мозг.
Трудовая теория антропогенеза Ф. Энгельса. Факторы и критерии гоминизации
Гоминизация (от лат. "Ното"—человек)—процесс очеловечения: обезьяны—от появления первых специфически человеческих особенностей до возникновения вида человека разумного (Н. sapiens). Это комплексный феномен, включающий развитие прямохождения, прогрессивные преобразования мозга и особенно его эволюционно более позднего отдела — неокортекса (неэнцефализация), адаптацию руки к трудовой деятельности, изменения зубочелюстного аппарата, формирование членораздельной речи, концептуального мышления, сознания, интеллекта, перестройку онтогенеза, развитие социальной организации, материальной культуры и другие аспекты.
Решающим фактором гоминизации явилась трудовая деятельность. Трудовая теория антропогенеза в своем первоначальном классическом варианте была изложена Ф. Энгельсом в его известной работе "Роль труда в процессе превращения обезьяны в человека", написанной в 1873—1876 гг. и впервые опубликованной в 1896 г. Основу концепции Ф. Энгельса составляет понятие о труде как первом основном условии всей человеческой жизни, и притом в такой степени, что мы в известном смысле можем сказать, что "труд создал самого человека" (Маркс К., Энгельс Ф. Соч. 2-е изд. Т. 20. С. 486). В этой работе устанавливается последовательность основных этапов гоминизации с выделением прямохождения как решающего шага на пути очеловечения обезьяны; дается определение руки как органа и продукта труда; рассматривается возникновение звукового языка и членораздельной речи, человеческого мышления как следствия общественного развития; подчеркивается качественное своеобразие антропогенеза как процесса активного приспособления человека к среде, экологическое превосходство человека разумного над другими видами.
Трудовая теория антропогенеза успешно выдержала проверку временем, получив подтверждение всей совокупностью новых палеонтологических и археологических аргументов. Таковы бесспорные факты опережающих темпов развития прямохождения по сравнению с другими структурными комплексами гоминизации и доказанного появления первых орудий задолго до формирования современного человека и даже питекантропов.
Основное положение трудовой теории антропогенеза—о решающей роли изготовления ору дий в процессах гоминизации—разделяется ныне подавляющим большинством антропологов мира, хотя распространенное в зарубежной антропологии понятие "культурной адаптации" и не вполне соответствует основным принципам трудовой теории; в этом случае речь идет обычно об "аутокаталитической реакции" или "кибернетическом механизме" обратной связи между развитием биологии (мозга) и культуры при ведущей роли генетических факторов.
Главное внимание уделяется сейчас поиску причин, обусловивших переход к прямохождению как ключевой адаптации предков человека, а также проблеме критериев гоминизации, "грани", разделяющей предчеловеческую и человеческую фазы эволюции.
Формирование семейства гоминид было связано с освоением предками человека новой, более обширной эколого-адаптивной ниши, возникновением принципиально нового способа адаптации, при котором выживание отдельных особей и их групп стало решающим образом зависеть от применения и изготовления орудий. Этот "выход" предков человека из природных экологических систем происходил на фоне все усиливавшегося вымирания человекообразных обезьян в плиоцене.
Предложен ряд гипотез о факторах, которые могли вызвать первоначальные изменения в морфологии или поведении предковой формы и повлиять на ход гоминизации и ее темпы. Среди них: кратковременное повышение фона радиации, обусловленное тектоническими перемещениями, разломами земной коры и усиленным вулканизмом, инверсии геомагнитного поля Земли, иные стрессовые ситуации, изоляция, гетерозис, а также вызванные изменениями экологической обстановки перемены в поведенческих реакциях, начиная с пищевого поведения и "стратегии размножения" и кончая "предкультурным поведением".
Важным и даже первостепенным фактором гоминизации многие исследователи считают развитие механизма защиты с помощью бросания камней и особенно охоту, которая могла повлечь за собой переход к бипедии, изготовление орудий, развитие руки и мозга. Вместе с тем, изучение костей животных в Олдувае и Кооби Фора из слоев с остатками австралопитеков и ранних Ното показало редкость следов расчленения туш каменными орудиями и преобладание использования падали. Видимо, активная охота особенно на крупных животных стала основным способом добывания мяса позднее.
Расхождения существуют в оценке роли естественного отбора в процессе гоминизации вообще и при формировании поведения определенного типа. В частности, высказывается сомнение в возможности применения дарвиновской теории для объяснения конкретных свойств, сформировавшихся в эволюции человека, основной особенностью которой являлось возникновение новых социальных качеств, приведших к резкому ослаблению естественного отбора; важнейшими факторами считаются миграции и смешения. Однако на ранних этапах антропогенеза естественный отбор, видимо, играл весьма существенную роль в реализации и закреплении качественных изменений, в том числе, возможно, и в развитии большей социальности. Указывается и на вероятную роль в эволюции человека полового отбора, о чем писал еще Ч. Дарвин: например, в становлении бипедии как адаптации к добыванию и доставке мясной пищи, ранее проявлявшейся у самцов, которые благодаря этому получали преимущественный шанс на оставление потомства.
Одним из факторов, обусловивших изменения поведения в ходе гоминизации, была нейроэндокринная система, которая могла быть точкой приложения отбора. В частности, возможна роль первичной реорганизации структур лимбической области, в известной мере контролирующей эмоциональное поведение, а также и некоторых других структур, участвующих в становлении и развитии социального и коммуникативного поведения предков человека.
Возрастающие требования к регуляторной деятельности мозга и факторы стресса, усиливающие изменчивость регуляторных систем, могли через механизм дестабилизирующего отбора (Беляев, 1982) способствовать значительному ускорению темпов эволюции нервной системы и поведения.
Другая позиция исходит из первичной роли крупных мутаций, вызвавших значительные анатомические изменения при переходе к прямохождению. Повышение уровня радиации в Восточной и Южной Африке привело к появлению ранних гоминид, тогда как "необлученные" приматы из Экваториальной и Западной Африки морфологически не изменились (Матюшин, 1982). Речь идет таким образом о связи активных проявлений мутагенных факторов, прежде всего в области Восточноафриканского рифта, со временем анатомических преобразовании обитавших здесь гоминоидов (прямохождение, потеря крупных клыков, увеличение мозга). Палеонтологические материалы пока не очень согласуются с этой гипотезой, так как мозг ранних австралопитеков имел вполне понгидные размеры, зубная система также характеризуется многими примитивными чертами, включая довольно развитые клыки,. а пропорции конечностей близки к их типу у генерализованных антропоидов, достаточно распространенному в предшествовавшую эпоху. Сказанное, разумеется, не означает отрицания важной роли мутагенных факторов и мутаторных генотипов в эволюции человека, но не соответствует представлению о "грубых" анатомических изменениях как результате макромутаций у плиоплейстоценовых гоминид.
Несомненно, что возникновение человеческой линии эволюции было уникальным феноменом, возможность которого определялась стечением благоприятных обстоятельств в данном месте и в данное время; это крупнейшее событие в эволюции органического мира невозможно объяснить какой-то одной причиной или предпосылкой, хотя в основе его, вероятно, были изменения экологической обстановки и употребление орудий как адаптация к жизни в саванне. Невозможность точного воспроизведения подобной ситуации в последующем объясняет уникальность человеческого рода на нашей планете.
Все рассмотренные выше факторы так или иначе связаны с экологией, с изменениями природной обстановки. Резкий сдвиг в глобальном состоянии природных комплексов произошел на рубеже плиоцена и сопровождался значительным похолоданием и вытеснением лесов открытыми травянистыми пространствами типа саванны. По-видимому, к этому времени относится возникновение ранних двуногих человекообезьян—австралопитеков. В позднем плиоцене (3—2 млн лет назад) отмечается дальнейшее похолодание, аридизация климата, расширение площади африканских саванн. С этим палеоклиматическим рубежом связывается выделение линии Ното. Распространение открытых пространств могло стимулировать переход к двуногости; известно, что спорадическая двуногость чаще наблюдается в популяциях шимпанзе из более открытых лесистых местностей, чем из джунглей. В условиях африканской саванны полуназемные мартышкообразные обезьяны (анубисы, толстотелы) тоже достаточно часто встают на короткое время на задние конечности. В филогенезе высших форм животного мира поведение может выступать в качестве "лидера" эволюционного процесса, возникая эволюционным путем и представляя собой адаптивное явление. Прямохождение как новый способ локомоции есть часть общей стратегии вида, направленной на его выживание, по нельзя и слишком упрощать картину зависимости между экологическими изменениями и становлением бипедии, т. е. смена лесов саваннами в Африке была мозаичной во времени и пространстве.
Качественные различия между современным человеком — воплощением высшего этапа развития материи, носителем ее социальной формы, разумным существом, обладающим сознанием,—и самыми развитыми современными обезьянами очевидны. Значительно сложнее оказывается провести грань между первыми гоминидами и их животными предками в "вертикальном аспекте". В свете трудовой теории антропогенеза наиболее адекватным представляется критерий различия человека и животного по следам специфически человеческой деятельности, изготовления орудий труда. Однако для решения таксономических задач археологический критерий применим далеко не всегда, так как ископаемые остатки гоминид отнюдь не во всех случаях сопровождаются каменными артефактами, к тому же не было и полной синхронности в развитии морфологических комплексов гоминизации и материальной культуры. Возможна также ассоциация орудий более чем с одним морфотипом гоминид, как, например, в 1 слое Олдувая (зиндж и презинджантроп). Согласно наиболее распространенному мнению, таксономический критерий семейства гоминид и его подразделений должен быть морфологическим, палеонтологическим. Только такой критерий может явиться основой для определения филогенетических взаимоотношений и позволяет объединить в рамках единой системы неонтологические и палеонтологические таксоны.
Морфологический критерий гоминид базируется на развитии важнейших структурных комплексов гоминизации, по которым человек в наибольшей мере отличается от других приматов и которые, следовательно, подверглись самым значительным эволюционным изменениям (прямохождение, мозг, рука, дентиция). Все эти системы в той или иной степени отражают и изменения в поведении, степень адаптации к трудовой и культурной деятельности. Существует довольно широкий спектр мнений по поводу нижней границы семейства гоминид, от признания первым гоминидом понгидообразных рама- и сивапитеков до точки зрения, что истинным гоминидом является только Н. sapiens. Здесь следует иметь в виду, что таксономический (биологический) подход к оценке своеобразия человека неизбежно отличается от философского, который делает главный упор на рассмотрение общества (В. П. Алексеев, 1984). Оставаясь в рамках систематики, нельзя лишать человека филогенетических корней, отрывать "готового человека" от длинной цепи его предков — "формирующихся гоминид".
Многие специалисты считают первым бесспорным гоминидом Н. Наbilis, чей гоминидный статус доказывается прямохождением, высоким уровнем церебрализации, признаками орудийной деятельности на кисти и ассоциацией с орудиями олдувайской культуры. Большинство систематиков включают в семейство гоминид и австралопитеков как двуногих гоминоидов, уже освоивших в той или иной мере новую адаптивную нишу и, следовательно, прибегавших к использованию, а затем и изготовлению искусственных средств культурной адаптации в новой экологической обстановке или потенциально способных к такой деятельности. Таким образом, за основу принимается морфологический комплекс прямохождения, который формировался опережающими темпами сравнительно с другими системами гоминизации и являлся первым и необходимым условием перехода к трудовой деятельности. Австралопитеки уже были высокоразвитыми прямоходящими гоминоидами, т. е. удовлетворяли главному условию систематического критерия гоминид. Уровень развития мозга, и особенно руки, у них был тоже выше, чем у современных понгид.
Принципиально важно, что, по имеющимся данным, не исключена и возможность употребления, подработки или даже спорадического изготовления австралопитеками примитивных орудий. Так, в Чесовандже-(Северная Кения) богато представленная олдувайская культура из 1000—1500 орудий встретилась в сочетании с тероморфным черепом массивного австралопитека, тогда как остатков более прогрессивных гоминид не обнаружено; в Сварткрансе (ЮАР) олдувайские орудия найдены вместе с многочисленными остатками парантропов и крайне скудными фрагментами костей предполагаемых представителей рода Ното (телантропов), хотя обычно большое количество каменных орудий ассоциируется с преобладанием хабилисов над массивными австралопитеками, и наоборот, как, например, в Олдувае, области Вост. Туркана и других местах. Поэтому нельзя полностью исключить предположение, что массивные австралопитеки тоже могли по временам использовать и даже изготовлять орудия, однако эта "культурная деятельность", видимо, не получила у них дальнейшего развития скорее всего из-за выраженной растительноядности и незначительной роли охоты у этих гоминид.
Довольно загадочной остается пока и ситуация в Хадаре: олдувайские орудия древностью 2,5 млн лет обнаружены in situ в верхней части горизонта Када Хадар, где вообще отсутствуют гоминиды такого возраста. На западном берегу р. Гона открыты отходы производства из 18 каменных изделий, предположительно родственных олдувайскому комплексу, но еще большей древности. В Омо описана так называемая "кварцевая культура" на отщепах с редкими следами ретуши и употребления, видимо представляющая более примитивную стадию, чем олдувайская. Следовательно, возможно, что типичным находкам олдувайской культуры предшествовала более ранняя "предолдувайская" ("шунгурская") стадия—древнее 2 млн лет назад. Предполагается, что ее творцами могли быть существа, близкие к афарским гоминидам.
Вырисовывается альтернатива: или принятие гипотезы, что изготовление древнейших орудий было стигматом только линии Ното, и дальнейшее удревнение последней, или допущение возможности изготовления орудий и другими двуногими, по крайней мере некоторыми из австралопитеков.
Нужно отметить, что появление новых фактов из области первобытной археологии, а именно удревнение первых изделий из камня, сокращает хронологический разрыв между двумя важнейшими феноменами гоминизации—прямохождением и началом орудийной деятельности, существование которого побуждало некоторых исследователей к отрицанию причинно-следственной связи между ними. К тому же многие специалисты принимают гипотезу Р. Дарта о широком использовании австралопитеками костей, рогов и зубов животных—так называемой "остеодонтокератической культуры", отражавшей зачаточные формы орудийной деятельности.
Если исходить из концепции, что именно принципиально новый способ адаптации послужил основой выделения линии гоминид, и рассматривать переход к прямохождению в прямой связи с орудийной деятельностью как ее предпосылку (а, возможно, отчасти и следствие),. то надо признать, что все двуногие могли в принципе использовать орудия. В этом случае к семейству гоминид следует относить всех двуногих высокоразвитых гоминидов как освоивших в той или иной степени новую адаптивную нишу. Поэтому на современном этапе изучения этой проблемы, видимо, правильнее включать австралопитеков в качестве особого подсемейства Аuvstralopithecinae в семейство гоминид и рассматривать их как древнейших гоминид—"австралантропов", хотя формально такая замена термина и невозможна. Род Ното выделяется в подсемейство Ноmininae.
Следовательно, если речь идет о морфологической грани между высокоразвитой обезьяной и двуногой человекообезьяной — австралопитеком, то она достаточно четко выявляется по комплексу прямохождения. Что же касается существования грани в пределах самих двуногих человекообезьян между существами, уже перешедшими к изготовлению орудий, и еще не перешагнувшими этот рубеж, то этот вопрос пока решается, скорее, отрицательно. Среди ранних австралопитеков есть популяции без сопутствующих орудий, как, например, в Летоли, где следы орудийной деятельности не обнаружены, несмотря на тщательные поиски. Хотя биологические предпосылки к трудовой деятельности были уже сформированы у австралопитеков, которые сделали решающий шаг на пути перехода к адаптации с помощью культуры, но это был, несомненно, весьма длительный и неравномерный процесс. Поэтому нельзя ожидать на самых ранних его этапах четких структурных индикаторов перехода к трудовой деятельности.
В силу еще не вполне понятных причин наибольшую способность к культурной адаптации проявила линия Ното. Ее отличительной чертой стало прогрессивное увеличение мозга и усложнение его структуры. В филогенезе гоминид существует значимая статистическая связь между геологическим возрастом находки и массой мозга в период от 4 до 2 млн лет назад, но с отступлениями от линии регрессии, типичными для криволинейной зависимости. Экспериментально было показано, что масса мозга и многие его структурные и химические характеристики (толщина коры, размеры ядер нейронов, cинаптогенез, химический состав нервной ткани, активность ферментов и др.) меняются в зависимости от условий содержания животных и, следовательно, являются приспособительными. Возможно поэтому, что аналогичные сдвиги наследственной природы могли поддерживаться механизмом естественного отбора. На ранних стадиях эволюции гоминид это могло приводить к дивергенции в размерах и иных параметрах мозга, которая прослеживается, например, у современников—питекантропов и массивных австралопитеков из Кооби Фора. Но в последующем возрастающая "буферная" роль формирующейся культурной среды, вероятно, препятствовала проявлениям кладогенеза, основным путем эволюционных изменений в линии гоминид становится анагенез, или ароморфоз.
В науке нет вполне однозначного мнения о роли кладо- и анагенеза на ранних этапах эволюции гоминид. Большинство специалистов приняли в целом кладогенетическую схему Д. Джохансона—Т. Уайта c ключевым положением в ней афарского австралопитека (или вообще австралопитека грацильного типа). Но в принципе при видообразовании во времени эволюция может происходить и без внутривидовой дифференциации—как историческое преобразование всей массы ("пласта") особей данного вида (Шмальгаузен, 1968). Однако, как уже говорилось выше, общая картина распределения палеоантропологических материалов плиоплейстоценовой эпохи находится в большем соответствии с концепцией кладогенетической эволюции. Представители вида Н. Наbilis и даже древнейшие Н. егесtus (последние на протяжении нескольких сотен тысяч лет) сосуществовали с австралопитеками массивного типа. Так, например, в верхнем горизонте формации Кооби Фора в Кении найдены морфологически четко различимые формы—архантроп (череп 3733) и поздний зиндж (череп 406). В Олдувае-сосуществовали Н. Наbilis (ОН-7, ОН-13, ОН-16) и А. boisei (ОН-5,. Пениндж). В Стеркфонтейне и Сварткрансе, видимо, тоже одновременно обитали представители родов Avstralopithecus и Ното. Сосуществование двух видов австралопитеков считается доказанным в течение по крайней мере 1 млн лет. Если подтвердится датировка 2,5 млн лет для восточноафриканского массивного австралопитека, окажется, что специализация массивных форм существовала уже у истоков дифференциации линий человека и австралопитеков, что, скорее, свидетельствует в пользу мнения о более ранней дивергенции этих линий и независимой эволюции линии зинджа.
Проблема сосуществования является одной из наиболее дискуссионных. Известно, что видообразование в пределах ареала расселения одного вида (симпатрическое), считавшееся вообще достаточно редким: событием, получает все новые подтверждения в разных группах позвоночных. Имеются симпатрические виды и у современных приматов, например кошачий и желтый лемуры, нередко встречающиеся на одном и том же лесном участке, но различающиеся образом жизни. Однако сосуществование разных видов у ранних гоминид в случае симпатрии иногда подвергается сомнению из-за малой вероятности первичной "культурной адаптации" в различных линиях—"гипотеза одного вида".
В основе этой гипотезы предположение, что морфологическое сходство между ранними гоминидами не является результатом параллелизма, но определяется самой природой культурной адаптации; случаи же морфологической гетерогенности объясняются с позиций аллометрии как следствие высокой внутрипопуляционной изменчивости. В соответствии с принципом конкуретного исключения Г. Ф. Гаузе (1934) утверждается, что два сходных вида, использующих один и тот же жизненно важный ресурс, не могут занимать одну эконишу и, таким образом, на Земле не могло одновременно существовать более одного вида прямоходящих предков человека.
В принципе "гипотеза одного вида" совместима и с кладогенезом, и с анагенезом. В первом случае речь может идти о "стволе с короткими боковыми ветвями", во втором—это модель простой прямолинейной трансформации единого исходного варианта "по вертикали", которую пока трудно согласовать с реальными палеонтологическими фактами. Кстати, уже давно было замечено, что при крайней "вертикальной" тенденции весь органический мир можно причислить к одному виду. Следует также признать, что эволюция приматов изобилует примерами параллелизмов и конвергентного развития иногда весьма далеких в систематическом отношении форм (например, би-лофодонтия у церкопитекоидов и плейстоценовых просимий Мадагаскара — археолемуров).
Можно полагать, что фундаментальная ниша Ното уже с самого начала не была вполне идентичной нише австралопитека. Вероятно,. разные виды могли сосуществовать на одной территории, если каждый: из них имел свое "поле адаптации", в том числе различные комплексы форм поведения, обеспечивавшие неодинаковое использование природных ресурсов (Бунак, 1980). В основе дивергенции различных линии плиоплейстоценовых гоминид, прежде всего Ното и собственно австралопитеков, могли лежать какие-то механизмы изоляции: генетической (пониженная жизнеспособность гибридов), экологической (различные местообитания в пределах одной территории, разные пищевые предпочтения и специализации). Так, у массивных австралопитеков предполагается преимущественная зерноядность, питание семенами, стручками, корневищами и прочим, тогда как у ранних Ното, очевидно, преобладала более разнообразная и калорийная пища: плоды, мясо, костный мозг. Изолирующим фактором могли явиться и особенности репродуктивного поведения, слабая взаимная привлекательность, делавшие скрещивания настолько редкими, что видовая специфика сохранялась, а также общественная структура групп.
Проблема сосуществования возникала и на последующих этапах эволюции, уже в пределах рода Ното. "Стадиальная гипотеза", согласно которой в каждую геологическую эпоху на Земле был представлен только один вид гоминид, нуждается в переосмыслении. Хорошо известно, что на границах "стадий" различные формы гоминид сосуществовали в достаточно широких временных интервалах. Сейчас трудно представить эволюцию гоминид как простую трансформацию единого исходного варианта. Конечно, этапы (фазы) существовали в филогенезе человека, но они не имели монотонного характера, их представители морфологически были разнообразными и могли различаться по типу культуры. Центростремительное воздействие социальных факторов, возраставшее в ходе гоминизации, приводило к закреплению превосходства ароморфной эволюции над проявлениями адаптивной радиации, однако эта тенденция к "канализированию" была значительно менее выражена в плиоплейстоценовую эпоху, когда слабее проявлялись факторы сдерживания кладогенеза (Зубов, 1983, 1985).
Общая картина морфологической эволюции гоминид еще далеко не выяснена. Пока нельзя говорить о какой-то единой общепризнанной "модели", с помощью которой можно было бы описать этот процесс. Даже применительно к такому хорошо изученному признаку, как масса мозга, предложено несколько различных моделей: постепенного возрастания массы с постоянной скоростью, аутокаталитического ускорения темпов по мере возрастания мозга и, наконец, "эпизодического увеличения", когда быстрые изменения чередуются с периодами замедленного развития. Для процесса эволюции гоминид в целом также в основном разрабатываются модели постепенного филетического развития ("градуализм") и "прерывистого равновесия", со сменой длительных периодов относительной стабильности видов (стазисы) периодами очень быстрых изменений, приводящих к формированию новых видов.
Палеоантропологические материалы, по-видимому, допускают обе эти возможности. Так, относительно стабильными видами могли являться афарский австралопитек и Н. Егесtus, примером сравнительно быстрого симпатрического видообразования считают "переходные виды"—Н. Наbilis и, может быть, архаического Н. sapiens. Иногда применительно к гоминидам используется даже понятие "катастрофическое видообразование", под которым подразумевается радикальное изменение направления эволюционного процесса, внезапный эффект постепенного сдвига соотношения сил в неожиданном направлении (массивные австралопитеки). Скачкообразная ("квантовая") эволюция может связываться с освоением новых адаптивных ниш или биосферными кризисами (Красилов, 1986), поскольку значительные изменения среды могут иметь большое селективное значение и повысить скорость эволюционных изменений.
Процесс антропогенеза следует рассматривать как диалектическое единство прерывистого и непрерывного. Видимо, элементы скачкообразности (дискретности), обусловленной мутациями регуляторных генов или "транспозиционными взрывами" мобильных генов, способных. перемещаться в наследственном материале, могли сочетаться с постепенностью развития, опосредованной накоплением малых мутаций под. контролем естественного отбора. Противоречивость рассмотренных выше концепций и соответствующих им моделей ранних этапов антропогенеза является результатом не только неполноты палеонтологической летописи, но и отражением реальной диалектической противоречивости эволюционного процесса.
5. Кроманьонский человек. Человек современного типа
Кроманьонский человек и человек современного физического типа (неоантропы)
Под этим названием понимают как ископаемые формы человека современного физического типа, так и ныне живущих людей. В 1868 г. в пещере Кро-Маньон (Франция) был найден скелет человека, объем черепной коробки которого составлял 1800 мл. Для него было характерно отсутствие надбровных валиков (они разделились на две надбровные дуги), наличие подбородка, прямой лоб. Рост его составлял около 180 см. Этого человека назвали кроманьонцем. В дальнейшем сходные ископаемые формы были найдены также на территориях Китая, Палестины, Чехии, России и в других странах Европы, Африки и Азии. Средний возраст кроманьонцев составлял около 30 лет.
Считалось, что человек современного физического типа появился 40—50 тыс. лет назад. В соответствии с новыми данными этот срок определен в пределах 100-200 тыс. лет назад. Кроманьонцы не отличаются от людей современного физического типа. Если бы кроманьонец, одетый в современную одежду, шел по улице или находился бы в учебной аудитории, его было бы трудно выделить среди окружающих людей. В их пещерах были найдены стрелы, гарпуны, иглы, что указывает на усовершенствованные методы изготовления орудий. Они были искусными охотниками и ответственны за истребление мамонтов, саблезубых тигров, бизонов и др. Примерно около трети родов млекопитающих было уничтожено еще 50 тыс. лет назад. Предполагают, что резкое уменьшение диких животных способствовало переходу людей от охотничьего сообщества к сельскохозяйственному. На стадии существования неоантропа на развитие человека стали оказывать все большее влияние культура и социальные закономерности.
6. Время и место формирования человека разумного
Прародина человечества
Широкое распространение высокоразвитых гоминоидов в Афроевразии на рубеже миоцена и плиоцена дает возможность в принципе рассматривать каждый из этих континентов как возможную зону очеловечения—древнейшую прародину человечества. Однако накопленные к настоящему времени палеонтологические, археологические и биомолекулярные факты позволяют заключить, что и ныне, как и во времена Ч. Дарвина, наиболее вероятной прародиной человечества представляется Африка, и в первую очередь область Восточноафриканского рифта—гигантского горного разлома, протянувшегося с севера на юг, от Ближнего Востока до Южной Африки, на 8000 км. В пользу этого мнения можно привести следующие аргументы: Восточная (как и Южная) Африка являлась областью обитания и широкого расселения древнейших гоминид — австралопитеков и ранних представителей рода Ното,, здесь же обнаружены и самые ранние каменные орудия. Наиболее древняя пока датировка получена для артефактов из стоянки Гона в Афаре (Эфиопия): 2,70—2,89 млн лет (Дж. Харрис, Д. Джохансон, 1982).
Указанные районы обладали комплексом природно-экологических факторов, вполне соответствующим теоретическим представлениям об экологической нише первых гоминид. Сюда относятся: а) повышенная геологическая активность с периодическими смещениями земной коры;, область Восточноафриканского рифта изобилует вулканами и следами прошлой вулканической деятельности в виде мощных слоев пепла;. вулканизм сопровождался изменениями ландшафта, сменой фаун, периодическими уничтожениями растительности, которые могли стимулировать переход к всеядности; б) повышенный уровень ионизирующей радиации, инверсии геомагнитного поля Земли, что могло способствовать возникновению и закреплению мутаторных генотипов; в) преобладание богатой источниками водоснабжения, сравнительно открытой местности типа саванны или лесосаванны (Омо, Олдувай, оз. Туркана,. Хадар и другие местонахождения Восточной Африки располагались на берегах водоемов), что имело немаловажное значение в изменении поведенческих реакций. Обилие фауны могло побудить наряду с другими причинами к более широкому использованию мясной пищи—важного условия для развития мозга; типичный для многих восточноафриканских стоянок гоминид скалистый ландшафт благоприятствовал зарождению ранних форм изготовления орудий.
Наконец, в Африке представлены и самые ранние стадии эволюции гоминоидов (египтопитек, проконсул и др.); здесь и в настоящее время обитают наиболее близкие к человеку и тесно связанные с ним общностью происхождения понгиды (шимпанзе, горилла). В пользу африканской прародины свидетельствуют и биомолекулярные данные.
В начале XX в. в науке обсуждалась гипотеза о центральноазиатском происхождении человечества. Переход понгидных предков к прямохождению связывался с климатическими изменениями, обезлесением, как следствием горнообразовательных процессов на рубеже третичного и четвертичного периодов. Данные палеогеографии, палеонтологии и археологии не подтвердили эту концепцию. Конечно, Азия не может быть полностью исключена из числа возможных территорий очеловечения, учитывая вероятную тесную связь между Африкой и Азией в период от 18 до 5—6 млн лет назад, как и некоторые находки каменных артефактов олдувайского облика в Азии, впрочем, с недостаточно ясной датировкой. Высказывалось также предположение, что в зону очеловечения могли входить и южные районы Европы, в том числе территория Закавказья. Но, вне сомнения, концепция африканской прародины аргументирована сейчас гораздо лучше, и, по существу, только в Африке и представлены все этапы эволюции человеческой линии. Находки самых ранних гоминид в Южной Евразии пока единичны или вообще сомнительны.
Появление человека явилось величайшим событием четвертичного периода, который предложено именовать "антропогеном" (А. П. Павлов). При определении границы третичного и четвертичного периодов принимают во внимание и данные первобытной археологии и палеоантропологии, хотя понятно, что этот рубеж должен быть универсальным эталоном для датирования событий на всем земном шаре, с учетом всего комплекса биологических, геологических и хроностратиграфических фактов. Что касается палеоантропологических и археологических материалов, то они, как мы видели, больше согласуются с теми из применяющихся сейчас датировок нижней границы антропогена, которые отражают тенденцию к ее удревнению.
1.Время возникновенияЕсли отбросить библейскую легенду о сотворении человека, то вопрос о времени появления современного человека на нашей планете стал занимать умы ученых сравнительно недавно – каких-нибудь 40-50 последних лет, так как до этого обсуждалась в основном древность человеческого рода вообще. Даже в серьезной научной литературе очень долго господствовала тенденцияувеличения геологического возраста Homo sapiens и в соответствии с этим использовались антропологические находки с неясной или недостаточно ясной геологической датировкой. Список таких находок довольно длинен, он постепенно менялся – на место дискредитированных находок становились новые, но все последующие исследования не подтверждали глубокой древности техкостных остатков, которые могут быть отнесены к современному человеку. Гипотеза пресапиенса отражает туже тенденцию, но не получает поддержки со стороны морфологической ; находки на которые она опирается, хотя датированы безупречно и действительно древни, но отнесение их к современным людям , а не палеоантропам вызывает самые серьезные сомнения.Все древнейшие находки в верхне-палеолитических слоях датируются в абсолютных цифрах 25000-28000,а иногда и 40000 лет, т.е. практически синхронны или почти синхронны, находкам наиболее поздних палеоантропов. Едиственное убедительное исключение составляет сделанная в1953 г. А.А.Формозовым находка в Староселье под Бахчисараем. Современный облик обнаруженного в мустьерском слое младенца в возрасте 1.5 лет не вызывает не малейших сомнений, хотя исследовавший его Я.Я.Рогинский и отметил на черепе несколько примитивных признаков: умеренное развитие подбородочного выступа, развитые лобные бугры, крупные зубы. Датировка этой находки в абсолютных цифрах не ясна, но инвентарь, найденный с ней, показывает, что она значительно древнее, чем верхнепалиолетические местонахождения с костными останками современных людей. Этим фактом твердо устанавливается синхронность древнейших форм современного человека и позднейших групп палеоантропов, их существование на протяжении довольно значительного отрезка времени. На первый взгляд это обстоятельство кажется несколько неожиданным, но стоит подумать, как оно теряет кажущуюся парадоксальность: перестройка морфологии – процесс длительный, коль скоро мы принимаем наличие неандертальской фазы в эволюции человека, мы должны сделать вывод о формировании отличительныхморфологических особенностей человека разумного в недрах групп палеоантропов,а раз так, то существование палеоантропа и современного человека на каком-то отрезке времени представляется и теоретически неизбежно. В рамках такого взгляда легко находит себе объяснение, отмеченное Я.Я.Рогинским сходство черепа из Староселья с детским черепом из пещеры Схул в Палестине, где найдены морфологически прогрессивные скелеты неандертальцев. Кстати сказать, сосуществование древних примитивных и более поздних морфологическихпрогрессивных форм составляло характерную черту эволюции гоминид практически на всех этапах их истории.Итак, формирование человек разумного на базе палеоантропа привело к сосуществованию поздних прогрессивных форм неандертальцев и зарождающихся пока малочисленных групп современных людей на протяжении нескольких тысяч лет. Процесс вытеснения старого вида новым был довольно длительным, а, следовательно, и сложным.Локальные варианты внутри неандертальского видаРешение проблемы центров возникновения современного человека неразрывно связано с систематикой неандертальского вида, с числом локальных вариантов внутри него , а главное - с их систематическим положением и отношением к прямой линии человеческой эволюции. Все эти вопросы получили широкое освещение в антропологической литературе.В пределах неандертальского вида в нашем понимании можно выделить несколько групп, имеющих морфологическую, географическую и хронологическую специфику. Европейские неандертальцы, составляющие компактную географическую группу, распадаются в соответствии с распространенным мнением на два типа, своеобразных морфологически и существовавших в разноевремя. Литературная традиция связывает выделение этих типов с именем Ф.Ванденрайха, выступившего со статьей на эту тему в 1940 году, однако М.А.Гремяцкий осуществил его раньше в докладе, сделанном в Институте антропологии МГУ в 1937 году. К сожалению, текст этого доклада был опубликован лишь через 10 лет и остался малоизвестным западноевропейской иамериканской науке. Выделенные типы именуются различными исследователями "классическими", или "типичными" и "атипичными" неандертальцами, "группой Шаппель и Ферасси" и "гуппой Эрингсдорф" по названиям мест важнейших находок и т.д. Вторая группа по сложившейся традиции якобы более ранняя, она датируется перидом рисского оледенения (около 110-250 тыс. лет назад) и рисс-вюрмским межледниковьем. Первая группа относится к более позднему периоду и датируется началом и серединой вюрмского оледенения (от 70 до 110 тыс. лет назад). Хронологические различия сопровождаются морфологическими, но последние парадоксальным образом не соответствует ожидаемым, и характеризуют обе группы в обратном порядке по сравнению с геологическим возрастом: более поздние неандертальцы оказываются более примитивными, более ранние – прогрессивными. Мозг у последних, правда, несколько меньше по объему, чем у поздних неандертальцев, но более прогрессивен по строению, череп выше, рельеф черепа меньше (исключение составляют сосцевидные отростки, развитые сильнее - типичный человеческий признак), на нижней челюсти намечается подбородочный треугольник, размер лицевого скелета меньше.Происхождение и генеалогические взаимоотношения двух этих групп европейских неандертальцев много раз обсуждались с самых разных сторон. Была высказана гипотеза, согласно которой поздние неандертальцы приобрели свои отличительные особенности под влиянием очень холодного, сурового ледникового климата в условиях Центральной Европы. Их роль в формировании современного человека была меньше, чем ранних, более прогрессивных форм, которые и явились прямыми и основными предками современных людей. Однако против такой трактовки морфологии и генеалогических отношений хронологических групп в составе европейских неандертальцев выдвигалось то соображение, что географически они были распространены на одной и той же территории и ранние формы также могли быть подвержены воздействию холодного климата в приледниковых районах, как и поздние. Приводились и общетеоретические возражения против попытки рассматривать поздних палеоантропов как боковую ветвь, не принимавшую совсем или принимавшую малое участие в сложении физического типа человека разумного. Таким образом, вопрос о степени участия обеих групп европейских палеоантропов в процессе формирования Homo sapiens остается открытым; скорее следует ожидать, что поздние неандертальцы также могли явиться непосредственной базой для сложения физического типа современного человека в Европе.Любопытно отметить, что перечисленные выше различия констатированы разными авторами преимущественно при сравнении отдельных черепов "на глазок", при игнорировании того очевидного обстоятельства, что классические неандертальцы представлены преимущественно мужскими, атипичные – женскими черепами. Если же принять во внимание это обстоятельство и вычислить средние по группам, то при ничтожном числе наблюдений, которым представлена каждая группа, невозможно подтвердить приведенный перечень различий сопоставлением средних: различия являются случайными и разнонаправленными. Их оценка с помощью простых статистическихприемов показала, что суммарные различия примерно равны тем, которые разделяют современные расовые ветви, и, следовательно, говорить о двух группах разного уровня эволюционного развития в составе неандертальского вида с морфологической точки зрения нет оснований. Не больше этих оснований географии находок (ареолы обеих групп примерно совпадают) и их хронология (время их существования также более или менее совпадает в широких границах). Разумеется, в составе европейских неандертальцев могли существовать локальные варианты, приуроченные к отдельным популяциям и их группам, но в целом неандертальское население Европы образовывало достаточно однородную группу. География этой группы не полностью соответствует географическим рамкам Европы, и по этому мы лишь условно можем называть ее европейской. Расчеты и сравнительные сопоставления продемонстрировали сходство с этой группой также известных нам североафриканских находок из Джебел Ирхуда и одного из черепов, найденного при раскопках пещеры Схул в Палестине, того черепа, который обозначается в научной литературе как Схул IX. Таким образом, европейская группа территориально охватывала Северную Африку и какую-то прибрежную часть территории восточного Средиземноморья уже в пределах азиатского материка.Однако и на территории Европы, в самых южных ее районах, проживали формы, которые не могут быть по морфологическим признакам включены в европейскую группу. Речь идет о черепе из Петралоны в Греции. Череп был найден в 1959 году одним из рабочих, принимавших участие в раскопках Петралонской пещеры, и поэтому его стратиграфическое положение, аследовательно, и хронологическая датировка не вполне ясны. Своеобразие его морфологии отразилось в оценках его положения внутри неандертальского вида. Авторы первых описаний и измерений П.Коккорос, А.Канеллис и А.Саввас, как всегда бывает в таких случаях, ограничились лишь самым предварительным диагнозом и отнесли череп к группе классических неандертальцев Европы. Совершенно очевидно, что в этом сказались гипноз бесспорно примитивных особенностей строения черепа по сравнению с современным, его бесспорно неандертальские признаки. Однако реферировавший работы греческих ученых М.И.Урысон не согласился с их диагнозом и впервые отметил наличие признаков, сближающих череп из Петралоны с африканскими формами. Окончательный вывод М.И.Урысона: петралонский череп представляет собой промежуточную форму между африканскими и классическими европейскими неандертальцами. Э.Брайтингер в докладе на VIII Междунарадном съезде антропологических и этнографических наук в Москве в августе 1964 года специально подчеркнул замеченное М.И. Урынсоном сходство с африканскими формами.Включившийся позже в изучение петралонского черепа А.Пулянос, используя сначала предшествующие, а затем и самостоятельные измерения черепа, оспорил эту точку зрения и сближал череп сначала с европейскими неандертальцами, подчеркивая, правда, его своеобразие. В ряде его работ, посвященных не столько детальному сравнительно-морфологическому исследованию черепа, сколько тщательной характеристике обстоятельств его находки, включая геологическое и палеонтологическое изучение пещеры, хронологический возраст черепа определяется в 700000 лет и предполагается, что он принадлежал представителю самостоятельного вида внутри рода архантропов или питекантропов - Archantropus europeus petraloniensis. Номер греческого журнала "Антропус", в котором опубликованы эти работы А. Пуляноса, содержит большое число палеонтологических, стратиграфических и геофизических данных, в целом подтверждая эту версию. И датировка, итаксономический диагноз, если они справедливы, ставят находку на выдающееся место в палеоантропологии Европы, делая ее одной из древнейших. Также датируются с помощью палеомагнитного метода сталактиты, упавшие с потолка пещеры; на одном из них был найден череп. Не будучи лично знакомым, с пещерой и обстоятельствами раскопок, трудно противопоставить что-либо определенное этим выводам, но, логически рассуждая, без специальных доказательств трудно принять точку зрения о полной синхронности возраста сталактитов, упавших с потолка пещеры и черепа. Н. Ксиротирис в докладе на симпозиуме по проблемам антропогенеза , состоявшемся в мае 1981 года в Веймаре в ГДР, привел очень убедительные сомнения в столь древнем возрасте петралонской находки , которая по его мнению, является одной из древнейших неандертальских находок в Европе, но геологическая древность которой, по самым расточительным подсчетам, непревышает 150000-200000 лет.Морфология находки также не свидетельствует об исключительной примитивности петралонского черепа. После того как почти со всех костей черепа были сняты минеральные натеки , он был подвергнут в1979-1980 годах повторному и очень подробному измерению , которое наконец дает достаточно полную сводку размеров без условных поправок на известковые покрытия костейлицевого скелета и черепного свода. На основании сравнительного анализа этих измерений исследователи приходят с выводу, что находка имеет ряд примитивных признаков, но все же, как и все американские авторы, пользующиеся таксонометрической схемой Эмайра, включают ее втаксонометрическую категорию Homo sapiens .К.Стнгер еще ранее подтвердил этот диагноз с помощью суммарных статистических сопоставлений. И статистическое ,и географическое сравнение черепа из Петралоны с другими формами показало, что наибольшее сходство он имеет с африканскими неандертальцами, в первую очередь с черепом из Брокен –Хилла. Отдельныечерты сходства с европейскими находками также имеют место, но они не должны нас особенно удивлять: весьма вероятно, что на окраинах ареалов европейских и африканских палеоантропов происходил процесс метизации, приводивший к появлению промежуточных форм. В целом же череп из Петралоны, которому мы посвятили много внимания в связи с продолжающейся дискуссиейвокруг его датировки и таксонометрического места, должен быть включен во вторую африканскую локальную группу внутри неандертальского вида, к характеристике, которой мы переходим.Морфология африканских неандертальцев чрезвычайно своеобразна. Реконструкция так называемого африкантропа, осуществленная Г.Вайнертом, в высшей степени проблематична, так как она базируется на большем числе фрагментов, не полностью или вовсе не соприкасающихся между собой. Гораздо полнее может быть охарактеризовано строение черепов из Брокен-Хилла (замбия), Салданья (Южная Африка) и Афара (Эфиопия). Им свойственно сочетание в высшей степени примитивных особенностей, сравнительно малого объема мозга и его примитивного строения, исключительно мощного развития рельефа черепа, у родезийца (так обычно в палеоантропологической литературе по старому наименованию г. Кабве в Замбии называют череп из Брокен-Хилла) – еще и огромного лицевого скелета с некоторыми прогрессивными признаками. М.А.Гремяцкий был, кажется, первым, кто отметил сходство африканских неандертальцев с черепами из Нгандонга.Но последние, как мы убедились выше, должны быть отнесены не к неандертальской группе, а к группе архантропов. Некоторое сходство их с черепами из Брокен-Хилла и Салданьи отражается только в строении черепной коробки (сильное развитие рельефа черепа, мощный сагитальный валик), так как лицевой скелет сохранился только у черепа из Брокен-Хилла.Другая находка с лицевым скелетом - совсем новая находка черепа неполной сохранности, реконструированного из многих фрагментов , в местности Бодо в Афаре на территории Эфиопии. Датировка черепа – средний плейстоцен, то есть, по мнению авторов находки, примерно в пределах 150000-600000 лет. Хотя измерения черепа еще не опубликованы, но, судя по его строению ,он производит впечатление неандертальского черепа, в общем сходного с другими представителями этого вида. Интерес к этой находке состоит в том, что она подтверждает групповой характер строения лицевого скелета у родезийца. Г.Конрой пишет, что "доминирующей характеристикой лица... является его исключительная массивность". Своеобразие африканской группы, как уже подчеркивалось, несомненно, и она может быть выделена в качестве второго локального варианта палеоантропов. Раньше можно было думать , что хронологически это вариант поздний, по-видимому, частично синхронный наиболее поздним находкам европейских неандертальцев. Но теперь, когда опубликованы новые данные о геологическом возрасте черепа из брокен-Хилла, позволяющие отодвинуть его от современности на 125000 лет, и когда в нашем распоряжении есть череп средне плейстоценового возраста и неандертальского типа из Бодо, геологический возраст всей группы следует увеличить. В этой связи особое значение приобретают некоторые морфологические наблюдения над строением черепной коробки африканских питекантропов, в частности черепа Олдувай II; исключительная массивность черепного рельефа в этом случае дополняется наличием значительного сагиттального валика , исключительно сильно выраженного и на черепах из Брокен-Хилла и Салданьи.Возможно, это морфологический намек на какую-то специфически генетическую связь между африканскими питекантропами и африканскими неандертальцами в пределах одного и того же материка.Третий, вполне четко выраженный вариант в составе палеоантропов - группа Схул (пещера Мугарет-эс-Схул в Палестине раскопанная д. Гаррот в 1931-1932 гг.). Несколько скелетов из этой пещеры синхронные, очевидно, поздним находкам европейских неандертальцев, сразу же обратили на себя внимание чрезвычайно прогрессивным строением. Череп Схул IX, как мы помним исключениз этой группы и включен в группу европейских неандертальцев. Но черепа взрослых особей, обозначенных как Схул IV и Схул V, типичны для этой группы и как раз отличаются прогрессивной морфологией, приближающейся к сапиентному типу. Здесь и высокий свод черепной коробки с относительно мало наклонной лобной костью и большим объемом мозга.Центр происхождения человека
Ученым долго не удавалось обнаружить ископаемые останки древнейших человекообразных обезьян, которые ближе всего стояли бы к питекантропам. Только в конце XIX в. были найдены их останки. Они были обнаружены в разных местах присредиземноморья, Африки, Юго-Восточной и Передней Азии. Однако эти находки сами по себе еще не объясняли процесса происхождения человека. Поэтому главную роль играли не факты, а теории. Появление человека как вида, в частности, объяснялось так. В результате похолодания сократились области тропической растительности, стало больше степей, открытых пространств. Некоторые высокоразвитые обезьяны приспособились к этим новым условиям, что привело к естественному изменению их образа жизни; другие же виды навсегда остались привязанными к тропическому лесу и, надо полагать, дали начало современным человекообразным обезьянам. За последние сто лет в разных местах земного шара, исключая Америку, которая не является зоной происхождения человека, были обнаружены сотни останков ископаемых людей, стоявших на разных ступенях развития. Все это на первый взгляд подтверждает эволюционную теорию происхождения и развития человека. И действительно, человек в своем развитии прошел три стадии: древнейшие люди — архантропы; первобытные люди — палеантропы; современный человек. Таковы основные этапы антропогенеза. Эволюцию человека нельзя оторвать от исторической геологии земли и ее хронологии. Разобраться в проблеме человек — среда — археологическая культура поможет приведенная схема, так как наука о природе и наука, о человеке для палеолита смыкаются. Антропогенез протекал в четвертичный период геологии земли и охватывает: эоплейстоцен (2,5 млн.—350 тыс. лет назад), плейстоцен (350—12 тыс. лет назад), что соответствует палеолиту и фазам ледникового периода. За пределами палеолита находится современный период геологии земли — голоцен. Вопрос о прародине человека, т. е. той части земного шара, где имел место начальный период антропогенеза, решался в зависимости от находок останков австралопитеков, палеантропов и архантропов. Этот вопрос и сейчас вряд ли можно считать окончательно решенным, так как наука постоянно развивается, и каждый год можно ожидать принципиально новых открытий. В науке рассматривались в основном два центра происхождения человека: азиатский (Юго-Восточная Азия) и африканский, причем до открытий в Африке предпочтение отдавалось азиатской прародине человека. В современной науке центральное место отводится "африканской" теории. Останки австралопитека впервые были найдены в 1924 г. Р. Дартом, обнаружившим в Африке череп ископаемого существа. В настоящее время известно несколько основных разновидностей австралопитеков. По физическому строению ископаемые люди отличались от своих ближайших предков тремя основными признаками: прямохождением, приспособленной к тонкому манипулированию кистью руки с противопоставленным большим пальцем, высокоразвитым, относительно крупным мозгом. В пересмотре хронологии и концепции происхождения человека большую роль сыграли М. Лики, Г. Айзек, Д. Кларк, К. Хауэл, Ф. Тобайас. Истоки человечества современная наука связывает с олдувайской культурой, которая была выделена на основе находок , в знаменитом ущелье Олдувай в Танзании. Работы, которые долгие годы проводили там антропологи Луис, Роберт и Мери Лики, увенчались блестящим успехом. Обнаружены останки ископаемых человекообразных существ и примитивные отшепы из кварцита и базальта со следами искусственного скалывания. Известны две разновидности австралопитеков, встречающихся здесь,— презинджантроп и зинджантроп, жившие около 2 млн. лет назад. Они пользовались галечными орудиями труда, сделанными путем очень грубых сколов с поверхности гальки.
Место формирования сапиенса
"Процессы сапиентации, как об этом свидетельствуют палеоантропологические данные, происходили в различных районах Старого Света, хотя и с разной скоростью. Здесь могли играть роль различные обстоятельства, особенности окружающей среды, специфика общественной структуры популяции и т.д. Таким образом, обе гипотезы — множественности центров сапиентации (полицентризм) или ограниченности ее одной достаточно обширной территорией (широкий моноцентризм) — имеют точки соприкосновения. Можно предположить, что, так сказать, "опережающими темпами" она происходила в Восточной Африке Юго-Восточной Европе и на Ближнем Востоке.
Сейчас много споров вызывает проблема приоритета Африки или Европы. Судя по некоторым данным, на древнейшей прародине человечества сапиенс мог появиться на несколько десятков тысяч лет раньше, чем на других территориях. Однако, если и принять эту гипотезу, она вовсе не исключает возможности того: что сапиентация происходила и в других районах Старого Света. Большая часть сторонников полицентризма признает умеренное число центров сапиентации: от двух ("дицентризм") до четырех-пяти.
Высказывалось, правда, и такое соображение: очагов возникновения сапиенса могло быть столько же, сколько было мест формирования культуры верхнего палеолита. Ведь мустьерская культура в основном связана с неандертальцем, а культура верхнего палеолита — с сапиенсом. <...>
Однако жесткой связи между каменным инвентарем и физическим типом не было, не было, видимо и непреодолимой границы между неандертальцем и сапиенсом в культурном отношении. <…>
В пользу полицентризма говорит и почти одновременное появление сапиенса на рубеже верхнего палеолита — примерно 40—35 тыс. лет назад, притом в таких удаленных друг от друга, а иногда даже окраинных районах, как Индонезия (Ниа на Калимантане), Западная Европа (Кроманьон, Ханёферзанд) или Южная Африка (Флорисбад). Один из самых существенных вопросов касается древности формирования рас современного человечества — негроидной, европеоидной и монголоидной <...>
Сами же большие расы современного человечества образовались только в постпалеолитическое время. То есть сапиенс древнее рас, процессы сапиентации и расогенеза совпадали лишь частично, и последние происходили уже на достаточно смешанной основе.
Заключительный этап гоминизации — процесс сапиентации — занял в основном последние 100 тыс. лет. На этом отрезке антропогенеза произошли существенные сдвиги в морфологической организации, познавательных способностях, снизилась скорость процессов старения, увеличилась продолжительность жизни (с. 99—101).
7. Индивидуальное развитие человека
Индивидуальное развитие человека, как и всякого другого организма, есть онтогенез с заложенной в нем филогенетической программой, его периодизация неизбежно покоится на выделении ряда универсальных возрастных процессов (рост, созревание, развитие, старение), в ходе которых формируются соответствующие возрастные свойства (различия). То и другое обобщается в понятии возрастных стадий (фаз, этапов, периодов) или этапов развития (детство, переходный возраст, зрелость, старость и др.). Возрастные процессы подразумевают вопрос, как формируются возрастные свойства и каким путем (постепенно или резко, скачкообразно) происходит переход из одной возрастной стадии в другую.
В современной науке существует три главных термина для описания индивидуального развития в его целом – время жизни, жизненный цикл, и жизненный путь. Хотя их не редко используют как синонимы, они существенно различны по содержанию. Время жизни, ее протяженность или пространство, обозначает временный интервал между рождением и смертью. Продолжительность жизни имеет важные социальные и психологические последствия. От нее во многом зависит, например, длительность существования поколений и продолжительность первичной социализации детей. Понятие жизненный цикл более определенно и содержательно. Оно предполагает, что ход жизни подчинен известной закономерности, а его этапы ("возрасты жизни", или "времена жизни", подобные временам года) представляют собой постоянный круговорот. Идея циклического круговорота жизни, подобного цикличности природных процессов (чередование дня и ночи, смена времен года и т.д.). Многие биологические и социальные возрастные процессы действительно являются циклическими. Организм нормально проходит фазы рождения, роста, созревания, старения и смерти. Личность усваивает, затем выполняет и, наконец, постепенно оставляет определенный набор социальных ролей (трудовых, семейных, родительских), после чего тот же цикл повторяют ее потомки. Цикличность характеризует и смену поколений в обществе, где младшие (дети) сначала учатся у старших, затем активно действуют рядом с ними, а потом, в свою очередь, социализируют младших. Понятие жизненного пути отличается от "жизненного цикла" прежде всего многомерностью, тем, что оно предполагает множество разных тенденций и линий развития в пределах одной и той же биографии, причем эти линии одновременно автономны и взаимосвязаны. В основе его периодизации – нелинейные, раз и навсегда определенные фазы, а конкретные жизненные события. Время, последовательность и способ осуществления любого жизненного события, будь то вступление в брак или выход на пенсию, не менее важны, чем сам факт, что данное событие имело место. Это требует сочетания социологического, психологического и исторического анализа. Таким образом, содержательная характеристика процессов, свойств и стадий индивидуального развития возможно либо в системе онтогенеза, либо в системе жизненного цикла, либо в системе жизненного пути. Однако эти системы не рядоположны: жизненный путь личности включает в себя жизненный цикл индивида, а этот в свою очередь включает онтогенез.
Рост развития человеческого организма
На процессы роста и развития оказывает влияние большое число самых разнообразных эндо - и экзогенных факторов. "Рост" и "развитие" обычно употребляются как понятия тождественные, неразрывно связанные между собой. Между тем, биологическая природа этих процессов различна, отличаются их механизмы и последствия.
Рост - это количественное увеличение биомассы организма за счет геометрических размеров и массы отдельных клеток.
Развитие — это качественные преобразования в многоклеточном организме за счет дифференцированных процессов (увеличения разнообразия многоклеточных структур), которые приводят к качественным и количественным изменениям функций организма.
Процесс роста и развития характеризуется следующими закономерностями:
1. Дифференциация и интеграция частей и функций, автономиза-ция развития, возрастающая в ходе филоонтогенеза.
2. Диалектическое единство непрерывного и прерывистого; постепенность и цикличность. На кривой роста человека можно отметить три основных цикла в постнатальном развитии:
а) от рождения до 10—13 лет при постоянном снижении скорости;
б) пубертатный спурт;
в) падение скорости ниже уровня до пубертатного периода и прекращение роста.
Сочетание периодов ускоренного развития и относительной стабилизации можно выявить даже на коротких отрезках онтогенеза. Например, при долговременном наблюдении с недельными интервалами детей от рождения до 2-х лет. Нелинейность роста (мини-скачки) наблюдается примерно у 70 % детей и подростков при еженедельном обследовании в течение 6-10 месяцев.
3. Гетерохрония (разновременность) в созревании разных систем организма (или тканей) и разных признаков в пределах одной системы. Согласно концепции системогенеза П.К.Анохина определяющими темпами созревают жизненно важные функции, обеспечивающие первоочередное формирование комплексных адаптивных реакций, специфических для каждого конкретного этапа взаимоотношений организма с внешней средой.
4. Значительное индивидуальное разнообразие возрастной динамики по отдельным этапам онтогенеза, зависящее от генетической программы.
Тема 11. Периодизация индивидуального развития (онтогенеза)
Онтогенез - комплекс последовательных преобразований организма, начиная со стадии оплодотворения яйцеклетки и до окончания жизненного цикла.
Пифагор (VI в. до н.э.) выделял 4 периода человеческой жизни: весну (до 20 лет), лето (20-40 лет), осень (40-60 лет) и зиму (60-80 лет), которые соответствовали становлению, молодости, расцвету сил и угасанию.
Гиппократ разделил весь постнатальный онтогенез на десять семилетних циклов.
Онтогенез подразделяется на 2 периода: эмбриональный и фетальный (плодный). В течение первого периода, который продолжается 8 недель, происходит формирование органов и частей тела, свойственных, взрослому человеку. В фетальный период увеличиваются размеры и завершается органообразование. Скорость роста плода возрастает до 4-5 месяцев. После 6 месяцев скорость роста линейных размеров уменьшается.
На VII Всесоюзной конференции по возрастной морфологии, физиологии и биохимии, состоявшейся в 1965 г. в Москве была принята следующая схема возрастной периодизации онтогенеза человека. Эта схема нашла широкое применение в антропологии, педиатрии, педагогике.
Схема возрастной периодизации онтогенеза человека
| Новорожденные |
1-10 дней |
| Грудной возраст |
10 дней—1 год |
| Раннее детство |
1-3 года |
| Первое детство |
4-7 лет |
| Второе детство |
8-12 лет (мальчики) 8-11 лет (девочки) |
| Подростковый возраст |
13-16 лет (мальчики) 12-15 лет (девочки) |
| Юношеский возраст |
17-21 лет (юноши) 16—20 лет (девушки) |
| Зрелый возраст (1 период) |
22-35 лет (мужчины) 21-35 лет (женщины) |
| Зрелый возраст (2 период) |
36-60 лет (мужчины) 36-55 лет (женщины) |
| Пожилой возраст |
61-74 лет (мужчины) 56-74 лет (женщины) |
| Старческий возраст |
75-90 лет |
| Долгожители |
90 лет и выше |
С момента рождения наступает период новорожденности. В это время ребенок вскармливается молозивом в течение 8-10 дней. Следующий период - грудной - продолжается до 1 года. Начало его связано с переходом к питанию "зрелым молоком". Длина тела увеличивается от рождения до года примерно в 1,5 раза, а вес утраивается. С 6 месяцев начинают прорезываться молочные зубы, и на 2-3 году жизни заканчивается прорезывание молочных зубов. В период первого детства, начиная с 6 лет, появляются первые постоянные зубы. В период второго детства выявляются половые различия в размерах и форме тела, а также начинается усиленный рост в длину. В среднем к 12—13 годам у мальчиков и девочек заканчивается смена зубов. Начинают развиваться вторичные половые признаки. Следующий период - подростковый — называют периодом полового созревания, или пубертатным, к концу которого размеры тела составляют 90-97 % своей окончательной величины, а основные функциональные характеристики подростков приближаются к характеристикам взрослого организма. В период юношеского возраста заканчивается процесс роста и формирование организма, все основные размерные признаки достигают дефинитивной (окончательной) величины. В зрелом возрасте форма и строение тела изменяются мало. Между 30-ю и 45-50-ю годами длина тела остается постоянной, а потом начинает уменьшаться. В пожилом и старческом возрасте происходят инволютивные изменения организма.
Тема 12. Пубертатный скачок роста
Важным событием пубертатного периода является спурт - скачкообразное увеличение роста, наблюдающееся у мальчиков в среднем в 13-15 лет, а у девочек - в 11-13 лет. В течение этого времени абсолютная скорость роста непостоянна и постепенно снижается: у мальчиков - от 12 до 7 см / год, у девочек - от 11 до 6 см / год.
В осуществлении ростового спурта задействованы многие факторы, и в первую очередь половые гормоны и гормон роста - СТГ, оказывающий свое воздействие посредством соматомединов - пептидов, синтезирующихся в печени. Помимо этих гормонов, существенное значение в данном периоде имеет универсальный анаболический гормон инсулин и гормоны щитовидной железы - тироксин и трийодтиронин. Инсулин регулирует белковый и жировой обмен, а тироксин и трийодтиронин влияют на рост и дифференцировку тканей, повышение интенсивности основного обмена и теплопродукции. Созревание репродуктивной функции завершается к 18—20 годам.
Морфологическое созревание лобной корки достигается только к 12 годам, а окончательное формирование полушарий конечного мозга - в 20-22 года. Для пубертатного периода характерными считаются: усиление подкорковых влияний и ослабление деятельности коры головного мозга, нарушение вегетативной сферы, повышенная эмоциональность, особенно у девочек.
8. Конституции человека
Конституция — это совокупность функциональных и морфологических особенностей организма, сложившихся на основе наследственных и приобретенных свойств, которые определяют своеобразие реакции организма на внешние и внутренние раздражители.
Конституция человека, в отличие от его физического развития, это характеристика конкретного человека, которая остается постоянной на протяжении всей его жизни, то есть она не имеет возрастной периодизации. По сути, конституция человека — это генетический потенциал человека, продукт наследственности и среды, реализующей наследственный потенциал.
Из факторов внешней среды, под влиянием которых реализуются особенности конституции (социально-экономические условия, питание, перенесенные болезни, занятия физической культурой и спортом), особенно в детском и подростковом возрасте.
Конституция, как обобщенная морфофункциональная характеристика индивидуума, отражает особенности не только телосложения, но также психической деятельности, метаболизма и функционирования вегетативных систем, адаптационных, компенсаторных и патологических реакций человека. Хотя проблема конституции имеет многовековую историю, до настоящего времени нет общепринятой формулировки этого понятия, пригодной для лиц различного пола и возраста. Различные методические подходы к выделению конституциональных типов (соматоскопия, антропометрия и их комбинация), неоднозначное толкование самого понятия "конституция" — все это привело к созданию многочисленных схем конституциональной диагностики. Длительное время конституционные типы изучали в связи с предрасположенностью к определенным заболеваниям, затем их стали рассматривать как нормальные варианты строения и функционирования организма. Если рассматривать весь спектр жизненных доминант и установок, то, в конечном счете, они детерминированы нашей конституцией: потребности, способности, интересы, желания, искушения, проблемы алкоголизма, курения, наркомании имеют генетическую компоненту при всей значимости среды и воспитания в этих вопросах. Все проблемы предрасположенности (не предрасположенности) к болезням — тоже конституционально детерминированы, также как и личные предпочтения в образе жизни, духовные установки, психический мир знаний, эмоций и воли, поведение, любовь и ненависть, сексуальный потенциал.
Генотипически астеник никогда не станет гиперстеником и, наоборот. Среда лишь модифицирует наш конституционный потенциал в рамках: конституция — генетически устойчива, стабильна, а фенотип — лишь модифицируется, изменяет конституцию в пределах геномных законов реагирования.
Реальная конституция человека складывается из набора составляющих:
· Рефлексивная конституция — генетическая память (эмбриональный путь развития), иммунная память (о перенесенных заболеваниях), нейронная память (память, фиксируемая нейронами).
· Генотипическая конституция — это исходный "наследственный паспорт" (геномная характеристика, хромосомная система человека), определяющий регенерационные способности нашей морфологии (например, заживление ран).
· Фенотипическая конституция — это традиционное (а подчас единственное) представление о конституции человека, на основе наследственной структуры его костно-мышечного "портрета" (по современной терминологии — это эктоморфы, мезоморфы, эндоморфы, различные типы астенических, нормостенических и гиперстенических конституций). По мнению В. П. Петленко выделяют 5 конституциональных типов (варианты нормы): 1) нормостенический, 2) гиперстенический, 3) гипостенический (астенический) 4) грациальный (маленький, пропорциональный, изящный) 5) атлетический тип. Чаще всего эти типы вариабельны, то есть чистый тип выделить на практике удается редко.
· Соматическая конституция — особенности телосложения. Соматотип — тип телосложения — определяемый на основании антропометрических измерений (соматотипировании), генотипически обусловленный, конституционный тип, характеризующийся уровнем и особенностью обмена веществ (преимущественным развитием мышечной, жировой или костной ткани), склонностью к определенным заболеваниям, а также психофизиологическими отличиями.
· Иммунная конституция — система глобулиновой защиты, в основе которой лежит механизм "антиген-антитело", определяющих характер и интенсивность иммунологических реакций ("нормального" или аллергического, патологического характера), обеспечивающих постоянство внутренней среды (гомеостаз), либо ее патологию.
· Нейронная конституция — состояние неврологической реактивности, определяющее базисные основы обучения, приобретения знаний; это — базисная основа эмоций и волевых процессов, определяющих наше здоровье и нашу неврологическую симптоматику.
· Психологическая конституция определяет психологический тип личности, ее характер и темперамент.
· Лимфогематологическая конституция — это особенности лимфотока и группа крови, носящие геномный, конституциональный характер и определяющие интенсивность метаболизма и энергетики организма.
· Гормонально-половая конституция характеризует существенный момент во всех поведенческих реакциях человека. Половое поведение определяется взаимодействием определенных мозговых структур с половыми гормонами (андрогенами при мужском половом поведении и эстрогенами — при женском половом поведении).
Люди отличаются друг от друга как по морфологическим и функциональным, так и по психическим особенностям. Различны они и по характеру реакции на патогенные раздражители. Гален (ок. 130 - ок. 200 гг. н.э.) ввел понятие о габитусе, то есть о совокупности признаков и особенностей строения тела, характеризующих внешний облик человека.
Constitucio в переводе с латыни означает устройство. Впервые с понятием конституции мы встречаемся у Гиппократа (460-377 гг. до н.э.). Он различал конституцию хорошую и плохую, сильную и слабую, жировую, влажную, потную и т.д. По Эмпедоклу, в основе мира лежит 4 элемента: огонь, вода, воздух и земля. Согласно Гиппократу, тело человека состоит из 4-х элементов: огонь — теплая кровь; воздух - холодная селезеночная жидкость; земля - сухая, горькая желчь печени; вода - холодная и влажная слизь, вытекающая из мозга. Пока все 4 свойства уравновешивают друг друга, человек здоров, нарушилось равновесие — наступает болезнь. Люди разных конституций склонны к разным болезням.
Классификация конституции человека по Сиго
Сиго и его ученики Шайю и Мак-Олаф считали, что конституционные типы есть результат воздействия окружающей среды, и разделили всех людей на 4 типа:
1. Церебральный (мозговой)
2. Респираторный (дыхательный)
3. Дегустативный (пищеварительный)
4. Мускулярный (мышечный)
Классификация конституции человека по Э.Кречмеру
Согласно Кречмеру, существует связь между строением тела и психическими особенностями человека. По его мнению, понятие конституции является психофизическим, общебиологическим и относится как к телесному, так и к психическому. Он отрицал влияние среды на конституцию человека и выделил 3 типа:
1. Астенический
2. Пикнический
3. Атлетический
Особенности пикнического сложения:
1) большие размеры внутренних полостей (голова, живот, грудь);
2) склонность к образованию жировых отложений.
Классификация конституции человека по В.М.Богомольцу
В.М.Богомолец в основу классификации положил строение соединительной ткани человека и выделил 4 типа:
1. Астенический (преобладает тонкая, нежная соединительная ткань)
2. Фиброзный (преобладает плотная волокнистая соединительная ткань)
3. Пастозный (преобладает рыхлая соединительная ткань)
4. Липоматозный (преобладает жировая ткань)
Классификация конституции человека по У.Шелдону
Ее основу составлют три типа (компонента): эндоморфия, мезо-морфия и эктоморфия, приблизительно соответствующие крайним вариантам пикника (преимущественное развитие жирового компонента), атлета (преобладание костно-мускульного компонента) и астеника (отсутствие этих тенденций). Степень выраженности каждого типа оценивается по 7-балльной шкале на основе фотографического описания, 17-ти измерительных признаков и весоростового индекса. Например, формула соматотипа, по У.Шелдону, 3-5-2, где приводятся средние оценки соответственно для компонентов эндо-, мезо- и эктоморфии.
I тип - эндоморфный;
II тип - эктоморфный;
III тип - мезоморфный.
Классификация конституции человека по М.В.Черноруцкому. Индекс Пинье
М.В.Черноруцкий выделил 3 типа конституции человека:
1) астенический;
2) нормостенический;
3) гиперстенический.
Для того чтобы определить, какой тип конституции у данного человека, нужно определить индекс Пинье по формуле L - ( Р + Т ), где L -длина тела, Р - вес тела в кг, Т - окружность грудной клетки в см.
Если индекс Пинье больше 30 - астенический тип сложения, меньше 10 - гиперстенический. От 10 до 30 - нормостенический.
Конституцию можно определить как основную биологическую характеристику целостного организма человека, проявляющуюся в особенностях его реактивности и резистентно (устойчивости), в основе которых лежит специфическая норма реакций индивидуального генотипа.
9. Эволюция мозга
Существовали две точки зрения на эволюцию мозга
гоминид. Одни исследователи считали главным показателем развития размеры мозга, его объем. Другие авторы придавали большее значение структурным качественным преобразованиям коры.
Сторонники первой точки зрения считали основным критерием выделения семейства гоминид размеры мозга. В связи с этим существовала гипотеза "мозгового рубикона", выдвинутая английским антропологом А.Кизсом. Суть ее состоит в следующем - по объему мозга современные и ископаемые гоминиды оказывались как бы на разных берегах символической реки Рубикон. На одном берегу -
австралопитековые, объем мозга, которых не превышает 700 куб. см, на другом берегу - все ископаемые и современные люди с мозгом не менее 850 куб.см. А.Кизс предположил, что существует "критическая масса" мозга, без которой невозможно изготовление орудий и другие сложные формы поведения. Такая масса составляет, по его мнению, 750 куб.см. Иными словами, если объем мозга равен 700 куб.см, то это еще не человек, а если 755 куб.см - то уже человек.
Как известно, объем головного мозга как таксономический признак имеет малую ценность. Его величина вариабельна даже в пределах одного вида. Его показатели могут перекрываться у нескольких видов. Поэтому сторонники второй точки зрения главным морфологическим критерием выделения семейства гоминид считали систему прямохождения. Увеличение размеров мозга, безусловно, характерно для приматов. Однако большее значение в эволюции мозга гоминид имели качественные перестройки коры, появление у гоминид новых специфических человеческих областей коры головного мозга.
Можно заключить, что в эволюции головного мозга гоминид сочетались увеличение размеров мозга и перестройка его отдельных участков с увеличением зон абстрактного мышления и уменьшением зон чувственного восприятия. За период от 4 млн лет до 10 тыс. лет назад мозг вырос от 500 до 1500 куб.см (в среднем), т.е. в 3 раза. Причем на более поздних этапах антропогенеза развитие мозга опережало развитие зубной системы и локомоторного аппарата. Еще большие изменения происходили в микроструктуре мозга.
В.И.Кочеткова связывает изменения, происходившие в
макро- и микроструктуре мозга гоминид, с их деятельностью.
Существует мнение, что укрупнение головного мозга у млекопитающих сопровождалось увеличением размеров тела. И.Эйсенберг назвал эту связь "энцефализацией". Вес головного мозга связан с весом тела. Абсолютный вес мозга больше у крупных животных, чем у мелких. Относительный вес мозга - вес мозга/вес тела у крупных животных в среднем меньше, чем у мелких.
Я.Я.Рогинский (1978) ввел индекс E2/S, где Е - вес мозга, S - вес тела. Этот индекс позволяет сравнивать животных по степени их церебрализации более объективно. У животных, имеющих аналог руки, - паукообразная обезьяна с осязательным хватательным хвостом (8,64), слон с хоботом (9,62) и др., - этот индекс имеет наибольшие указатели среди млекопитающих. У человека этот индекс равен 32.
Р. Фоули полагает, что укрупнение головного мозга у гоминид является следствием питания мясом. Причем основное влияние на увеличение размеров головного мозга оказывало изменение стратегий поиска пищи. Разнообразие пищевого рациона, расширение охотничьих территорий было взаимосвязано с усложнением социальных связей, усилением кооперации и заботы о молодых, об их обучении. В свою очередь, развитие социальности и средств коммуникации также вело к развитию мозга.
Помимо экологических и поведенческих существуют гипотезы, объясняющие увеличение мозга генетическими факторами.
Гипотеза мутации. Считают, что такие явления, свойственные человеку, как мясоедение, гиперсексуальность и проявление агрессивности, могли возникнуть как мутация. Причиной последней могла быть жизнь в условиях повышенной радиации.
Гипотеза изоляции - гетерозиса. Ускоренную эволюцию мозга в эпоху
плейстоцена связывают с демографическими изменениями популяции древнейших гоминид. Речь идет о крупных популяциях, подразделенных на много мелких домов, почти не скрещивающихся между собой. Предполагают, что при скрещивании ранее изолированных групп возникал новый стимул развития головного мозга в виде эффекта гетерозиса. Гетерозис - это увеличение размеров тела и внутренних органов вследствие скрещивания отдаленных групп (
Никитюк, Чтецов, 1983 ).
10. Социальные факторы происхождения человека
К социальным факторам антропогенеза относят труд, общественный образ жизни, развитое сознание и речь. Роль социальных факторов в антропогенезе была раскрыта ф.Энгельсом в работе "Роль труда в процессе превращения обезьяны в человека" (1896). Этим факторам принадлежала ведущая роль на более поздних этапах становления человека.
Важнейший фактор эволюции человека - труд. Способность изготавливать орудия труда свойственна только человеку. Животные могут лишь использовать отдельные предметы для добывания пищи (например, обезьяна использует палку, чтобы достать лакомство).
Трудовая деятельность способствовала закреплению морфологических и физиологических изменений у предков человека, которые называют антропоморфозами.
Важным антропоморфозом в эволюции человека было прямохождение. В течение многих поколений в результате естественного отбора сохранялись особи с наследственными изменениями, способствующими прямохождению. Постепенно сформировались приспособления к прямохождению: S-образный позвоночник, сводчатая стопа, широкие таз и грудная клетка, массивные кости нижних конечностей.
Прямохождение привело к высвобождению руки. Сначала рука могла выполнять лишь примитивные движения. В процессе труда она совершенствовалась, стала выполнять сложные действия. Таким образом, рука является не только органом труда, но и его продуктом. Развитая рука позволила человеку изготавливать примитивные орудия труда. Это дало ему значительные преимущества в борьбе за существование.
Совместная трудовая деятельность способствовала сплочению членов коллектива, вызывала необходимость обмена звуковыми сигналами. Общение способствовало развитию второй сигнальной системы - общению при помощи слов. Сначала наши предки обменивались жестами, отдельными нечленораздельными звуками. В результате мутаций и естественного отбора шло преобразование ротового аппарата и гортани, формирование речи.
Труд и речь влияли на развитие мозга, мышление. Так в течение длительного времени в результате взаимодействия биологических и социальных факторов осуществлялась эволюция человека.
Если морфологические и физиологические особенности человека передаются по наследству, то способность к трудовой деятельности, речь и мышление развиваются только в процессе воспитания и образования. Поэтому при длительной изоляции ребенка у него не развиваются совсем или развиваются очень плохо речь, мышление, приспособленность к жизни в обществе.
11. Строение тела и характер
Строение тела и характер Э. Кречмер
(Общий взгляд на систему) …Мы установили три постоянно повторяющихся главных типа строения тела, которые мы назвали астеническим, атлетическим и пикническим. Эти типы встречаются у мужчин и женщин, но при более слабой морфологической дифференцировке женского тела, яркие картины у женщин бывают гораздо реже. Способ же распределения этих типов в шизофренической и циркулярной группах весьма различен и очень любопытен. В здоровой жизни мы всюду встречаем эти же три типа, сами по себе они не содержат ничего болезненного, но свидетельствуют об определенных нормально-биологических предрасположенностях, из которых лишь незначительная часть достигла патологического завершения, как в психиатрической области, так и в определенных внутренних заболеваниях. Наряду с этими большими главными типами мы нашли затем различные небольшие группы, которые объединяем под общим названием диспластических специальных типов, поскольку они представляют собой сильные отклонения от среднего типа и обнаруживают тесную морфологическую связь с дисгландулярными синдромами патологии желез внутренней секреции… …Мы еще раз подчеркиваем, что морфологию строения тела надо сначала изучать на мужчинах, а затем уже на женщинах. Строение женского тела (в чертах лица, развитии мышц и жира) в общем менее рельефно, поэтому женщины чаще демонстрируют типичные и менее резкие проявления.
(Астенический тип у мужчин) Понятие астенического габитуса введено Штиллером, …и ограничено в желательном смысле Бауэром. Полученный на нашем психиатрическом материале typus astenicus совпадает с таковым же типом Бауэра. Из отдельных черт, упомянутых в краткой работе Бауэра, большинство подтверждается и нашими исследованиями. Но мы не можем согласиться с утверждением, что астеники преимущественно долихоцефалы, по нашим данным, череп астеников по сравнению с другими типами небольшого объема, средней ширины, короткий и низкий… …Короче говоря, габитус мужчины-астеника характеризуется главным образом слабым ростом по толщине при среднем не уменьшенном росте в длину. Это недостаточное развитие толщины проходит через все части тела - лицо, шею, туловище, кости, сосудистую систему - и через все формы тканей, кожу, жировую ткань, мышцы, кости, сосудистую систему. Вследствие этого средний вес, а также размеры в объеме и ширине понижены по сравнению со средними размерами, характерными для мужчин (см. табл. 1.)
Таблица 1Основные размеры тела у людей астенического типа (в среднем)
| Признаки |
Мужчины |
Женщины |
| Вес (кг) |
50.5 |
44.4 |
| Длина тела (см) |
168.4 |
153.8 |
| Ширина плеч (см) |
35.5 |
32.8 |
| Объем груди (см) |
84.1 |
77.7 |
| Объем живота (см) |
74.1 |
67.7 |
| Объем в области бедер (см) |
84.7 |
82.2 |
| Объем предплечья (см) |
23.5 |
20.2 |
| Объем кисти (см) |
19.7 |
18.0 |
| Объем икр (см) |
30.0 |
27.7 |
| Длина ног (см) |
89.4 |
79.2 |
Следовательно, в тяжелых случаях мы имеем картину худого, тонкого человека, кажущегося выше, чем он на самом деле, малокровного, с узкими плечами, с сухими тонкомышечными руками, с тонкокостными кистями, с длинной, узкой, плоской грудной клеткой, на которой можно пересчитать ребра, с острым реберным углом, с тонким, лишенным жира животом, с такими же руками и ногами (см. рис. 1). У мужчин наблюдается явное отставание веса тела. Представители этого типа имеют более широкие плечи при плоской, как доска, грудной клетке и очень нежных плечевых костях. Вместо тонкого живота в отдельных случаях наблюдается дряблый, энтероптотичный отвисший живот или отложения жира евнухоидного или женского распределения, что нельзя смешивать с пикническим жирным животом. Часто возникают варианты астенического типа с более сильным или слабым выражением стигмат телосложения дисгенитальной группы, инфантилизма (акромикрия), феминизма (образование талии, увеличенный объем таза, резкая округленность бедер) и особенно евнухоидного высокого роста с чрезмерной длиной конечностей… Нередко образуются варианты и комбинации астенического и атлетического типов, когда наряду с астеническими стигматами выступают и атлетические (например, длинная, узкая грудная клетка с крепкими конечностями, инконгруэнция между лицом и строением тела и т.д.) или средний тип жилисто-стройной фигуры, для которого характерна грациозная худощавость или крепкие мышцы. Если мы рассмотрим тип в течение развития его жизни, то заметим, что во всех возрастных периодах он остается довольно постоянным. В детском возрасте эти люди бывают слабыми и нежными; в период созревания они быстро растут и вытягиваются, в зрелости и старости они не обнаруживают ни малейшей склонности к правильному накоплению жира и развитию мышц. Они, как крестьяне, могут выполнять тяжелую физическую работу, но у них очень слабо утолщаются мышцы. Они могут прекрасно питаться и даже, как многие обитатели больниц в мирное время, отличаться прожорливостью, но, несмотря на это, оставаться такими же худыми. Возраст изменяет у части астеников тип распределения волос. Черты лица лишь на восемнадцатом году приобретают свои характерные формы, которые будут позже описаны, и могут при прогрессирующем похудании в пожилом возрасте сделаться еще резче. У части астеников бросается в глаза как важный биологический признак их преждевременное старение. Я встречал мужчин в возрасте между 35 и 40 годами, казавшихся стариками с морщинистой, совершенно сухой, дряблой и поблекшей кожей и с перевитыми височными артериями. В таких случаях, несмотря на нормальный образ жизни пациентов, наблюдается общая атрофия жира и мышц, свидетельствующая о тяжелом хроническом нарушении обмена веществ. Этим страдают даже те пациенты, которые постоянно много едят… мышцы их, могут даже при постоянной работе достичь таких степеней атрофии…, что появляется диастаз абдукторов бедер… …Что же касается астенического строения тела у мужчин в период полового созревания, то в описываемых случаях рост в длину происходил нормально, иногда бывал очень сильным и преждевременным. Шестнадцатилетний гебефренический юноша имел рост 165 см (что у нашего швабского населения значительно превосходит средние размеры), 18-летний был ростом 176 см. Но это нельзя считать правилом. Наименьший из наших взрослых мужчин-астеников имел рост 158 см, самый большой - 178 см, однако низкорослые среди них встречаются не часто. Только что упомянутый был единственным среди 50 мужчин, чей рост меньше 160 см. Резкие детские, неразвитые черты лица встречаются у отдельных астеников в возрасте от 20 до 25 лет, особенно у женщин; многие астеники при худощавом лице с резко выступающими чертами уже в этом возрасте выглядят старше своих лет. О сексуальном инстинкте и волосяном покрове мы скажем позже.
(Астенический тип у женщин) Астенические женщины по своему габитусу напоминают астенических мужчин, за исключением одного: они не только худощавы, но часто и малорослы. Нормальный и даже усиленный рост в длину у них встречается, но не так часто. Среди 20 женщин я встретил одну ростом 169 и одну ростом 161 см, остальные имели рост 160, а самая маленькая - 145 см. Эта группа женщин не просто астенична, а астенично-гипопластична, причем во всей нашей работе под астеническим мы понимаем задержанный рост в толщину, под гипопластическим - общее недоразвитие тела и его частей, особенно роста в длину. Так, у астенических женщин самый низкий средний рост 153,8 см, между тем как у астенических мужчин он равен 168,4 см, что соответствует среднему росту швабского населения мужского пола. Несоответствие между ростом и весом тела у астенических мужчин выражено более резко, чем у женщин. У мужчин оно равно примерно 18, у женщин - 9 (см. табл. 1).
(Атлетический тип у мужчин) Мужской атлетический тип характеризуется сильным развитием скелета, мускулатуры, кожи (см. рис. 2). Общий облик самых ярких представителей этого рода: среднего или высокого роста мужчины с особенно широкими и выступающими плечами, статной грудной клеткой, упругим животом, с формой туловища, сужающегося книзу, так что таз и все еще статные ноги по сравнению с верхними частями тела и особенно гипертрофичным плечевым поясом кажутся иногда тонкими. Плотная высокая голова прямо держится на свободной шее, причем линейные контуры trapezius , видимые спереди, придают шейной части плеча особый отпечаток. Линии, очерчивающие тело, образуются выступающими мышцами хорошей или гипертрофической мускулатуры, которая вырисовывается пластически, как мускульный рельеф. Костный рельеф наиболее заметен в чертах лица. Грубое строение костей характерно для ключицы, запястья и плюсны. Наряду с плечевым поясом трофический акцент лежит еще на концах конечностей, которые могут напоминать акромегалию. Наибольший объем руки у атлетиков достигает 25 см, величины, превышающей на 5 см средние размеры, сравнительно умерен, почти нормально выражен. Этим прежде всего обусловливается точный мускульный рельеф, причем чрезмерно развитая мускулатура заметно проступает через тонкий жировой слой у мужчин Объем руки в 23 см довольно част. Кроме объема руки, при этом типе любопытна и ширина плеч, которая в двух случаях достигает наибольшей цифры в 42,5 см, превышающей среднюю величину ширины плеч нашего населения (37,5-38 см) на 5 см. Длина конечностей скорее велика, чем мала. Наряду с костями и мышцами в гипертрофии участвует и кожа. Она обладает хорошим упруго-эластичным тургором, а на лице кожа особенно плотная, толстая… Рост тела в общем выше среднего, длина больше 180 см встречается нередко - самый большой атлетик в нашем исследовании имел рост 186 см. Нижнюю границу фиксировать невозможно, так как морфологические переходы между атлетическим типом и типом гипопластическои плечистости довольно неясны. Любопытны переходы к формам великанов (см. табл. 2)
Таблица 2Основные размеры тела у людей атлетического типа (в среднем)
| Признаки |
Мужчины |
Женщины |
| Вес (кг) |
62.9 |
61.7 |
| Длина тела (см) |
170.0 |
163.1 |
| Ширина плеч (см) |
39.1 |
37.7 |
| Объем груди (см) |
91.7 |
86.0 |
| Объем живота (см) |
79.6 |
75.1 |
| Объем в области бедер (см) |
91.5 |
95.0 |
| Объем предплечья (см) |
26.2 |
24.2 |
| Объем кисти (см) |
21.7 |
20.0 |
| Объем икр (см) |
33.1 |
31.7 |
| Длина ног (см) |
90.9 |
85.0 |
В остальном у представителей атлетического типа, главным образом в чертах лица, наблюдаются варианты. В строении тела мы находим наряду с только что намеченным типом, обладающим сравнительно стройной нижней частью тела и пластическим выступанием форм тела, еще вариант, который отличается общей неуклюжестью. Различие в развитии плеч и таза при этом не заслуживает внимания, все некрасиво, грубо, неуклюже, кожа лица пастозна, мускульный рельеф затушевывается диффузным развитием жира. Чисто морфологически нельзя решить, имеет ли здесь место вариант, комбинация, другая стадия развития или, что вероятнее, особенный тип с существенным отклонением биологической основы. В моем исследовании есть лишь несколько случаев такого рода, один из пациентов, наряду с тяжелым кататоническим слабоумием, страдал эпилептическими припадками. У больного с инфантильным развитием жира наблюдались слаборазвитые половые органы и двусторонний крипторхизм. Среди неуклюжих атлетиков я встречал в больнице и таких, у которых громадная верхняя часть туловища сидела не просто на стройной, а крайне тонкой его нижней части, на инфантильно узком тазе и астенически тонких ногах. Также многочисленны морфологические взаимоотношения между атлетическим типом и дисгенитальной группой. Комбинации атлетических элементов с евнухоидной великорослостью не представляют собой необычного явления, например, чрезмерная длина конечности с большими кистями, частичная плотность костей, мускулистый плечевой пояс и т.д. Наблюдается также резкий феминизм. Недавно я видел шизоидного психопата с атлетическим строением тела, но с узкими опущенными округленными плечами, с талией и женским тазом большого объема… … Исследование развития атлетического типа в зависимости от возраста дает мало интересного. Этот тип ясно выступает уже в периоде созревания приблизительно с 18 лет; с полным развитием головы после 25 лет он становится более пластичным и четким. Мне удалось установить несколько случаев атлетиков в 50-летнем возрасте. В более позднем возрасте, на шестом десятке, я не имею в своем распоряжении ни одного испытуемого, что объясняется случайностью, так как атлетическая группа гораздо меньше астенической и охватывает лишь около 30 случаев, считая мужчин и женщин вместе.
(Атлетический тип у женщин) Атлетический тип у женщин, поскольку его здесь можно распознать, такой же, что и у мужчин, но с известными характерными отличиями. Развитие жира у атлетических женщин не задержано, а обильно: конечно, оно гармонирует с остальными тканями, прежде всего с костями и мышцами, и, по крайней мере, в наблюдаемых нами случаях не элективно усилено, как у пикников. Наряду с этим типом атлетических женщин с женственно-округленными формами мы встречаем, однако, и пациенток с выраженным маскулинизмом в чертах лица и строении тела. Несколько таких случаев, по мускульному рельефу и пропорциям плеч и таза точно напоминают мужчин. Вообще следует иметь в виду, что трофическое акцентирование плечевого пояса можно часто наблюдать и у атлетических женщин (до 39 см ширины плеч), причем оно не идет по линии вторичных половых признаков, а скорее противоречит ей. Последнее указывает на то, что такое строение плечевого пояса представляет собой не что-то случайное, а вытекает из своеобразно направленных импульсов роста. Кроме грудино-плечевого пояса часто сильно развит и таз.
(Нормальность и анормальность строения. Субъективизм оценок) Строение тела атлетических женщин производит впечатление ненормального, чрезмерно выступающего, грубоватого и массивного в отличие от строения тела у атлетических мужчин. Это происходит потому, что мужчины-атлетики иногда приближаются к нашему художественному идеалу красоты, в то время как наш идеал женской красоты не согласуется с атлетическим. Здесь уместно предостеречь от внесения субъективных оценок в диагностику строения тела. Мы мало чего достигнем характеристикой "нормальный" и "анормальный" по отношению к нашим трем главным типам. Все три встречаются как у здоровых, так и у душевнобольных и больных с внутренними болезнями. Нельзя также утверждать, что один из трех типов в физическом отношении лучше приспособлен к жизненной борьбе. Атлетику легче быть борцом, астенику - летчиком, при многих профессиях совершенно безразлично, посвящает ли себя им тот или другой. Многие астеники в течение всей жизни здоровы и доживают до глубокой старости, тогда как их более статные ровесники давно умерли от сердечного удара. Поэтому неправильно (и здесь я совершенно согласен с Бауэром) говорить о morbus asthenicus. И определение "артритический габитус" или "физический габитус" являются узкими клиническими терминам. Для совершенно объективного натуралистического учения о строении тела они как крайне односторонне устанавливающие тенденцию к определенному заболеванию не могут быть использованы. Если астеник очень предрасположен к туберкулезу, то он более иммунен к ревматизму, диабету, артериосклерозу, чем пикник, и наоборот. В отличие от психиатрических, о предрасположенности атлетиков к внутренним заболеваниям мы ничего определенного не могли найти. Поэтому ни одну из больших конституциональных групп нельзя признать ни более здоровой, ни более больной.
(Пикнический тип у мужчин) Пикнический тип в среднем возрасте характеризуется сильным развитием внутренних полостей тела (головы, груди, живота) и склонностью торса к ожирению при нежной структуре двигательного аппарата (плечевого пояса и конечностей). Общее впечатление при выраженных случаях довольно характерное, среднего роста плотная фигура, с мягким широким лицом на короткой массивной, сидящей между плечами, шее, основательный жирный живот выступает из расширяющейся книзу глубокой сводчатой грудной клетки. Если мы рассмотрим конечности, то заметим, что они мягкие, круглые, со слабовыраженным рельефом мышц и костей, часто изящные; руки мягкие, короткие и широкие. Кистевые суставы и ключицы нередко стройные и нежные. При этом плечи не выступают широко как у атлетиков, но (главным образом у старых людей) круглы, не сколько приподняты и сдвинуты вперед, часто спускаются к груди при резком изгибе на внутреннем крае deltoideus. Создается впечатление, что весь плечевой пояс сдвинут вперед и вверх по отношению к несколько вздутой грудной клетке; голова принимает участие в этом статическом перемещении она опускается вперед между плечами, так что короткая, толстая шея постепенно почти исчезает и верхняя грудная часть позвоночника делает легкий кифотический изгиб. В профиль шея не выглядит, как у других типов, стройной, круглой колонной, на которой держится резко выступающая и отграниченная голова, а в выраженных случаях среднего и более пожилого возраста кончик подбородка косой линией непосредственно связан с верхним концом грудины (см. рис. 3) Пропорция груди, плеч, шеи, не касаясь конфигурации лица, характера ожирения, является самой интересной в пикническом строении тела. Отношение умеренном ширины плеч к большому объему груди отличается от соответствующих пропорций у атлетиков, где объем груди незначителен по сравнению с чрезмерной шириной плеч. Если туловище атлетического типа кажется широким, то пикнического - глубоким. Конечности в среднем скорее коротки, чем длинны (см. табл. 3).
Таблица 3Основные размеры тела у людей пикнического типа (в среднем)
| Признаки |
Мужчины |
Женщины |
| Вес (кг) |
68.0 |
56.3 |
| Длина тела (см) |
167.8 |
156.5 |
| Ширина плеч (см) |
36.9 |
34.3 |
| Объем груди (см) |
94.5 |
86.0 |
| Объем живота (см) |
88.8 |
78.7 |
| Объем в области бедер (см) |
92.0 |
94.2 |
| Объем предплечья (см) |
26.5 |
22.4 |
| Объем кисти (см) |
20.7 |
18.6 |
| Объем икр (см) |
33.2 |
31.3 |
| Длина ног (см) |
87.4 |
80.5 |
Пикники имеют определенную тенденцию к ожирению. Характер ожирения также типичен и должен быть сразу замечен не для сравнения с атлетиками и астениками, которые не проявляют склонности к ожирению, а для сопоставления с известными грубодиспластическими типами. Ожирение пикников держится в умеренных границах и в первую очередь проявляется в склонности к ожирению торса, жир преимущественно отлагается в компактном жирном животе. Все остальные формы тела благодаря диффузному отложению жира мягки и закругленны, но не скрыты и не уродливы… Кожа не дряблая, как у астеников, не упругая, как у атлетиков, а мягкая и хорошо облегает тело. Она средней толщины с сильными изгибами контуров, в особенности у скуловых костей. На наружной поверхности плеч кожа натянута. Мышцы средней силы, но мягкой консистенции. Рост пикников средний - 167,8 см. Относительно сильное отложение жира сказывается на том, что, в противоположность другим типам, а также атлетическому, вес тела у пикников превышает рост (68,0). Вес тела свыше 100 кг наблюдается редко и только в определенные периоды жизни. Наибольший вес у пикников в нашем исследовании был равен 107 кг при росте 171 см. Тем не менее мы встречаем особенно у старых лиц значительное снижение веса как следствие сильной инволюции. Пикники нередко обнаруживают в зависимости от периодов и смены психотических фаз сильные и резкие колебания веса тела. В упомянутом случае приблизительно на 30-м году началось сильное повышение веса, который после многочисленных колебаний достиг 107 кг, а затем в 60-летнем возрасте одновременно с психической депрессией быстро снизился до 76 кг и после депрессии больше не поднимался. Небольшие коренастые фигуры среди пикников нашего населения встречаются довольно часто, но только один из обследованных нами был ниже 160 см. Очень высокие пикники бывают редко. Мы обнаружили только два случая сростом 181 и 182 см (при наличии атлетических элементов строения тела), которые перешли границу 180 см. Пикнически и тип является достаточно очерченным и не обнаруживает каких-либо неожиданных вариантов. Следует подчеркнуть, что он, благодаря строению скелета и прежде всего не зависящим от жирового слоя размерам черепа лица и кистей, часто имеет характерные пропорции груди, плеч и шеи. Для его диагноза вовсе не требуется наличия более сильного жирового слоя. Грубые очертания тела существенно различаются взависимости оттого, имеетли пикник жирный живот или толстую шею. Если не забывать того, что большинство рабочих тяжелого труда и лица моложе 35-40 лет не обладают компактным пикническим жировым слоем, то можно понять, почему возможны ошибочные диагнозы, если полагаться только на этот весьма важный и производящий впечатление, но не всегда постоянный симптом. Присоединение признаков других типов может совершенно затушевывать пикническую картину, но при тщательном рассмотрении и измерении обнаруживаются характерные пикнические компоненты. Комбинации с атлетическими элементами нередки - в этом случае плечи шире и конечности более костлявы и грубы. Астенически-пикнические структурные интерференции наблюдаются в следующем сочетании: небольшой жирный живот, длинный thorax, длинные узкие конечности. Кроме того, в конфигурации лица и черепа можно отметить легкую оксицефалию с мягкими пикническими щеками и широкими челюстями. Здесь, как и при других типах, возможно бесчисленное множество таких комбинаций; нет вообще ни одного отдельного симптома, который не комбинировался бы со стигматами другого типа. Морфологические различия у пикников разных возрастных групп значительно больше, чем при других типах. Пикнический тип приобретает свою характерную форму в более зрелом возрасте, между 30-40 годами, а после 60 лет стирается. Эти различия зависят прежде всего от жирового слоя и отчасти от вторично обусловленного этим изменения формы грудной клетки. Встречаются случаи, когда жирный живот и одновременное с ним расширение нижней апертуры грудной клетки развиваются вскоре после 20 лет. Но это исключения. В большинстве случаев у молодых пикников в возрасте 20-30 лет мы находим следующий габитус: широкая, нежная форма лица с правильными пропорциями и характерной нижней челюстью, короткая, чаше толстая, но весьма плотная и резко отграниченная от нижней поверхности подбородка шея. Thorax еще не вздут, соблюдена гармония между шеей и плечами, поэтому нет еще кифоза и не отмечается характерного наклона головы вперед между приподнятыми плечами. При таком телосложении молодой пикник на первый взгляд может быть принят за атлетика… …В старческом возрасте жирный живот мало заметен, он иногда западает, и тогда грудная клетка не вытягивается кверху. Кожа дряблая и сухая. Существенные признаки строения тела, однако, сохраняются.
(Пикнический тип у женщин) Пикническое строение тела у женщин несколько модифицируется. Жир откладывается главным образом на торсе, но его больше на груди и бедрах. Соотношение груди и плеч такое же, как у мужчин. По объему груди и бедер пикнические женщины относительно превосходят атлетических. Низкий рост (меньше 150 см) у них наблюдается довольно часто. Минимальный рост наших пациенток был 145 см. Очень молодых пикнических женщин с изящным строением тела, не имеющих значительного жирового слоя, при первом взгляде можно принять за атлетических. Такой ошибки мы избежим, если уделим должное внимание данным измерения, конфигурации лица, формам, которые и здесь являются более круглыми и полными. Молодые пикнические мужчины с хорошей мускулатурой и свежим тургором кожи на первый взгляд могут казаться атлетическими. Если форма лица и пропорции между грудью и плечами типичны, то их смешать нельзя. В отдельных случаях дифференциальный диагноз может быть довольно сомнительным. Вообще при сравнении юношеских фотографий пожилых циркулярных больных бросается в глаза, что отдельные мужчины к 20 годам обнаруживали совершенно атипические формы тела, удлиненное лицо, узкое телосложение, которые позже развились совершенно пикнически. Поэтому надо быть весьма осторожным с отрицательным суждением о молодом циркулярном, так как до 40 лет нельзя с уверенностью утверждать, что отсутствуют пикнические компоненты. Вопрос о смене явлений именно при этих эпизодически наступающих пикнических компонентах играет немаловажную роль. У нас было только двое циркулярных моложе 17 лет; оба при хорошо выраженных, округленных формах обнаруживали явное недоразвитие для своего возраста. Из-за недостаточности наблюдений нельзя утверждать, что в этом проявляется определенная закономерность.
(Циркуляная и шизофреническая группы. Ограничения методики) Прежде чем перейти к более тонкой диагностике строения головы и поверхности тела, следует дать обзор того, как выражаются в цифрах типы строения тела циркулярной и шизофренической групп. Следует отметить, что, само собой разумеется, не существует резкой разницы между отдельными типами, и, следовательно, отнесение пограничных случаев к той или иной группе не может быть точным. У циркулярных случаи с несомненным преобладанием пикнических структурных элементов мы выделили в отдельную рубрику (58 случаев). Остальные 14 - это смешанные варианты, которые обнаруживают явные пикнические симптомы строения тела, но вместе стем и сильные гетерогенные налеты, например пикнически-атлетические (5 случаев) и пикнически-астенические смешанные картины. У шизофреников астенически-атлетические смешанные формы мы также рассматриваем особо. Некоторые черты других типов в деталях строения тела мы, разумеется, сможем установить в большом количестве "чистых" случаев. Им следует уделить должное внимание; в связи с наследственностью, характерологией и психотическим построением симптомов они могут быть весьма интересны. В общем статистическом обзоре они не играют никакой роли. Картина, которую дает таблица 4, является показательной и имеет большое биологическое значение (см. табл. 4).
Таблица 4Строение тела и психическая предрасположенность
| Типы |
Циркулярные |
Шизофреники |
| Астеническое |
4 |
81 |
| Атлетическое |
3 |
31 |
| Атлетически-астеническое смешанное |
2 |
11 |
| Пикническое |
58 |
2 |
| Пикнические смешанные формы |
14 |
3 |
| Диспластическое |
- |
34 |
| Расплывчатые картины и не внесенные в рубрики |
4 |
13 |
| Всего |
85 |
175 |
Разумеется, нельзя полагаться на абсолютные цифровые данные. Надо считаться с пограничными случаями и возможностью возникновения ошибок из экзогенных факторов. Следует, прежде всего, помнить, что наши исследования хотя и относятся во всех группах к самым разным внешним условиям жизни, профессиям и возрастам, но все же при этом использовано преимущественно швабское население, поэтому необходимо тщательно проверить и сопоставить этот материал с соматическим состоянием других. Наше подробное сравнение портретов различнейших немецких племен и других европейских народов позволяет предположить, что и там можно встретить приблизительно сходные, хотя в деталях не совсем одинаковые, соотношения между строением тела и психической предрасположенностью. При всех этих оговорках мы можем установить для швабского населения следующее:
· Мы находим у циркулярных больных среди большого числа смешанных и неясных картин значительное преобладание пикнических типов строения тела, с одной стороны, при слабом участии классически выраженных астенических, атлетических и диспластических форм тела - с другой.
· Напротив, у шизофреников мы находим среди известного числа гетерогенно-смешанных и неясных картин явное преобладание астенических, атлетических и диспластических типов (или их комбинаций), с одной стороны, при слабовыраженном участии классических пикнических форм тела - с другой.
Следовательно, мы можем сделать выводы, что:
· между психической предрасположенностью к маниакально-депрессивным заболеваниям и пикническим типом строения тела существует ясное биологическое родство;
· между психической предрасположенностью к шизофрении и строением тела астеников, атлетиков и некоторых диспластиков существует ясное биологическое родство;
· напротив, существует незначительное родство между шизофренией и пикническим типом строения тела, с одной стороны, и циркулярным психозом и астенически-атлетически-диспластическим типом строения тела - с другой.
12. Видовое единство человека. Понятие о расе
Видовое единство человечества
История науки знает неоднократные попытки доказать, что человеческие расы представляют собою различные виды и даже роды, однако большинство исследователей определяет человечество как единый вид.
Все расы при смешении дают плодовитое потомство. Поэтому не удивительны высокие цифры рождаемости населения Южной Америки, Южной Азии и других областей. В Бразилии количество "чистых" индейцев, живущих в лесах, составляет 2% от общего количества населения, а количество метисов - 33%.
Все зоны соприкосновения разных расовых типов всегда являлись одновременно зонами смешения. Стихийный процесс расового смешения не могут остановить социальные преграды и террористические меры, направленные на сохранение "чистоты" расы. Некоторые расы выглядят внешне достаточно контрастно (пигмеи Африки и северо-американские индейцы, северные европейцы и южные китайцы), но все человеческие расы связаны рядом промежуточных типов.
Понятие расы
Все ныне живущее человечество с точки зрения биологической науки представляет один вид Homo sapiens ("человек разумный"), распадающийся на более мелкие подразделения, именуемые расами. Термин "раса" созвучен арабскому "ras" ("начало", "происхождение"), итальянскому "razza" ("раса", "племя").
Расы человека отличаются друг от друга рядом признаков - цветом кожи, волос, глаз, типом волос, чертами лица, формой черепа, пропорциями тела и другими не существенными для жизнедеятельности признаками. Расовые признаки связаны с наследственностью и мало меняются под воздействием среды.
Расы связаны с определенной территорией (ареалом).В настоящее время данная связь может не обнаруживаться (черная раса представлена в населении Африки и Америки), однако она существовала в прошлом. Не все биологические признаки человека являются расовыми признаками. Например, развитие жироотложения, мускулатуры, различия в осанке - не расовые отличия, так как связаны с влиянием среды сейчас.
На Земле уже насчитывается около 6 млрд. людей. Среди них нет, и не может быть двух совершенно одинаковых людей; даже близнецы, развившиеся из одного яйца, несмотря на огромное сходство их внешнего облика, и внутреннего строения, всегда какими-то мелкими чертами отличаются друг от друга. Наука, изучающая изменения физического типа человека, известна подименем "антропологии" (греч, "антропос" – человек). Особенно сильно заметны телесные различия между территориальными группами людей, удаленными друг от друга и живущими в различной естественно-географической обстановке.Деление вида Homo Sapiens на расы произошло два с половиной столетия назад. Происхождение термина "раса" точно не установлено; возможно, что он представляет собой видоизменение арабского слова "рас" (голова, начало, корень). Есть также мнение, что термин этот связан с итальянским razza, что значит "племя". Слово "раса" примерно в том смысле, как оно употребляетсятеперь, встречается уже у французского ученого Франсуа Бернье, который опубликовал в 1684 г. одну из первых классификаций человеческих рас.Расы - это исторически сложившиеся группировки (группы популяции) людейразной численности характеризующиеся сходством морфологических и физиологических свойств, а также общностью занимаемых ими территорий.Развиваясь под влиянием исторических факторов и относясь к одному виду(H.sapiens), раса отличается от народа, или этноса, который, обладая определенной территорией расселения, может содержать несколько расовых комплексов. К одной и той же расе может принадлежать ряд народов и носителей многих языков. Большинство ученых сходятся во мнении, что существует 3 крупных расы, которые в свою очередь распадаются на более мелкие. В настоящее время по мнению разных ученых, насчитывается 34 – 40 рас. Расы отличаются друг от друга по 30-40 элементам. Расовые особенности наследственны и являются приспособительными к условиям существования.Целью моей работы является систематизировать и углубить знания о человеческих расах.13. Социальные корни расизма. Научная несостоятельность расистских теорий
Антропология (наука о человеке) как самостоятельная дисциплина начала развиваться во второй половине 18- го века во Франции и Германии. Ее границы с этнологией, биологией, анатомией человека тогда еще не были четко очерчены, но она считалась точной наукой и на начальной стадии занималась изучением различных человеческих рас. (Только позднее она превратилась в общественную дисциплину.)
Интерес к расам возник в мире вследствие развития путешествий и последующих описаний увиденного, а также распространения в мире колониализма. Вначале антропология имела лишь описательный характер, затем появляются различные классификации.
В 1738 году швед К.Линней опубликовал "Систему природы" ("Systema nature"), в которой разделил человечество на четыре расы, описав их физические и нравственные черты. Позже было опубликовано еще несколько аналогичных сочинений, и хотя мнение о принципах деления на расы у разных авторов были различные, все они основывались, прежде всего, на цвете кожи представителей рас.
Наконец, британский врач (живший, однако, во Франции) У.Ф.Эдвардс произвел научную революцию, разделив расы по телосложению; в 1829 году он опубликовал знаменитую книгу "О физиологических свойствах человеческих рас в связи с их историей". Ее новизна заключалась, в частности, в том, что в отношении французов он пришел к мнению, что они происходят от двух различных рас и, одновременно, что эти расы существуют в несмешанном виде до сих пор (т.е. в 19-ом веке). Он также восстановил в правах термин "исторические расы", вследствие чего появилась возможность выделения разных расовых групп, хотя бы и имевших одинаковый цвет кожи. В связи с этим он указал и на физические особенности части евреев. Эдварде не сомневался, что самым достоверным признаком расы является размер и форма черепа; рост и цвет волос тоже важны, но они играют второстепенную роль (дополнительно он использовал для различения рас еще и итальянские исследования в лингвистике по особенностям произношения).
Уверенность Эдвардса в преобладающем значении формы черепа для различения рас была неслучайной: десятилетиями, с конца 18 века, процветала в Европе френология, "наука об изучении человеческого черепа", утверждавшая, что особенности мозга связаны с теми или иными чертами его "хранилища" - черепа. Размер той или иной части черепа определяет, якобы, ту или иную характеристику самого мозга, следовательно, человеческий характер можно узнать, изучив тот или иной череп. В начале 40-х годов 19 века швед Рециус предложил коэффициент для измерения черепов (длина черепа делится на его ширину и умножается на 100) - так называемый cephalic index. Многие годы эта формула служила исходной точкой в исследованиях не только псевдо- но и настоящих ученых.
В 1893 году французский философ Альфред Фуйе предсказал - не осознавая в полной мере силу своего предвидения: "Люди еще будут убивать друг друга из-за разницы в один балл между их черепами".
Итак, антропология изучала тогда расовые проблемы, связанные преимущественно с особенностями телосложения, с привлечением для этого достижений общественных наук.
Обращаем особое внимание на этот переход науки от сухого научного факта к социальным спекуляциям; подобный процесс, как мы убедимся далее, происходил и в других отраслях знания. Повсеместное его распространения было связано с тем абсолютным значением, которое общество придавало научным открытиям.
Многие ученые пришли к выводу, что существует огромная разница не только во внешнем облике, но и в характерах представителей разных рас. В этой среде было принято считать, что различие цвета кожи, толщины губ, строения глаз, качества волос обуславливает природное неравенство людей. "Обязательно должна обнаружиться связь между внутренними и внешними различиями рас" - историки романтической школы оказались первыми, кто принял этот тезис в своих научных комментариях.
Так был сделан первый шаг к расизму, развившемуся в полной мере в первой половине 19-го века.
У идеологического расизма была, конечно, связь и с общественно-политическими процессами развития национализма и колониализма, но эта связь переживала еще так называемый "наивный" - младенческий период; пока у людей еще оставалась возможность быть расистами, отрицая насильственный и иерархический его характер.
Новая стадия расизма, вскоре ставшая доминирующей, началась, когда в систему антропологических проблем стали вплетать проблемы биологические.
Биология
В отличие от общепринятой в то время Библейской версии о единовременном сотворении мира и идентичности современной природы с той, которая была сотворена Творцом, в 19 веке появилась биологически обоснованная теория эволюции природы.
Одним из первых эволюционистов стал французский биолог Жан-Пьер Ламарк, предположивший, что разнообразие видов, форм, оттенков живых существ есть результат их приспособления к окружающей среде. Длительные процессы привели, в конце концов, к изменению природных форм. Но теория Ламарка строилась, в основном, на предположениях и была плохо обоснована. Тем не менее, она распространилась во Франции и других странах, ибо время от времени гипотезу Ламарка подкрепляли вновь открытые феномены. Летом 1830 года во Французской Академии состоялся горячий диспут о достоверности ламаркизма, немецкий поэт Гете внимательно следил за ходом этих споров.
Несколько позже в Англии нашлось множество сторонников теорий социолога Герберта Спенсера, обосновавшего в 50-х годах свои социологические гипотезы именно новейшими биологическими открытиями. Окончательно сформулировал и доказал теорию эволюции знаменитый англичанин Чарльз Дарвин: его "On origin of species" ("Происхождение видов"), впервые опубликованное в 1859 году, переиздавалось бесчисленное множество раз на всех языках Европы. Биолог обосновал процесс эволюции в природе и сформулировал его главнейшие законы: закон естественного отбора и закон борьбы за существование.
На научные гипотезы Дарвина жадно, как на сокровищницу, набросились люди, решившие использовать их для разработки социальных тем (отсюда и возник термин - "социальный дарвинизм"). Вот главные тезисы "социал-дарвинистов":
а. природа построена на эволюции, и самая высшая ступень эволюции - млекопитающие, среди которых первенствуют приматы, а венчает пирамиду человек - хомо сапиенс;
б. у эволюции "нет границ" (Дарвин), но если это так, то, следовательно, среди людей нынче тоже протекает процесс эволюции. Следовательно, внутри современного человечества можно и нужно искать различные виды, а среди них - самый высший.
Так, уцепившись за уже существующий расизм, социал-дарвинисты создали расистский дарвинизм. Но как можно было среди прочих рас выделить высшую расу? Расисты начали утверждать, что таковой является белая раса:
- в войнах белый человек завоевал мир и создал повсюду свои колонии, духовно поработив все народы;
- в сфере духа породил величайших философов-европейцев;
- в музыке - создал классику;
- в архитектуре белыми созданы величайшие творения, например, - готические соборы;
- в технологии - паровозы и пароходы - изобретения белого человека, заставившего природу работать на себя и т.д.
Сразу видно, что это субъективные критерии, основанные как на посылке на собственном же выводе, предполагавшем, что белая раса - высшая. Но в свое время (а, может быть, не только в свое) они считались вполне объективными, доказывающими подлинное различие рас в талантах и интеллекте.
Второй вывод, как бы дополняющий первый, был связан с "борьбой за существование".
Считая, что раз борьба за существование есть закон природы, и, следовательно, она не может носить отрицательного характера, социал-дарвинисты утверждали, что возражение пацифистов-моралистов против войн не обоснованы. Более того, лица, отрицавшие справедливость "законов природы", возможно, и есть "злоумышленники", "слабые", придумавшие эти гуманистические теории, чтобы с их помощью предотвратить свое поражение от руки "высших", "лучших" по природе. Мир и гармония в среде народов, проповедовавшиеся в Европе ранее, перестали теперь казаться обязательными даже для нравственных людей.
Лингвистика
Развитию расистских идей содействовали исследования в еще одной науке - в лингвистике (языкознании), развивавшейся с конца 18 века.
Готфрид фон Гердер в книге "Происхождение языка" (1772 г.) обратился к проблеме рождения языков: по его мнению, в основе появления речи у человека лежал инстинкт. Когда европейцы открыли для себя санскрит, язык праиндийцев, и узнали о его близости к европейским языкам, в науке возникла новая лингвистическая школа (вместо признанной со времен Римской империи) - сравнительно-историческая.
Англичанин Вильям Джонс, первым исследовавший санскрит, предположил, что у готского, индийского, древнегреческого, латыни и, возможно, древнеперсидского языков - есть общий предок (1770 г.). Вслед за ним эту идею развил немец Фридрих Шлегель в книге "О языке и мудрости индийцев", 1880. Вскоре языковед Клафорт ввел термин "индоевропейская группа языков". Повлияли на развитие новой лингвистики и датские языковеды, занимавшиеся тевтонскими языками скандинавских эпосов, в частности - древнеисландским языком.
Новый вклад внес в лингвистику той эпохи Якоб Гримм, зачинатель германской языковедческой школы, романтик, изучавший как фольклор, так и древнюю немецкую литературу; он обнаружил древнейшие выражения в сохранившихся народных диалектах. Вместе с братом Вильгельмом ("братья Гримм") он создал новую немецкую грамматику (1819 г.) и важнейший словарь (1852 г.).
В 20-х годах 19-го века исследователи индоевропейских языков начинают употреблять новый профессиональный термин - "ариец". В санскритских легендах этим словом обозначали древних завоевателей Индостана, и дословное его значение, по-видимому, - "хозяин", "аристократ" или "чистый", т.е. отличающийся от аборигенов.
Эти исследование были использованы расистами в своих целях. В частности, было сделано предположение, что все племена, говорящие на индоевропейских языках, состоят в кровном родстве (возможность кровного родства без родства языков, как и родства языков без кровного родства, представлялась невероятной). Так и возникли термины "арийский народ" и "потомки арийцев", хотя существовали многочисленные разногласия о том, какие именно народы должны входить в "арийскую группу". Одним из главных сочинений подобного рода стала книга швейцарца Адольфа Пише "Индоевропейские источники или примитивные арийцы". В предисловии он утверждал, что именно санскритские корни помогут понять историю человечества: в среде индоевропейцев живет "раса, которая с помощью Высших сил будет властвовать над миром". Она "выделяется красотой своей крови и интеллекта", доказательством чему служит "богатство гармоний и совершенство структуры ее языка".
19-й век дал толчок изучению еще одного пласта лингвистики - восточных языков. Катализатором этого процесса послужило открытие и расшифровка аккадского языка, сыгравшего в истории роль, подобную роли санскрита для индоевропейских. Среди лингвистов-востоковедов, в особенности тех, кто изучал иврит, выделялся в середине века француз Эрнест Ренан. Он и дал группе языков Ближнего Востока название "семитские языки", использовав библейское сказание о происхождение человечества от Шема (Сима), Яфета и Хама, сыновей Ноаха (Ноя).
В 1855 году, опубликовав "Общую историю и сравнительную систему семитских языков", он обосновал в ней общность группы этих языков и народов, использовав для их обозначения термин "семитские народы" (более подробно он развил те же идеи в "Новых мыслях об общности характера семитских народов" Сказанное выше кратко обрисовывает как истоки расизма, так и методологию расистской аргументации.
Истоки расизма
Итак, в 19 веке сложился подходящий общественный климат для внедрения расизма в духовную жизнь общества. Все его составляющие оказались объединены в книге французского аристократа-дипломата графа Артура де Гобино "О неравенстве человеческих рас" (1853-55 гг., т.е. еще до "Происхождения видов" Дарвина - это доказывает, что идеи витали в воздухе). Во Франции, на родине автора, его книга не пользовалась популярностью, но в Германии, благодаря его близкой дружбе с великим композитором Рихардом Вагнером, на сочинение Гобино откликнулись многие. Кроме вышеупомянутых постулатов ("ключ к пониманию истории таится в расовых особенностях ее субъектов", "критерии величия белой расы", похвала арийцам и проч.), творение Гобино отличалось также крайним историческим пессимизмом. Из-за недооценки миром важности расового фактора все расы, мол, перемешались, и потому в будущем нельзя будет отыскать силу, которая поведет человечество к прогрессу:
"Счастливая случайность помогла выполнить главную задачу... но мало шансов на счастливое будущее". Гобино считал, что явно поздно исправлять испорченную историю человечества.
Другие расисты с ним не соглашались: дарвиновские идеи о возможности повлиять на "природу", т.е. "улучшить расу", их явно ободряли. Кроме того, эту группу идеологов объединяло трагическое ощущение необходимости и срочности каких-то действий для "спасения мира", "гонки наперегонки со временем".
Следует отметить попутно, что лично Гобино относился к евреям с определенной симпатией, хотя и считал их расой ниже арийцев. Он ценил их, как он выражался, за "мудрость народа", понявшего еще во времена Эзры и Нехемии, т.е. в 5-ом веке до нашей эры, важность принципа чистоты расы и воспринявшего объявленный запрет на смешанные браки.
Расизм и его социальные корни
С расистской точки зрения, человечество делится на "высшие" и "низшие" расы, причем различия между ними наследственны и не могут изменяться под влиянием внешних условий, вследствие чего никакие социальные преобразования не могут превратить "низшую" расу в "высшую" или даже сгладить их различия. "Низшие" расы, по утверждению расистов, всегда подчинялись "высшим", а в тех случаях, когда по каким-либо случайным обстоятельствам этого не происходило, культура деградировала, и средством для ее возрождения могло служить только приведение "низшей" расы к покорности или полное ее уничтожение.
Антропология была использована для "доказательства" того, что все те явления, которые в действительности, неизбежно вытекают из определенной системы производственных отношений на современном историческом этапе.
Для этой цели была создана особая отрасль тенденциозной публицистики, имеющая некоторое внешнее сходство с антропологией, но в действительности менее всего заинтересованная в фактах и их научном освещении <...> (с. 511—512).
Антропосоциология, социальный дарвинизм и их реакционная сущность
Расизм становится системой, пытающейся объяснить основные закономерности общественной жизни, исходя из антропологических фактов. Вырастает так называемая антропосоциология, которая сводит проблему развития общества к конкуренции различных расовых типов. <...> Важнейшим из всех направлений, близких к антропосоциологии, был социалдарвинизм. Число авторов конца XIX в., развивавших "концепции" о том, что европейские расы являются высшим продуктом естественного отбора и что руководящие классы — это победители в борьбе за существование, чрезвычайно велико. <...> Дарвину принадлежат важнейшие доказательства огромного сходства между расами.
Таким образом, социалдарвинисты в расовой проблеме развивали концепции, глубоко враждебные и идеям, и чувствам самого Дарвина. <...> Основателем современного евгенического движения был Гальтон.
Защитники наиболее реакционных вариантов евгенической теории вскоре выступили с проектами принудительной стерилизации "наследственно неполноценных" людей. В ряде стран эти проекты получили юридическое оформление и частично практически осуществлялись.
Невиданные размеры приняла расистская фальсификация науки на службе германского фашизма. Приход гитлеровцев к власти ознаменовался превращением расизма в основу государственной идеологии Германии.
Научная несостоятельность расистских теорий
Необоснованность расистских теорий можно показать на примере одного из самых распространенных положений расизма, а именно на утверждении, что североевропейская раса — это наиболее духовно одаренная раса человечества, которой мир якобы обязан созданием всех индоевропейских языков, а также культур тех народов, которые говорят на этих языках.
Прежде всего следует обратить внимание на полное несоответствие этой "теории" картине распространения индоевропейских народов. Исчисляющееся многими десятками миллионов темноволосое и темноглазое население Южной Европы, исключая небольшой народ — басков, говорит только на индоевропейских языках; к этой же семье языков относится язык иранцев и северных индийцев, в подавляющей массе весьма темноволосых и темноглазых.
Положение об "исключительной одаренности" северной расы расисты "основывают" в значительной степени на современном развитии техники и цивилизации у тех европейских народов, антропологический тип которых относится к северной расе.
Однако и это положение не выдерживает критики.
Творчество человеческого гения уже горело ярким пламенем в Месопотамии, в Египте, в долине Инда (Мохенджо-Даро), на Крите, в то время как север Европы еще был объят глубоким мраком. Более поздние цивилизации — Карфагена, Греции, Ирана, Бактрии, Согдианы, Хорезма, Рима, Аксума (в Абиссинии) — также возникли задолго до расцвета культуры у северных европейцев. Для развития античной культуры исключительное значение имела древнеегипетская культура. Здесь уместно напомнить, что цвет кожи древних египтян был очень смуглым, близким к коричневому и общий их тип довольно близок к эфиопской восточноафриканской расе.
Совершенно неверно представление, что негрская Африка была всегда "страной хижин". Большие и многочисленные древние города имеются в области иорубов, далее — между Тимбукту и Ниамини, где они особенно часты, и, наконец, у гауссов. Город Ибадан в прошлом веке имел свыше 150 тысяч жителей.
Человечество, включая все его расы принадлежат к одной единой стадии развития человеческого рода. Выделившись из среды своих предков, новый вид человека вступил в мир, уже обладая комплексом свойств, необходимым для речевой и мыслительной деятельности. Это наследие было получено от общего предка всеми ныне живущими расами. Опираясь на это последнее, люди любых рас, независимо от различий в "темпераменте", обладают способностью к бесконечному продолжению своего культурного роста" (с. 512—516).
Чебоксаров Н.Н., Чебоксарова И.А. "Народы, расы, культуры", 1985:
"В сентябре 1967 г. в Париже состоялось совещание экспертов ЮНЕСКО, принявшее "Декларацию о расе и расовых предрассудках", в которой подчеркивается, что "все современные люди относятся к одному виду и происходят от одного корня", что "современная биологическая наука не позволяет относить различия в культурных достижениях народов за счет различий в их наследственных свойствах", что "расизм грубо фальсифицирует данные биологии человека... препятствует развитию тех, кто от него страдает, развращает тех, кто его исповедует, разделяет нации между собой, усиливает международную напряженность и угрожает миру во всем мире". "Основные средства борьбы с расизмом, — читаем мы в "Декларации", — заключаются в изменении общественных условий, порождающих предрассудки: в противодействии поступкам лиц, убеждения которых заражены предрассудками, в борьбе с самыми ложными убеждениями" (с. 166).
Несостоятельность расизма
Расселение популяций неоантропов в Европу, Азию и Австралию, по Берингийскому мосту на Американский континент, их дальнейшая изоляция, привели к морфологическим адаптациям, приспособлениям к раз-личным климатическим условиям. Сформировались большие и малые человеческие расы - систематические подразделения внутри вида Homo sapiens, к которому относится все население Земли. Различают три большие расы: евразийскую - европеоидную, экваториальную - австрало-негроидную, и азиатско-американскую - монголоидную. Внутри каждой расы выделяют малые расы и расовые группы. Все расы относятся к одному виду, об этом свидетельствует плодовитость межрасовых браков. Кроме того, все расы равноценны в биологическом и психологическом отношениях. Для негроидной расы характера темная кожа, курчавые волосы, широкий и плоский нос. Темная кожа нег-роидной расы за счет пигмента меланина предохраняет организм от избытка ультрафиолетовых лучей и избыточного образования витамина D. Противорахитный витамин D образуется в коже под действием ультрафио-летовых лучей и необходим для поддержания баланса кальция в организме. Если слишком много витамина D, кальция в костях больше нормы, они становятся хрупкими. У европейцев, живущих в широтах с меньшим ко-личеством солнечной радиации, кожа светлее, в ней меньше меланина, соответственно образуется достаточное количество витамина D. Монголоидная раса характеризуется кожей с желтоватым оттенком, плоским лицом с широкими скулами, прямыми черными волосами, разрезом глаз и развитым эпикантусом - припухшим верхним веком. Эти особенности являются адаптациями к жизни в определенных условиях освещенности на открытых пространствах. В каждой расе есть люди, считающие свою расу особенной, высшей. Расисты придерживаются гипотезы полицентризма, считают, что расы имеют различное происхождение, что есть "высшие" и "низшие" расы. Какие-то расы якобы произошли от кроманьонцев, какие-то от неандертальцев, какие-то вообще от питекантропов. Экономическую и культурную отсталость некоторых народов они объясняют расовой неравноценностью, а не общественно-экономическими факторами. Но никаких научных доказательств в пользу расовой неравноценности нет. Морфологические особенности рас - результат адаптаций к конкретным условиям жизни.
14. Экология дифференциации человека
Экология человека
Экология — наука о взаимоотношениях организмов между собой и с окружающей средой. Экология человека изучает закономерности взаимодействия человеческого общества и окружающей среды.
Биосоциальная природа человека
Человек - биосоциальное существо, генетически связанное с другими формами жизни, но отделившееся от них благодаря способности производить орудия труда; обладающее членораздельной речью и сознанием, творческой активностью и нравственным самосознанием; высшая ступень развития живых организмов на Земле.
Биосоциальная природа человека находит отражение в том, что его жизнь определяется единой системой условий, в которую входят как биологические, так и социальные элементы.
Биологические элементы
1. Наследственность
2. Изменчивость
3. Естественный отбор
Социальные элементы
1. Труд.
2. Речь
3. Организация совместной жизни
Эволюция биосферы и ее основные составляющие
Биосфера (с греч. bios - "жизнь", sphaira - "шар, сфера") - оболочка Земли, населенная организмами, составляющими в совокупности живое вещество планеты. Биосфера - это сфера жизни, расположенная в земной коре, воде и воздухе, простирающаяся примерно от 10 км вглубь Земли до 30 км над Землей. Жизнь, по В.И.Вернадскому, не только зависит от среды, но и сама производит колоссальную геохимическую работу. Идеи В.И.Вернадского об эволюции биосферы могут быть сформулированы следующим образом:
1. Вначале сформировалась литосфера — предвестник окружающей среды, а после появления жизни на суше - биосфера.
2. В течение всей геологической истории Земли никогда не существовало периодов, лишенных жизни. Поэтому современное живое вещество генетически связано с прошлыми геологическими эпохами.
3. Живые организмы — главный фактор миграции химических элементов в земной коре.
4. Геологический эффект деятельности организмов обусловлен тем, что их количество очень велико и действуют они в течение большого промежутка времени.
5. Основным движущим фактором развития процессов в биосфере является биохимическая энергия живого вещества. В учении о биосфере В.И.Вернадского заложены основы современных представлений о взаимосвязи и взаимодействии живой и неживой природы.
Наследственность человека
Генетическая программа homosapiens определяет его как биологический вид. Элементарной единицей наследственности является ген, характеризующийся рядом признаков. Гены располагаются в ядрах клеток. По своему уровню ген - внутриклеточная молекулярная структура. По химическому составу это нуклеиновые кислоты, в составе которых основную роль играют азот и фосфор. Гены - это "мозговой центр" клеток. Генетическая программа записана в молекулах ДНК и является достаточно консервативной.
Генетические процессы
От поколения к поколению ДНК человека вовлекается в разнообразные генетические процессы:
1. Мутационный процесс, непосредственно изменяющий структуру ДНК.
2. Миграция генов - их отток или приток из других популяций.
3. Дрейф генов - случайные колебания частот генов.
4. Естественный отбор — процесс, направленно изменяющий частоту генетических признаков.
Мутационный процесс в условиях естественного радиационного фона Земли не может повлиять на жизнь популяций, но повышенный уровень радиации обладает исключительной мутационной активностью, как и химические мутагены.
Случайный дрейф и миграция генов приводит к образованию эко-типов, которые, развиваясь изолированно, могут образовать в результате эволюции новую расу или популяцию.
Естественный отбор сыграл решающую роль в эволюции всех видов, в том числе и homosapiens. Человек современного типа возник в последнюю ледниковую эпоху — примерно 40-50 тыс. лет тому назад. В этот период он занимался охотой, собирательством, позже - скотоводством, земледелием и ремеслами. За последние 2-3 века получило бурное развитие промышленное производство, повлиявшее на величину и качественный характер давления естественного отбора. Человек, являясь биосоциальным существом, не освободился от действия общебиологических закономерностей, универсальных для всего живого. По данным научной литературы, 5 % эмбрионов человека погибает на ранних стадиях онтогенеза (спонтанные аборты), 3 % - смертность до наступления репродуктивного возраста, 20 % взрослых не вступают в брак и 10 % браков бесплодны.
Таким образом, примерно 50 % первичного генофонда не воспроизводится в следующем поколении, значительная часть этих явлений генетически обусловлена.
Влияние факторов окружающей среды на процессы роста и полового созревания человека
Человек на протяжении всей постнатальной жизни постоянно взаимодействует со многими физическими факторами, к которым организму приходится непрерывно приспосабливаться.
Факторы окружающей среды
- Температура окружающей среды
- Сила тяжести (гравитация)
- Уровень инсоляции (солнечная радиация)
- Влажность
- Газовый состав окружающего воздуха
- Геомагнитные поля
- Биологические факторы, внутривидовые и межвидовые взаимодействия
- Социальные факторы
Реакция организма на факторы окружающей среды связана с массой, геометрическими размерами, пропорциями тела и определяет особенности экологии человека.
В настоящее время экологические факторы все активнее воздействуют на людей. Поэтому современной тенденцией физического облика человека стали акселерация и секулярный тренд. Акселерация (от лат. acceleratio) - это ускорение роста людей и проявление их физиологических функций. Термин предложен в 1935 г. немецким врачом Е.Кохом.
Так, в начале XX в. длина тела у мужчин достигала максимума к 25—26 годам, в настоящее время - к 17-19 годам. Начало менструального цикла за последние годы сократилось с 14,5 лет до 12,5 лет.
В развитых странах вес при рождении увеличился на 100-300 гр. Половое созревание подростков происходит на 2 года раньше. Гипотезы причин акселерации весьма различны и противоречивы. Одни авторы считают, что основу акселерации составляют улучшение условий жизни и повышение уровня медицинского обслуживания населения. По мнению других, акселерации способствует появление новых сочетаний генов. Ряд авторов придерживается информационной гипотезы, согласно которой огромный объем информации, обрушившейся на детей с раннего возраста через средства коммуникации, стимулирует ростовые процессы и ускоряет созревание организма. Есть точка зрения, что акселерация - явление временное, связанное с экзогенными (например, зависящими от солнечной активности) или эндогенными (причины которых неизвестны) популяционными циклами, которые на протяжении веков приводили то к ускорению, то к замедлению, ретардации (от лат. retardo -"замедлять, тормозить"), физического развития человечества. Общепризнанной точки зрения на популяционные сдвиги нет до сих пор.
К сожалению, физическая акселерация практически не затрагивала темпов ментального и духовного развития, что создает определенные трудности в сфере обучения и воспитания.
Секулярный тренд (от лат. seculartrend - "вековая тенденция") -это увеличение длины тела, протяженности репродуктивного периода, продолжительности жизни и других свойств человека на определенных стадиях онтогенеза. Например, по обобщенным данным в развитых странах было отмечено увеличение длины тела на 3,5 см у всех родившихся в 1921-1936 гг. по сравнению с прошлым столетием. Природа секулярного тренда также не имеет единой точки зрения.
15. Этническая антропология
Этническая антропология, раздел антропологии, изучающий морфофизиологические особенности отдельных этнических общностей (этносов). Основные единицы исследования Э. а. — ареальные общности людей (популяции), различающиеся по расовым признакам (форма волос, пигментация, размеры и строение лицевого скелета и др.— см. Расоведение). Э. а. тесно связана с геногеографией, т. к. признаки с хорошо изученной наследственностью часто служат отметками (маркёрами), отражающими историю формирования, расселения и метисации популяций и сложившихся на их основе рас. В отличие от растений и животных история популяций человека современного вида протекает не только в окружающей его естественно-географической среде, но и в социально-культурной среде, создаваемой людьми в процессе коллективного труда. В человеческом обществе популяции сопряжены с системой специфических для людей социальных общностей, их подразделений и полиэтнических групп. Анализ взаимоотношений между этносами, популяциями и расами в различных странах и в различные периоды истории позволяет использовать данные Э. а. в качестве исторических источников при разработке многих проблем эволюции человека как биосоциального существа, популяционной генетики, расообразования, этногенеза и других важнейших вопросов антропологии, этнографии, демографии, исторической и медицинской географии.
Расы - это территориальные группы людей, выделяемые на основе их генетического родства, которое проявляется в определенном физическом сходстве. Большинство отечественных антропологов различают три большие расы человечества - монголоидную ("желтую"), европеоидную ("белую") и негроидно-австролоидную ("черную").
Европеоиды светлокожи, для них характерны светло-русые или темно-русые, мягкие или средней жесткости волосы, серые, серо-зеленые или каре-зеленые широко открытые глаза, умеренно развитый подбородок, широкий таз, узкий и сильно выступающий нос, нетолстые губы, обильный волосяной покров тела и лица.
Негроиды темнокожи, у них курчавые или шерстистые темные волосы, толстые губы, очень широкий и плоский нос, крупные зубы, карие или черные глаза, удлиненная форма головы, редкая растительность на лице и теле, узкий таз, большие ступни.
Монголоиды обладают желтой или желто-коричневой кожей. У них прямые жесткие черные волосы, плоское крупное скуластое лицо, узкие раскосые карие глаза со складкой верхнего века (эпикантус) во внутреннем углу глаза, плоский широкий нос, редкая растительность на лице и теле.
Австролоиды имеют шоколадный цвет кожи. У них темные волнистые волосы, крупная голова и массивное лицо с широким и плоским носом, выступающим подбородком, значительный рост волос на лице и теле.
Большие расы делятся на более мелкие. Последние распадаются на группы антропологических типов. Между расами существуют переходные группы, поэтому современное человечество можно рассматривать как переплетение множеств антропологических типов и рас, составляющих единое биологическое целое.
Этническая антропология. От биологии к истории
В предыдущей теме мы познакомились с понятием о расовом многообразии человечества - межгрупповой изменчивости по набору (правильнее сказать - комплексу) так называемых расовых признаков. Рассмотрев некоторые известные классификационные схемы, мы совершили "кругосветное путешествие", по ходу которого обратили особое внимание на антропологические черты, определяющие сходство и различие между отдельными популяциями человека и их крупными объединениями. Были описаны наиболее общие закономерности географической вариабельности антропологических признаков - расовая изменчивость. Полученная нами весьма поверхностная картина, вместе с тем, вполне достаточна для того, чтобы сделать заключение о необычайной "пестроте" древнего и современного населения Земли в биологическом отношении.
Приглядевшись к этому разнообразию, можно сделать еще два взаимосвязанных и бесспорных вывода:
o эта картина не статична и меняется во времени. Иначе говоря, формирование и поддержание биологической изменчивости (популяционных и расовых особенностей) тесно связано с ходом истории человечества;
o справедливо и обратное - антропологический материал, о специфике которого было уже так много сказано, потенциально имеет большое значение для решения исторических проблем, и в особенности вопросов этногенеза.
Изучению этой (биологической) составляющей процесса этногенеза, посвящен специальный раздел антропологии. За направлением закрепилось название "этническая антропология" (хотя корректность такого термина в последнее время вызывает возражения, обоснованность которых разбиралась в предыдущей теме).
Итак, этническая антропология - это раздел антропологии, изучающий морфофизиологические особенности отдельных этнических общностей (этносов).
Говоря более строго, предмет этнической антропологии - история формирования и специфики внутри- и межгрупповой биологической изменчивости этно-территориальных групп человечества и их генетических взаимоотношений. Полученные в результате этих исследований материалы дают возможность выяснить родственные взаимоотношения между группами населения и историю их возникновения, а именно место, древность и последовательность этногенетических событий (этногенезов). Этническая антропология является своеобразной "визитной карточкой" отечественной науки о человеке. На настоящий момент нет ни одной другой страны мира, антропологический состав населения которой был бы исследован с таким масштабом, тщательностью и методической строгостью, как это сделано для территории России и государств бывшего Советского Союза. И на этом пространстве нет, пожалуй, ни одного народа, этническая история которого исследовалась без привлечения данных физической антропологии. Работы в этой области, начатые еще основателями антропологической науки, продолжаются по сей день. В этой теме мы остановимся на конкретных примерах того, как антропологические данные становятся историческим источником и помогают решать вопросы происхождения народов. Вместе с теорией, конечно.
Объекты исследования: этнос и популяция
Что является объектом исследования этнической антропологии? Странный вопрос - раз антропология этническая, значит, она исследует этносы или этнические группы - наиболее очевидный субъект исторических преобразований. Так оно и есть. Но этнос - понятие совсем не биологическое, а мы уже знаем, что антропологи исследуют биологические признаки и биологические же популяции. Как соотносятся эти категории, и являются ли они столь разнородными? "Кто к нам с мечом придет, от меча и погибнет" - слова героя из известного отечественного кинофильма 1938 г. На самом деле, великий русский князь Александр Ярославович Невский (1252-1263) никогда не говорил этих слов. Они были специально написаны П.А. Павленко для патриотического образа, блестяще воплощенного актером Н.К. Черкасовым в фильме гениального С.М. Эйзенштейна. Сама фраза, и три "разноэтничные" фамилии выдающихся деятелей российского искусства упомянуты не случайно. Кто такие "мы"? Кто такие эти "они", которые хотят прийти на нашу землю? Вопросы не требует пространных комментариев. "Я", "мы" и "они" - категории, существующие и развивающиеся в нашем сознании. Эти понятия имеют множество значений, без них невозможна деятельность отдельного человека и жизнь любого объединений людей (семьи, малой группы, коллектива, государства и т.п.). Но смысл слов князя Александра понятен. "Мы" называем себя русскими (татарами, украинцами и т.п.), "они" - по-другому. "Мы" говорим на одном языке и понимаем друг друга. У нас близкая культура, похожие традиции и воспитание. "Мы" крестимся щепотью справа налево, у "них" другая вера (религия или идеология). "Они" живут на земле, где много лесов и дичи, а "мы" ловим рыбу и возделываем пшеницу на другой земле… У "них" пусть даже что-то одно, но не так, как у нас. У нас с ними разная судьба (история), которая, с одной стороны, определяет наши различия, а с другой - сама во многом связана с тем, что мы представляем собой разные общности людей.
Такое единство по языку, религии (или идеологии) и традициям (то есть чертам культуры в целом), а также территории и типу хозяйства характеризует этнические общности людей. Если сложить эти "признаки", можно сказать немного иначе: этнос - объединение людей на основании общего самосознания, самоопределения (этнонима) и истории.
Итак, этнос - историческая, а не биологическая категория (см. Хрестом. 6.1). На первый взгляд, для его изучения в антропологии особого места "не предусмотрено". Однако этнос и популяция сближаются по двум важным критериям. Это:
o единство ареала. Под этим следует понимать не только единство места обитания, то есть географическую локализацию, но и напрямую связанную с ней повышенную степень консолидированности населения. Для этноса эта консолидация является ключевой характеристикой и определяется единством культурной традиции, языком и вероисповеданием. Для биологической популяции она выражается в большей или меньшей степени жесткости популяционной структуры и степени панмиксии;
o единство происхождения. Естественно, и популяция и этнос не возникают "из ниоткуда", их развитие динамично. Имея материалы, относящиеся к разным этапам существования этих объединений, историк, археолог, этнограф, лингвист и антрополог (а лучше, если все они вместе) могут исследовать эту динамику - популяционную и этническую историю. Основной метод такого исследования очень сходен - это знакомое нам хронологическое и географическое сравнение выборок. Вся разница в том, что популяция исследуется по системам биологических признаков, а этнос - по признакам языка, свидетельствам материальной и духовной культуры.
Благодаря более интенсивному внутригрупповому обмену информацией разного рода, единство ареала способствует возникновению и сохранению единообразия в чертах культуры (поддержание и развитие традиций) и генофонда (сохранению и передаче генетического материала). Последний во многом определяет фенотип, то есть, в конце концов, и антропологический облик населения. Как для этноса, так и для популяции возникшее таким образом равновесное состояние может нарушаться в силу исторических причин. Например, в результате заимствования (культурного обмена) или смешения населения с носителями иных языковых и прочих традиций (в результате включения в группу инородного в биологическом отношении компонента) и т.п. Вот и получается, что часто, говоря о популяции и об этносе, мы имеем дело с одной и той же группой населения. А это в свою очередь означает, что могут возникать устойчивые параллели (ассоциации) между биологическими характеристиками группы (антропологическими признаками) и этническими определителями.
Этническая антропология и её основные задачи. Рогинский Я.Я., Левин М.Г. "Антропология", 1978:
"Этническая антропология (расоведение) изучает антропологический состав народов земного шара в настоящем и прошлом. Полученные в результате этого изучения материалы дают возможность выяснить родственные отношения между расами и историю их возникновения, т.е. древность, место и причины происхождения расовых типов.
Этническая антропология обслуживает также интересы и смежных с ней дисциплин, разрешая при этом ряд различных задач:
Сравнительное изучение человеческих рас весьма важно для разрешения проблемы происхождения человека современного вида и в первую очередь для разрешения вопроса о том, в одном или в нескольких местах возник человеческий род. В истории науки этот вопрос всегда рассматривался и решался в неразрывной связи с вопросом о степе ни взаимного сходства и различия между человеческими расами, а также об изменчивости расовых типов под влиянием географической среды.
Этническая антропология представляет существенный материал для изучения закономерностей роста, изменчивости и корреляций, весьма важных для биолога.
Антропологический (расовый) материал имеет большое значение и для разрешения некоторых вопросов истории, в особенности вопросов этногенеза. Наличие на территории типа, резко отличного от основной массы населения, позволяет с уверенностью говорить о приходе на данную территорию либо предков основной массы населения, либо предков типа, составляющего меньшинство. Более того, в этнографических и археологических данных в подобных случаях всегда обнаруживаются различия в культуре в настоящем или в прошлом между группами, отличающимися друг от друга по антропологическому типу. Совокупными усилиями антрополога, этнографа, археолога, а иногда и лингвиста можно определить область, из которой шло переселение, и время этого переселения.
Приведем некоторые примеры использования антропологического материала как исторического источника.
Появление азиатских тюркских кочевников на территории южной Украины сопровождалось отчетливым изменением физического типа, о чем свидетельствуют, например, скелетные останки, добытые на татарском кладбище XV в. в Тягинке близ Херсона.
Исследование черепов из Луговского могильника ананьинской культуры показало преобладание у ананьинцев монголоидных черт и явилось свидетельством того, что древнее население этой культуры было связано с лесными областями Зауралья (Трофимова). Это положение нашло свое подтверждение и в археологических материалах.
С другой стороны, антропологический анализ заставляет в некоторых случаях отвергнуть гипотезу о массовом переселении. Так, почти полное отсутствие монголоидных черт в тюркском по языку населении Азербайджана вынуждает отказаться от предположения, что тюркский язык распространился в восточном Закавказье в результате вторжения в эту область больших групп завоевателей монгольского расового типа.
Таким образом, изучение человеческих рас может оказать некоторую помощь истории. <...> (с. 339—340).
Изучение рас представляет интерес для историка и помимо проблемы переселений. По степени расчлененности на типы можно до известной степени заключить как о давности пребывания на территории тех или иных групп, так и о степени их изоляции. Так, сравнение коренного населения Америки с населением Старого Света с точки зрения разнообразия их расового состава заставляет сделать вывод, что разделение на расовые типы в Америке далеко не достигло той степени резкости, какую мы находим в Старом Свете. Этот антропологический факт представляет собой существенное подтверждение вывода, основанного на данных археологии и палеоантропологии, что Америка была заселена сравнительно поздно.
Очевидно, что вопросы о переселении и изоляции народов имеют близкое отношение к проблеме их образования и расчленения, т.е. к проблеме этногенеза. Примером использования антропологических данных для разрешения этой проблемы могут послужить исследования о происхождении народов угрофинской языковой группы.
По этнографическим материалам прослеживаются три основных направления связей угрофинских народов: первое направление ведет к историко-этнографической области 06скоЕнисейского бассейна с ее культурой лесных охотников и рыболовов; второе — связывает угрофинские народы с Волго-Балтийской историко-этнографической областью, с ее культурой земледельцев, первоначально мотыжных, а с середины I тысячелетия н.э. плужных, наконец, у многих различных финоязычных народов Волго-Камья прослеживаются связи со скотоводческими культурами причерноморских и аралокаспийских степей. Данные расоведения позволяют выделить у современных угрофинских народов как раз три группы антропологических типов, одна из которых — уральская (смешанная монголоидноевропеоидная) связана с Западной Сибирью, вторая — светлая европеоидная тяготеет к Прибалтике, третья — темноволосая европеоидная свидетельствует о былой их близости к странам Причерноморья. Таким образом, народы угрофинской языковой группы складывались на основе населения различных историко-этнографических областей, причем каждая из них характеризовалась и характеризуется определенной группой антропологических типов (Чебоксаров, 1951)" (с. 340).
16. Классификация рас по Деникеру
Деникер создал классификацию человеческих рас (1900), в которой впервые был последовательно проведён принцип выделения антропологических типов исключительно по физическим признакам (см. ниже). Серьёзным недостатком этой классификации являлось отсутствие исторического подхода, что не давало возможности проследить процесс возникновения рас. Некоторые принципы классификации Деникера используются и в настоящее время. Деникер также впервые создал карты расового состава Европы. И.Е.Деникер — разделил расы на шесть основных и более чем на двадцать дополнительных рас.
Система расовой классификации Деникера
Карта"Races de l’Europe" (РасыЕвропы), 1899.

Деникер выделял 6 расовых стволов:
· группа А (шерстовидные волосы, широкий нос): бушменская, негритосская, негрская и меланезийская расы;
· группа B (курчавые или волнистые волосы): эфиопская, австралийская, дравидийская и ассироидная расы;
· группа C (волнистые, тёмные или чёрные волосы и тёмные глаза): индоафганская, арабская или семитская, берберская, южноевропейская, иберо-островная, западноевропейская и адриатическая расы;
· группа D (волнистые или прямые волосы, блондины со светлыми глазами): североевропейская (нордическая) и восточноевропейская расы;
· группа E (прямые или волнистые, чёрные волосы, тёмные глаза): айносская, полинезийская, индонезийская и южноамериканская, североамериканская, среднеамериканская, патагонская расы;
· группа F (прямые волосы): эскимосская, лопарская, угорская, турко-татарская и монгольская расы.
Среди европейских рас, кроме вышеупомянутых, Деникер выделял определённые подрасы:
· северо-западную;
· субнордическую;
· субадриатическую;
· вистульскую (привислянскую) или восточную.[3]
В ходе дальнейших исследований почти все выделенные Деникером расы получили подтверждение как реальные единицы расовой систематики [1]
.
Деникер ввёл в научный оборот термин "нордическая раса" (la race nordique, "северная раса").
17. Правила Бергмана, Аллена, Глогера, Томсона-Бакстона
Правила Бергмана, Аллена, Глогера, Томсона-Бакстона
Правило Бергмана - Отражает зависимость размеров тела от температуры среды
Правило Алена - Устанавливает связь с климатом не размеров тела, а его пропорций
Правило Глогера - Выражает принцип: чем ближе к тропикам, тем интенсивнее окраска
Правило Томсона-Бакстона - Выявляет зависимость ширины носа от климатических показателей (максимальная ширина носа в тропиках)
Адаптация и человек
"Примеры адаптивной изменчивости человека очень многочисленны. В первую очередь возникает вопрос о возможности распространения на человека "экологических правил", установленных для животных. После длительной дискуссии на эту тему его можно считать решенным положительно. Более того, помимо экологических правил Аллена, Бергмана и Глогера, для человека было установлено новое правило Томсона и Бакстона, выражающее зависимость ширины носа от климатических показателей.
Согласно правилу Бергмана, наблюдается зависимость между температурой среды и размерами тела гомотермных животных: в районах сурового арктического и антарктического климата размеры тела больше, чем в тропической зоне.
Применительно к популяциям человека аналогичное соотношение продемонстрировано сразу для нескольких признаков: длины, веса и поверхности тела. Каждый из этих признаков в отдельности может служить показателем общих размеров тела.
Межгрупповые корреляции этих показателей со среднегодовыми температурами обнаруживают закономерный характер и высокую степень связи между ними: и длина тела, и его вес связаны со средней годовой температурой отрицательной корреляцией. Из этого следует, что в среднем малорослые группы людей встречаются чаще в центральном поясе ойкумены, чем на ее северных и южных окраинах. А это как раз и есть явление, которое постулируется правилом Бергмана.
Правило Аллена трактует связь с климатом не размеров, а пропорций тела гомотермных животных. В холодном климате они имеют укороченные конечности и отличаются более плотным сложением.
У человека сопоставление измерений, характеризующих пропорции тела с климатическими характеристиками обнаруживает довольно тесную связь между ними наподобие той, которая существует у животных. А именно: группы людей, которые проживают в тропическом климате, отличаются в среднем удлиненными пропорциями и менее плотным сложением, проживающие в умеренном и холодном поясах — более плотным сложением и укороченными пропорциями тела.
Правилом Глогера устанавливается интенсивность окраски животных в зависимости от широты местности: чем ближе к тропикам, тем окраска интенсивнее.
То же соотношение характерно и для человека. Цвет кожи, волос и глаз закономерно светлее по мере перехода от тропического пояса к умеренной зоне в обоих полушариях и затем еще более светлеет при переходе к арктической зоне. Отклонения от этого правила — а оно, так же как и предыдущие, имеет статистический характер и может быть установлено не при попарном сопоставлении групп людей из тропического и, скажем, умеренного пояса, а лишь на большом материале, отражающем изменчивость пигментации в десятках популяций, — легко объяснить поздними переселениями людей из другой климатической и ландшафтной зоны (эскимосы в Гренландии и на Аляске, лопари в Скандинавии и т.д.).
Любопытно отметить, что наиболее тесная связь пигментации с климатическими факторами в зависимости от широтной зональности наблюдается в Европе и Америке, где эта зональность имеет наиболее закономерный характер и где расселение народов в наибольшей мере отражало этапы первоначального (в палеолите) заселения территории человеком современного вида.
Правило Томсона — Бакстона привлекает особое внимание потому, что оно устанавливает зависимость от климата такой особенности, которая практически слабо развита у животных и составляет отличительную принадлежность человеческого лица; Зависимость вариаций ширины носа от интенсивности среднегодовой температуры и солнечной радиации, от широты местности также статистическая, как и во всех предыдущих случаях. Максимальная ширина носа в среднем характеризует те группы человечества, которые расселены в тропической зоне, минимальные величины зафиксированы у населения Скандинавии, северо-восточной оконечности Азии, Аляски, Гренландии и Огненной Земли. Наиболее четко эта зависимость представлена, как и в случае с пигментацией, на Американском континенте.
Климатические правила, правила, отражающие закономерности географич. изменчивости организмов, связанной с климатич. градиентами. К основным К. п. относятся правила Бергмана, Аллена, Глогера и др. Тот факт, что географич. изменчивость в значительной мере коррелирует с климатич. изменениями, доказывает, что разные виды могут сходным образом реагировать на воздействие одного и того же фактора среды. Эти различия имеют генетич. основу, следовательно, экогеографич. правила доказывают селективную роль среды. Многообразие в значительной степени независимых закономерностей свидетельствует о многообразии селективных компонентов среды. Постепенность градиентов признаков, обусловленная действием климатич. правил, показывает, что географич. изменения фенотипа есть следствие взаимодействия многочисленных генетич. факторов, каждый из к-рых создает небольшой фенотипич. эффект.
До демографического скачка в Европе и освоения Америки собственно „бледнолицые" составляли ничтожный процент человечества. Географический центр потери пигмента — побережье Балтийского моря, так называемый пояс блондинов. Несколько шире располагается „пояс голубых глаз", а ещё шире — „пояс светлокожих". Далее простирается сплошной массив весьма смуглых европеоидов (марокканцев, египтян, арабов, персов, индийцев). Многие антропологи присоединяли к этому ряду эфиопов, обладающих утончёнными и европейскими чертами лица. Если принять их точку зрения, окажется, что некоторым европеоидам присуща… самая тёмная кожа на Земле.
Причина огромного разнообразия этой расы в контрастах её ареала — от тропиков до Арктики. При движении с юга на север на европеоидов действуют общие правила экологии: Бергмана (увеличиваются размеры тела), Аллена (укорачиваются конечности), Глогера (падает интенсивность окраски). Для иллюстрации сравните, например, рост, пропорции и цвет лица бенгальского крестьянина и беломорского моряка.
Итак, европеоиды — не „белые люди". Как же тогда их охарактеризовать? В первую очередь, в глаза бросается отсутствие черт, присущих другим расам, — монгольской складки века (эпикантуса), курчавых волос, толстых губ. А что же присутствует? Пожалуй, рельеф лица: вперёд выдаются как его центр, так и „детали", — скулы, надбровье, носовые кости, подбородок. Отчасти это связано с тем, что европеоидные черты формировались в горных регионах с довольно суровым климатом. В частности, увеличенная полость носа лучше защищала дыхательные пути от холодного и сухого воздуха.
Как вытекает из ряда исследований, размеры и пропорции человеческого тела, а также состав его по основным компонентам подчиняются известным экологическим правилам Бергмана и Аллена, согласно которым в пределах одного вида, рода и семейства в тропическом поясе сосредоточены преимущественно некрупные формы с удлиненными конечностями, тогда как по направлению к полюсам увеличиваются размеры и уменьшается длина конечностей. Вес тела, его поверхность и отношение длины ног к длине туловища обнаруживают высокую корреляцию с климатической зональностью: вес последовательно увеличивается к полюсам, то же происходит и с поверхностью тела, хотя последняя зависимость менее отчетлива (она видна яснее, если поверхность соотносится с весом), длина же ног, наоборот, уменьшается к полюсам. По соотношению костной и мускульной тканей в разных популяциях пока нет достаточно убедительных данных; что же касается жировой ткани, то ее количество повышается в полярных популяциях.
Экологическое правило Томсона — Бакстона, названное по имени исследователей, впервые открывших эту зависимость, и постулирующее положительную связь ширины носа с влажностью и температурой (максимально - в тропических широтах).
18. Классификация рас по группам крови по Бойду
На основании результатов изучения антигенов крови (серологических различий) У.Бойдом в 1953 году было выделено в человечестве шесть рас, а именно:
1. Европейская группа, включающая лопарей, южных европейцев и северных африканцев (европеоиды, лапаноиды, средиземноморцы). 2. Африканская группа (негроиды Африки). 8. Азиатская группа, включая жителей индийского субконтинента (монголоиды Азии и индо-дравийская раса). 4. Американская группа, включая всех аборигенов континента. 5. Тихоокеанская группа (представители меланезийской, индонезийской полинезийской групп).
6. Австралийская – австралийцы.
Несмотря на наличие современных научных классификаций, в наше время очень широко распространено подразделение человечества на европеоидов, негроидов, монголоидов и австралоидов.
У индейцев Америки нет группы крови В и А, они являются носителями группы О. В Центральной Азии распространена группа В. В Индии, Китае, Африке лица с группой крови А очень редки. Люди с группой крови В устойчивы к чуме и оспе.
19. Природные факторы расообразования
Природные факторы расообразования
Природные факторы классифицируются на 3 группы:
1. Космические факторы воздействуют на наследственную изменчивость человека, вызывают мутацию ДНК
2. Физико-географические агенты выражают соотношение балансов тепла и влаги, индекс сухости и т.д.
3. Адаптация человека к биосфере планеты как защитная реакция против вирусных и микробных инфекций
Географическая изменчивость является основой расообразования. Согласно популяционной концепции раса есть совокупность популяций.
Согласно типологической концепции раса есть сумма индивидуумов.
"Какова роль природных факторов расообразования? Специалисты сопоставляли географические вариации тех или иных признаков с климатическими характеристиками. В результате получены убедительные положительные корреляции между шириной носа и среднегодовой температурой, высокие отрицательные — между среднегодовой температурой и признаками, характеризующими развитие тела в ширину. Все эти корреляции связаны с физиологией человека. Ширина носа явно определяет в числе других признаков уровень дыхания, и поэтому широкий нос способствует теплоотдаче. Очень трудно сравнивать толщину губ у разных рас в зависимости от географических вариаций температуры и влажности. Но все же следует помнить, что у представителей негроидной расы наиболее толстые губы, и они также способствуют терморегуляции. Удлиненные пропорции тела при малом весе аналогичным образом благоприятны в жарком влажном климате" (с. 18—19).
Алексеев В.П. "География человеческих рас", 1974:
"Для целей исследования и показа адаптивности расообразования целесообразно рассматривать три самостоятельные группы расообразующих факторов — космические, физико-географические и биотические. Их удельный вес, разумеется, неодинаков, но все они представляли собой в расообразовании самостоятельные области влияний и определяли на разных этапах эволюции семейства гоминид магистральную линию и характер расообразовательного процесса <...> (с. 27).
Представляется целесообразным специальное выделение совокупности космических факторов. Прозорливые прогнозы В.И. Вернадского (1926) о том, что вся биосфера буквально пронизана излучениями космического происхождения, получили многообразное подтверждение в последние десятилетия. Собрана огромная информация о влиянии этих излучений и на человека. В расообразовании их роль, как мы убедимся дальше, проблематична, хотя и вероятна. Их взаимодействие на наследственную изменчивость человека, если оно будет строго доказано, носит, очевидно, мутагенный характер и относительно свободно, во всяком случае на первом этапе, от влияния селективных механизмов. Все это наряду с локализацией самих источников космических излучений и оправдывает выделение космических факторов расообразования в особую группу <...> (с. 26).
Физические агенты — свет, температура, гравитация, электромагнитные явления в околоземном пространстве, радиоактивные вещества. Каждый из этих агентов воздействует на важнейшие жизненные функции, вызывая в избыточном количестве их угнетение и приводя к патологии. Теоретически говоря, можно полагать, что их колебания в нормальных пределах также небезразличны для организма, стимулируя одни системы органов, угнетая другие, вызывая развитие дополнительных адаптации. А это и есть географическая изменчивость — основа расообразования.
Свет и тепло, получаемые поверхностью Земли от Солнца, не могут быть четко дифференцированы в своем влиянии на расообразование. Поэтому в комплексах признаков, характерных для населения тропических районов и арктических широт, трудно назвать специальные приспособления отдельно к интенсивной инсоляции и активному или замедленному теплообмену. Такие приспособления составляют неделимое сочетание, и их нужно оценивать как целое (общие вопросы влияния тепла и света на человеческий организм рассмотрены: Чубинский, 1959, 1965). Они изучены полнее, чем приспособления к другим физическим агентам" (с. 28—29). Алексеев В.П. "Новые споры о старых проблемах", 1991:
"Помимо температуры и влажности, в список важных природных факторов расообразования можно включить интенсивность солнечного облучения, содержание кислорода в воздухе, некоторые почвенные агенты.
Солнечное облучение исключительно интенсивно в тропиках, и высокое содержание меланина в покровных слоях кожи предохраняет ее от ожогов. Недостаточность кислорода на больших высотах приводит к комплексу приспособлений к высокогорной гипоксии, обнаруженной у живущих на большой высоте над уровнем моря народов в Андах, на Тянь-Шане, в предгорьях Гималаев. Это большая экскурсия грудной клетки, разреженный пульс, значительный объем легких, пониженный обмен и т.д. Есть небезоговорочное утверждение, что проживание на базальтовых почвах способствует брахикефализации. По-видимому, минеральный состав гранитов и базальтов сильно влияет на состав вод соответствующего района. И видимо, содержащиеся в воде минералы влияют на соотношение продольного и поперечного диаметров головы в процессе роста, что и служит непосредственной морфогенетической причиной брахикефализации" (с. 20).
Алексеев В.П. "География человеческих рас", 1974:
"Человек зависим физиологически от электромагнитных явлений в атмосфере и приземном слое пространства. Замечательные исследования А.Л. Чижевского (библиографию его работ см.: Чижевский, 1964) открыли новую главу в биофизике, и в частности в антропофизике, показав целый комплекс связей между физиологическими реакциями человека и электромагнитным состоянием окружающей его среды. Особое значение он придавал ионизации воздуха, чему посвятил специальную монографию (Чижевский, 1960). А.Л. Чижевский продемонстрировал возникновение разнообразных патологий в зависимости от состояния атмосферы. Но атмосфера подчиняется закону географической зональности, меняется в результате изменений солнечной активности и неодинаковой интенсивности солнечного облучения, следовательно, ее влияние на организм человека не может не носить дифференцированного локального характера. Отсюда можно ожидать возникновения специфических физиологических приспособлений, наверное, можно истолковать и какие-то детали формы под этим углом зрения. В этой области предстоит еще непочатый край исследований.
Мутагенный эффект радиоактивных веществ общеизвестен. В высокой концентрации они приносят непоправимый вред всему живому. Но в то же время естественный фон радиоактивности есть постоянный фон жизни, в том числе и эволюции человека. Концентрация радиоактивных веществ заведомо непостоянна в разных районах, выделяются очаги малой и большой радиоактивности. Пока не замечено каких-либо правильностей в формообразовании, приуроченных к этим очагам, но наличие таких правильностей вероятно. Тогда в расообразование вводится мощный формообразующий фактор, значение которого, однако, нельзя сейчас учесть сколько-нибудь точным образом" (с. 33).
Алексеев В.П. "Новые споры о старых проблемах", 1991:
"Биотические факторы, в свою очередь, подразделяются на прямые воздействия биоты и то, что можно назвать трансмиссивной, т.е. передаточной функцией биосферы. Классический пример прямого воздействия биоты на географическую изменчивость человека — география аномальных гемоглобинов. Вызываемые ими гемоглобинопатии приурочены к областям с широким распространением малярийных паразитов и тропической малярии. Это и послужило поводом к формулировке сейчас все более подтверждаемой гипотезы об аномальных гемоглобинах, в которых не может нормально развиваться малярийный паразит. Эта аномалия сформировалась как адаптивный признак в условиях сильного распространения малярии" (с. 19).
Алексеев В.П. "География человеческих рас", 1974:
"Формообразующая роль пищевого рациона сводится к его количеству и составу. Эмпирические наблюдения давно выявили эту роль для отдельных групп, находящихся в экстремальных условиях существования. Исключительное развитие нижней челюсти у ительменов (мировой максимум нижнечелюстной ширины) ставят в связь с преимущественным употреблением в пищу сырой рыбы (Дебец, 1951). Пища в данном случае выступает в качестве механической причины появления признака, возникшего, разумеется, не вследствие прямого приспособления, а в результате селекции. Аналогичным образом объясняется происхождение отличительных физических особенностей пигмеев тропических лесов Африки и Юго-Восточной Азии — малый рост и миниатюрное сложение образовались как приспособительная реакция и селективное преимущество при многовековом недостатке пищи. Именно поэтому все большее число специалистов высказывается сейчас против старой теории о генетическом родстве всех народов карликового роста, тем более что в морфологии и физиологии между ними много различий — малые размеры тела и миниатюрное сложение легко могли возникнуть конвергентно. Многократные подтверждения этого мы видим в понижении роста вслед за многолетним голодом или длительной войной. Дополнительным фактором грацилизации, т.е. уменьшения массивности скелета по мере приближения к современной эпохе, послужило, очевидно, приспособление к характерному для тропического леса недостатку кальция и фосфора (Marrett, 1936) <...> (с. 38—39).
Многовековое воздействие стандартных пищевых рационов сыграло не последнюю роль в расообразовании, обусловив разницу между группами популяций не только в размерах тела и уровне жирового обмена, непосредственно реагирующих на избыток и недостаток пищи, но частично и в других признаках. Забегая вперед, укажу в качестве доказательства на совпадение очагов расообразования у человека с центрами происхождения культурных растений <...>.
Совпадение их могло образоваться за счет того, что оба процесса — расообразование и введение растений в культуру — протекали на совпадающих географических территориях и поэтому отражали географическую дифференциацию исторического процесса. Не исключено, однако, что некоторые морфологические признаки, свойственные перечисленным расам, образовались вследствие приспособления к условиям местного пищевого режима, в свою очередь отражающего конкретные результаты культурного преобразования данного участка биосферы.
Прямое доказательство непосредственной реакции интенсивности роста и размеров тела на полноценность питания, особенно в отношении содержания белка, получено неоднократно (уменьшение роста у средневековых предков исландцев под влиянием векового голода)" (с. 41—42).
Алексеев В.П. "Новые споры о старых проблемах", 1991:
"В чем заключается трансмиссивная функция биосферы? В передаче влияний косной природы на организм человека. Чаще всего она проявляется в опосредованном биосферой влиянии геохимической ситуации на расообразование. Наличие тех или иных микроэлементов способствует развитию отдельных признаков, отсутствие их угнетает эти признаки. Правда, тот или иной состав микроэлементов передается человеку и через воду, но больше всего все же за это ответственна пища растительного и животного происхождения, которая чаще всего непосредственно связана с местной биотой. Поэтому существует положительная корреляция размеров головы и лица с содержанием кальция. Кальций способствует развитию скелета, а на кальциевых почвах, как правило, мы застаем население с большими размерами лица и головы. <...>
Я должен подчеркнуть, что ни в коем случае нельзя оценивать сказанное как утверждение о прямом влиянии среды на признак и адекватной реакции организма на среду. Антропология, как и другие области биологии, не располагает однозначными данными о прямом влиянии среды на признак и передаче приобретенных особенностей по наследству. Что подразумевается, когда я говорю о прямом пути влияния природных условий на расообразование? То, что географические условия влияют не через биоту и биотические условия выступают не в роли трансмиссивной функции, а в роли непосредственных расообразующих агентов. В то же время во всех рассмотренных случаях адекватность географических вариаций тех или иных признаков и географических условий можно истолковать сколько-нибудь разумным образом, только допустив селекцию благоприятных вариаций признаков, т.е. действие отбора. Он сильнее проявляется на ранних этапах расообразования и в экстремальных условиях и слабее — на поздних его этапах.
Конечно, отбор признаков и их комбинаций, благоприятных в определенных условиях географической среды, не был единственной движущей силой расообразования. Ведьтогда расообразование у человека свелось бы к географической адаптации. Между тем за многими признаками расового комплекса трудно увидеть географическое приспособление. Таковы подавляющая часть генетических признаков, обильное развитие волосяного покрова у некоторых рас, сильный прогнатизм негров и т.д. Поэтому как ни велика роль адаптации к географической среде в расообразовании, они не единственные.
В расообразовательном процессе огромное значение имеют социальные адаптации, а также популяционно-генетические механизмы. Иногда они усиливали приспособление к среде, но чаще всего действовали самостоятельно, затушевывая роль непосредственных географических адаптации. Поэтому все отмеченные выше приспособления к географической среде довольно четко выявляются при рассмотрении накопленной информации в пределах всего земного шара. И лишь иногда с большим трудом прослеживаются при исследовании отдельных популяций, а порой они и вовсе не прослеживаются" (с. 19—21).
20. Гипотеза адаптивных типов в расогенезе
Гипотеза адаптивных типов по Т.И.Алексеевой (1986 г.)
Адаптивные типы человека отражают специфические нормы реакции на определенные природные факторы, разные формы адаптации к условиям среды.
Арктический адаптивный тип (чукчи, эскимосы, саамы, лесные ненцы) появился как защитная реакция на влажный и холодный климат. У его представителей наблюдаются усиленный газообмен, повышенный уровень холестерина и иммунных белков в сыворотке крови, усиленная минерализация скелета.
Предгорный адаптивный тип (предгорные таджики) возник от недостатка кислорода. У них пониженный газообмен, обезжиренная масса тела, низкий уровень холестерина и иммунных белков в крови, сильно минерализированный скелет.
Континентальный адаптивный тип (буряты) с характерным уменьшением удельного веса массы тела, значительным жироотложением, слабой минерализацией скелета, более высокой, по сравнению с жителями умеренных районов, теплопродукцией, но одинаковым уровнем холестерина в крови.
Экваториальный адаптивный тип (бушмены, банту, индийские народы) появился в результате адаптации к жаре. У его представителей наблюдаются: усиленная теплоотдача и пониженный уровень энергетических процессов, повышенный уровень иммуноглобулинов и пониженный уровень холестерина в крови; найден белок трансферин, регулирующий температурный режим тела.
Адаптация и человек
"Примеры адаптивной изменчивости человека очень многочисленны. В первую очередь возникает вопрос о возможности распространения на человека "экологических правил", установленных для животных. После длительной дискуссии на эту тему его можно считать решенным положительно. Более того, помимо экологических правил Аллена, Бергмана и Глогера, для человека было установлено новое правило Томсона и Бакстона, выражающее зависимость ширины носа от климатических показателей.
Согласно правилу Бергмана, наблюдается зависимость между температурой среды и размерами тела гомотермных животных: в районах сурового арктического и антарктического климата размеры тела больше, чем в тропической зоне.
Применительно к популяциям человека аналогичное соотношение продемонстрировано сразу для нескольких признаков: длины, веса и поверхности тела. Каждый из этих признаков в отдельности может служить показателем общих размеров тела.
Межгрупповые корреляции этих показателей со среднегодовыми температурами обнаруживают закономерный характер и высокую степень связи между ними: и длина тела, и его вес связаны со средней годовой температурой отрицательной корреляцией. Из этого следует, что в среднем малорослые группы людей встречаются чаще в центральном поясе ойкумены, чем на ее северных и южных окраинах. А это как раз и есть явление, которое постулируется правилом Бергмана.
Правило Аллена трактует связь с климатом не размеров, а пропорций тела гомотермных животных. В холодном климате они имеют укороченные конечности и отличаются более плотным сложением.
У человека сопоставление измерений, характеризующих пропорции тела с климатическими характеристиками обнаруживает довольно тесную связь между ними наподобие той, которая существует у животных. А именно: группы людей, которые проживают в тропическом климате, отличаются в среднем удлиненными пропорциями и менее плотным сложением, проживающие в умеренном и холодном поясах — более плотным сложением и укороченными пропорциями тела.
Правилом Глогера устанавливается интенсивность окраски животных в зависимости от широты местности: чем ближе к тропикам, тем окраска интенсивнее.
То же соотношение характерно и для человека. Цвет кожи, волос и глаз закономерно светлее по мере перехода от тропического пояса к умеренной зоне в обоих полушариях и затем еще более светлеет при переходе к арктической зоне. Отклонения от этого правила — а оно, так же как и предыдущие, имеет статистический характер и может быть установлено не при попарном сопоставлении групп людей из тропического и, скажем, умеренного пояса, а лишь на большом материале, отражающем изменчивость пигментации в десятках популяций, — легко объяснить поздними переселениями людей из другой климатической и ландшафтной зоны (эскимосы в Гренландии и на Аляске, лопари в Скандинавии и т.д.).
Любопытно отметить, что наиболее тесная связь пигментации с климатическими факторами в зависимости от широтной зональности наблюдается в Европе и Америке, где эта зональность имеет наиболее закономерный характер и где расселение народов в наибольшей мере отражало этапы первоначального (в палеолите) заселения территории человеком современного вида.
Правило Томсона — Бакстона привлекает особое внимание потому, что оно устанавливает зависимость от климата такой особенности, которая практически слабо развита у животных и составляет отличительную принадлежность человеческого лица; Зависимость вариаций ширины носа от интенсивности среднегодовой температуры и солнечной радиации, от широты местности также статистическая, как и во всех предыдущих случаях. Максимальная ширина носа в среднем характеризует те группы человечества, которые расселены в тропической зоне, минимальные величины зафиксированы у населения Скандинавии, северо-восточной оконечности Азии, Аляски, Гренландии и Огненной Земли. Наиболее четко эта зависимость представлена, как и в случае с пигментацией, на Американском континенте.
Наконец, неоднократно указывалось и на приспособительное значение некоторых других признаков: курчавоволосости негров и характерной для населения тропической зоны удлиненной высокой черепной коробки, уплощенности лица и строения глазной области монголоидов, необычайно узкого носа эскимосов. Физиологическим признакам в этой связи уделялось мало внимания, хотя некоторые факты его заслуживают. К их числу относится разная скорость кровотока при охлаждении у эскимосов и лиц европейского происхождения, проживающих в Арктике. Скорость кровотока у эскимосов при понижении температуры примерно вдвое больше, чем у европейцев, что помогает эскимосам значительно легче, чем европейцам, сохранить тепловой баланс организма на определенном уровне и оберегает от переохлаждения. Установлены приспособительные изменения в величине легких и интенсивности кислородного обмена у народов северных районов Анд, проживающих на больших высотах.
Не менее важны в связи с проблемой физиологической адаптации наблюдения, иллюстрирующие приспособительный характер вариаций групповых факторов крови. Таких наблюдений, концентрирующихся, правда, вокруг системы АВО, накопилось к настоящему времени довольно много. Они говорят об избирательном характере разных заболеваний в зависимости от той или иной группы крови, о разном предрасположении носителей разных групп крови системы АВО к болезням, обусловленным групповой принадлежностью. Это свидетельствует о том, что избирательная реакция на воздействия среды, в том числе и на заболевания, вероятна и для других эритроцитарных систем крови и вообще для многих биохимических свойств, что сразу же намного расширяет сферу адаптивной изменчивости человека.
По мнению Л.А. Зильбера, собравшего много данных об иммунитете представителей разных рас к различным заболеваниям, он примерно одинаков, но приводимые им сведения противоречат этому заключению. Они скорее указывают на разную иммунологическую реактивность организма представителей различных рас. Например, негроиды резистентны ко многим инфекционным заболеваниям, распространенным в тропической зоне, и, наоборот, весьма восприимчивы к новым инфекциям, принесенным европейцами или встречающим негров при переселении за пределы Африканского материка. Общеизвестны опустошения, произведенные среди различных народностей, не соприкасавшихся с цивилизацией, болезнями, которые принесли европейцы.
Перечисленные факты свидетельствуют, что адаптивный фактор играл и играет большую роль в формообразовании у человека и что адаптационные процессы у представителей разных расовых и конституциональных типов имеют определенную специфику. Возникает закономерный вопрос: наследственны ли все перечисленные выше морфологические признаки и физиологические реакции, могут ли они действительно служить для анализа процесса адаптации человека?
Для ответа можно использовать факты, почерпнутые непосредственно из генетических исследований, и косвенные соображения, основанные на рассмотрении времени и этапов формирования всех перечисленных особенностей человеческого организма.
Особенно показательны, а главное, точно изучены факты, относящиеся к наследуемости групп крови и генетических маркеров кровяной сыворотки. Подавляющая часть до сих пор изученных групповых антигенов у человека управляется в своей наследственной передаче двумя или тремя аллеломорфными генами. Никаких данных, свидетельствующих о значительном темпе мутирования этих генов или возникновении их в недавнее время, нет, наоборот, есть все основания считать их древними образованиями, тем более что аналогичные гены открыты и у человекообразных обезьян.
В каком порядке и последовательности они возникли — все сразу или по отдельности, представляют ли они собой действительно аллели одного гена или систему разных генов — все это совершенно неясно и высказываемые предположения не выходят за рамки рабочих гипотез. Но для нас важна строгая наследственная обусловленность групп крови и белковых фракций сыворотки, в том числе и тех, длякоторых отмечена связь с наклонностью к определенным заболеваниям.
Генетическое отличие морфологических признаков от физиологических заключается, согласно наиболее распространенной гипотезе, в гораздо большем числе управляющих их наследованием генов. Такие морфологические признаки с непрерывной трансгрессивной изменчивостью наследуются количественно, т.е. зависят в своей наследственной обусловленности от многих наследственных факторов. Это справедливо, по-видимому, и для размеров головы и тела и их соотношений, и для пигментации.
Все перечисленные признаки менее автономны, более обусловлены в своем развитии влияниями среды, чем физиологические особенности, но их генетический характер не вызывает сомнений и доказывается многими экспериментальными работами. Сводка данных, правда уже несколько устаревшая, приведена Р. Гейтсом и В.П. Эфроимсоном. Однако о передаче по наследству особенностей морфологического типа и пигментации если и не в отдельных индивидуальных случаях, то от поколения к поколению в целом можно судить не только по прямым генетическим наблюдениям, но и на основании косвенных данных.
Первое из них — исключительное постоянство физического типа человека во времени, зафиксированное многими палеоантропологическими исследованиями. Эта стабильность типа характерна не для веков или тысячелетия, а для нескольких тысячелетий. Такая стабильность основных особенностей строения лица и тела охватывает приблизительно 7 тысячелетий на территории Египта и Нубии, 5—6 тысячелетий в Северном Китае, 3—4 тысячелетия на территории Армении. В Египте можно проследить и преемственность типичной интенсивности пигментации, начиная с эпохи Древнего царства, поскольку на цветных фресках изображено не только население Древнего Египта, но и народы, платившие Египту дань, во всей конкретности их антропологических черт.
Второе свидетельство наследственной обусловленности морфологического типа — промежуточное положение антропологических особенностей народов смешанного происхождения между исходными группами. Это народы Западной Сибири и Казахстана, частично Средней Азии, Северной Африки, Эфиопии и Пиренейского полуострова. Аналогичные данные собраны и для народов, образовавшихся уже в историческое время как результат позднего смешения европейцев с населением других материков. Итак, все перечисленные особенности человеческого организма, в которых выявлено приспособление к условиям среды в широком смысле оказываются в той или иной мере наследственными. Следовательно, речь идет о действительно адаптивных свойствах человеческого организма, претерпевших под влиянием и в процессе приспособления к среде глубокую внутреннюю перестройку, затрагивающую и гены, а не о кратковременных фенотипических сдвигах акклиматизационного характера" (с. 63—67).
Происхождение адаптивных экологических типов
Человечество возникло в тропической зоне Африки, и следует полагать, что поэтому наиболее древним и исходным для остальных экологических типов человека является тропический тип. Об этом свидетельствует и наибольший полиморфизм морфофункциональных признаков человека в рамках этого типа на территории Центральной и Северо-Восточной Африки. В связи с этим уместно обратиться к учению Н.И. Вавилова о центрах происхождения и многообразия культурных растений, в соответствии с которым именно в зонах первоначального вовлечения биологических видов в социальную среду наблюдается наиболее выраженный наследственный полиморфизм местных популяций по большому количеству признаков.
Об этом свидетельствует и то, что именно в тропической Африке в непосредственной близости друг от друга, т.е. под действием сходных экологических факторов, обитают наиболее низкорослые и наиболее высокорослые племена, например пигмеи, готтентоты и бушмены, с одной стороны, и массаи — с другой. Пределы изменчивости африканских популяций, например, по росту и массе тела таковы, что в них укладываются все известные на Земле человеческие популяции. Основные же черты тропического типа в Африке, отмеченные выше, остаются неизменными. Они же характеризуют монголоидные популяции Индокитая, Малайского архипелага и некоторые группы индейских племен зоны влажных тропических лесов Центральной и Южной Америки.
Это указывает на то, что тропический адаптивный тип развивается в результате асинхронного параллелизма в эволюции человека, причем вначале на африканском континенте, а потом и в других областях (см. раздел 3.13). Отсюда следует и еще один вывод: адаптивный тип формируется на фоне расогенеза и вне зависимости от него. В процессе адаптогенеза в популяциях человека можно проследить и проявление закона гомологических рядов. Это выражается, например, в том, что в бассейне Меконга в Индокитае и на острове Суматра имеются пигмеоидные популяции, по антропометрическим признакам соответствующие африканским пигмеям.
Экологический тип умеренного пояса сформировался на базе исходного генетического и фенотипического полиморфизма тропического типа при расселении популяций человека в умеренных зонах Евразии и позже — Северной Америки. Он оформился в рамках двух больших рас: европеоидной и монголоидной.
При заселении человеком арктической зоны в Евразии и Северной Америке произошло формирование арктического типа. Независимо от него среди индейцев Южной Патагонии и Огненной Земли в приантарктической зоне Южной Америки возникли популяции индейцев, по основному комплексу признаков соответствующие арктическому типу. Это еще один убедительный пример параллелизма эволюции человеческих популяций и реализации закона гомологических рядов, а также доказательство вторичности адаптивных типов по отношению к большим расам человечества.
Формирование горного адаптивного типа иллюстрирует общие закономерности адаптогенеза, отмеченные выше. Этот тип также развился независимо от расовой и этнической принадлежности популяций — среди европеоидов Альп, Кавказа, Памира и Гималаев, а также в монголоидных популяциях Тибета, Тянь-Шаня и Анд. В связи с тем что высокогорья заселялись человеком в последнюю очередь, горный экологический тип является по происхождению самым молодым. Интересно, что, несмотря на особенно выраженную расовую и этническую разнородность этого типа, комплекс основных признаков его является монолитным. Однако в отличие от других типов, вероятно, именно горный проявляется в основном только на фенотипическом уровне и не имеет наследственной природы. Об этом свидетельствует то, что количество эритроцитов в крови и объем грудной клетки людей, переселяющихся в условия высокогорья и обратно, могут меняться на протяжении жизни одного поколения.
Следовательно, адаптивные типы человека не только отражают его прошлое, но формируются и в настоящем, а разные типы имеют и разную по длительности историю.
В настоящее время пока еще на фенотипическом уровне идет формирование адаптивного типа человека городской среды, который характеризуется широкой лабильностью психических реакций, обеспечивающих способность переживать состояния постоянного стресса, и рядом морфофизиологических особенностей, оптимальных для жизни в специфических условиях города (см. разд. 17.3.2).
Формирование экологических типов человека в значительной степени обеспечило всесветное расселение людей. Меняющаяся среда обитания под действием антропогенных факторов ставит перед популяциями человека новые задачи, решаемые и сегодня за счет не только социальных, но и биологических адаптации. Схему эволюционных взаимоотношений больших рас и адаптивных типов человека см. на рис. 15.10.

Рис. 15.10. Адаптивные типы человека и большие расы
Антропоклиматические аналоги и адаптивные типы
Рассмотрим теперь некоторые явления, имеющие отношение и к зональному, и к азональному расообразованию. Что подразумевают под антропоклиматическими аналогами? Такие вариации антропологических признаков, которые образуют сходные или тождественные ряды изменчивости в одинаковых климатических условиях. В разделе о зональном расообразовании мы перечислили такие признаки — пигментация, ширина носа и другие.
Понятие адаптивного типа много сложнее и соотносится с понятием антропоклиматического аналога приблизительно так же, как соотносятся между собой признак и сочетание признаков (комплекс). Это понятие предложила Т.И. Алексеева для обозначения территориально приуроченных сочетаний бесспорно адаптивных признаков, т.е. таких сочетаний, которые возникли и сложились как приспособительные" (с. 29—35).
"Человеческие популяции проявляют определенную реактивную изменчивость в соответствии с влиянием окружающей их естественной среды. Судя по характеру морфофункциональных особенностей популяций, живущих в определенных географических условиях, эта реакция носит приспособительный характер, что и дает основание для введения в научные исследования понятия адаптивного типа, представляющего собой норму биологической реакции на комплекс условий окружающей среды, обеспечивающей состояние равновесия популяции с этой средой и находящей внешнее выражение в морфофункциональных особенностях популяции.
Адаптивный тип независим от расовой и этнической принадлежности. В одних и тех же геоклиматических условиях разные по происхождению группы имеют одно и то же направление приспособительных реакций, так же как в различных условиях обитания близкие в генетическом отношении группы демонстрируют различия в морфофункциональных чертах в соответствии с воздействием окружающей среды.
По сходству морфофункциональных характеристик могут быть выделены регионы-аналоги. Например, в адаптивном комплексе арктических аборигенов проявляются черты приспособления к гипоксии и низким температурам, так же как в высокогорье, хотя природа гипоксии в этих регионах разная. Может быть, и в таких географических зонах, как пустыня и континентальная часть Сибири, есть какие-то общие факторы, вызывающие одинаковую реакцию у их коренных обитателей (я имею в виду понижение уровня минерализации скелета в условиях континентальной Сибири и Каракумов).
Адаптивные типы, проявляясь в пределах вида Homo sapiens, не представляют собой экологически специализированных форм, они выражаются в виде тенденции к изменению физиологических и морфологических черт в направлении, наиболее благоприятном для существования в определенной среде и не препятствующем возможности существования в других экологических нишах.
Адаптивные реакции осуществляются в двух формах — общей и специфической. Общей особенностью адаптивных типов можно, по-видимому, считать повышение сопротивляемости организма неблагоприятным воздействиям среды. Это выражается в увеличении иммунных фракций сыворотки крови и, может быть, в увеличении костно-мускульной массы тела. Специфические же реакции очень разнообразны. В одном случае это повышение теплопродукции, в другом — относительной поверхности испарения, в третьем — и того и другого признака и т.п.
Именно эти специфические реакции и дают право называть выделенные морфофункциональные комплексы арктическим, высокогорным, континентальным, тропическим и т.п. Адаптивный тип — это лишь одна из форм приспособительной реакции у человека — биологическая. Как можно представить себе природу адаптивного типа? Он не равноценен расе, поскольку проявляется независимо от этноса и расы в сходных условиях существования человеческих популяций. Адаптивный тип — это норма реакции, конвергентно возникающая в сходных условиях обитания в популяциях, которые могут быть генетически не связаны между собой, в то время как расовая общность непременно предполагает общее происхождение всех входящих в нее групп, живущих на определенной территории.
Независимость адаптивного типа от расовой и этнической принадлежности неоднократно демонстрировалась в предшествующем изложении. Сходный комплекс приспособительных черт в тропической зоне находим у европеоидного населения Индии, австралийцев и аборигенов Африки; на Крайнем Севере саамы, ненцы, чукчи и эскимосы также обнаруживают единое направление приспособительных реакций; то же самое относится и к коренным жителям пустынь, различным в этническом и расовом отношениях" (с. 236239).
"Легко понять, что адаптивные типы не совпадают с расовыми, которые могут быть приспособительными, а могут и не быть. Они иногда перекрещиваются с ними, а чаще всего охватывают несколько локальных расовых комплексов. Поэтому адаптивные типы имеют, как правило, очень большие ареалы, вытянутые в широтном направлении из-за приспособления к однородным условиям географической среды. Эти ареалы приурочены, следовательно, к определенным природным зонам. Но есть и азональные адаптивные типы, например, адаптивный комплекс приспособлений к высокогорью. В отличие от других адаптивных типов он не имеет сплошного ареала и приурочен к районам горных поднятий.
Автору гипотезы адаптивных типов принадлежит и первичная их классификация, а также попытка их морфологической и физиологической характеристик. Наиболее нейтральный из адаптивных типов — тип умеренного пояса. Его ареал охватывает благоприятную для жизни среду. Она полноценна в геохимическом отношении, и тем самым создается благоприятная возможность для развития продуктивного хозяйства, а значит, и для обеспечения разнообразным питанием.
У адаптивного типа умеренного пояса более или менее гармонично развиты морфологические и физиологические адаптивные признаки, соответствующие среднемировым. У адаптивного типа арктического пояса лучше развиты скелет и мускулатура, выше уровень жироотложения у женщин. Высокое содержание белка в сыворотке крови обеспечивает жизнестойкость организма, повышение активности обменных процессов. Для адаптивного типа континентальной зоны характерны до какой-то степени противоположные свойства: тенденция к жироотложению и понижение уровня активности обменных процессов. Нельзя не отметить, что эти физиологические особенности находят какую-то грубую аналогию в физиологии млекопитающих, населяющих пустынные районы с нарушенным водоснабжением. У тех также наблюдается сезонное усиление упитанности, причем у некоторых видов в благоприятных условиях и при обильном кормлении образуются локальные очаги жироотложения. Наконец, адаптивный тип высокогорья образовался при адаптации к гипоксии — недостатку кислорода на больших высотах. Поэтому они устойчивы к гипоксии, выражающейся в минимальном жироотложении, сравнительно низком уровне газообмена и содержания гемоглобина.
21. Соотношение биологического и социального в становлении личности человека
Биосоциальная природа человека
Человек - биосоциальное существо, генетически связанное с другими формами жизни, но отделившееся от них благодаря способности производить орудия труда; обладающее членораздельной речью и сознанием, творческой активностью и нравственным самосознанием; высшая ступень развития живых организмов на Земле.
Биосоциальная природа человека находит отражение в том, что его жизнь определяется единой системой условий, в которую входят как биологические, так и социальные элементы.
Биологические элементы
1. Наследственность
2. Изменчивость
3. Естественный отбор
Социальные элементы
1. Труд.
2. Речь
3. Организация совместной жизни
Понятие антропогенеза.В широком смысле "антропология" - наука о человеке (от греч. "антропос" - человек). Но так как человека изучает множество наук, как естественных, так и гуманитарных, то за антропологией в узком смысле осталась проблема происхождения человека и определения специфики его строения и эволюции.Бурное развитие антропология получила во второй половине XIX века после создания теории эволюции Дарвина. Однако разработав теорию естественного происхождения человека, он не включил в нее влияние социального фактора на его развитие. Кроме того, в теории Дарвина отсутствует качественное отличие ума человека от животного. Это во многом объясняется тем, что он не затрагивает роли труда в процессе антропогенеза.Внимание на это было обращено в трудовой теории антропогенеза, защитником которой был, в частности, Ф. Энгельс. Представители данной теории считали, что труд не отменяет действие биологических законов, но преобразовывает характер действия естественного отбора. Труд в процессе становления развивает в человеке способность преобразовывать природу по своим меркам, а также способствует его собственному формированию.Кроме того, орудия труда являются и основным способом передачи социального опыта, т.е. лежат в основе новой, социальной формы наследования, которую Дарвин не рассматривает.Следовательно, становление человека и общества - процесс взаимосвязанный, процесс антропосоциогенеза, и главная роль в нем принадлежит трудовой деятельности.Необходимо отметить, что антропогенез не следует представлять в виде линейного процесса.Основная проблема в восстановлении эволюции человека состоит в том, что у нас нет близких родственников среди живущих ныне предков. Наши ближайшие, хотя и не очень близкие, в настоящее время живущие родственники - шимпанзе и горилла - были связаны с нами общим предком не менее 7 млн лет назад.Новый аспект критики дарвиновской концепции возник на волне первых успехов генетики, зародившейся на рубеже XX в. Появилась мутационная теория эволюции нидерландского ученого Хуго де Фриза, согласно которой новые виды возникают скачкообразно, в результате крупных единичных мутаций в генном наследственном аппарате (геноме). И это явление никак не связано с естественным отбором, о котором писал Дарвин.Критика дарвинизма с различных точек зрения была широко распространена в биологии до конца 20-х гг., когда произошел синтез классического дарвинизма с новейшими достижениями генетики, который получил название синтетической теории эволюции. Большую роль при этом сыграла популяционная генетика, основанная отечественными биологами С.С. Четвериковым и Н.В. Тимофеевым-Ресовским. Она изучает элементарные эволюционные процессы не в индивидуальном организме, а в популяциях животных и растений.Существуют и другие теории, однако на наш взгляд, указанные подходы в общем виде согласуются с теорией самоорганизации систем.В ее основе лежит принцип самоорганизации как движущей силы развития любых открытых неравновесных систем, т.е. систем, обменивающихся со средой веществом, энергией и информацией, которые переходят от одного качественного состояния к другому в результате скачкообразного процесса. Причем состояние системы после скачка, согласно данной теории, носит случайный характер.К таким системам относятся все биологические системы, включая человека. Разработка теории самоорганизации началась совсем недавно и связана прежде всего с таким направлением в науке, как синергетика.Эволюция человека.РАМАПИТЕК. Самый древний из известных науке предок человека. Его останки обнаружены в Северо-Западной Индии и Восточной Африке. Рамапитек (назван по имени индийского бога Рамы) отмечает очень важную точку в эволюции родословной приматов, когда разделяются пути высших приматов и той ветви, которая ведет к человеку. Рамапитек жил 16-12 млн лет назад.АВСТРАЛОПИТЕК. В двух областях Восточной Африки обнаружены останки рода Australopithecus (австралопитек). Следы этих ближайших к человеку высших приматов обнаружены в молодых отложениях (2 млн лет) у озера Рудольф (Кения), откуда известны и самые древние представители австралопитеков возрастом 5,5 млн лет. Австралопитеки были "почти людьми" не только по внешнему виду. Они могли производить орудия труда, хотя и самые примитивные. Австралопитек жил 6-2 млн лет назад.ХАБИЛИС (Homo habilis - человек умелый). Хабилис - древнейший предок человека, он древнее и примитивнее питекантропа. К этому виду относят находки из Коби-Фор (Кения) возрастом 2,6 млн лет и из долины реки Омо (Эфиопия) возрастом 2 и 4 млн лет. Больше всего останков хабилиса (презинджантропы, зинджантропы) обнаружено в ущелье Олдовай (Танзания) в отложениях возраста 1,75-1,85 млн лет. Хабилис имел рост 122-140 см и объем мозга 656 см3
(больше, чем у австралопитека, но меньше по сравнению с питекантропом). Его анатомическое строение очень примитивно, создаваемые орудия весьма элементарны, а объем мозга ниже критической границы (750 см3
).АРХАНТРОП. На смену хабилисам пришли архантропы, представленные яванским питекантропом, синантропом, гейдельбергским человеком и рядом других форм, обнаруженных в Европе, Азии и Африке. Между этими древними существами имеются морфологические или хронологические переходы. Вот почему они были объединены известным антропологом Ле Гро Кларком (1964) под одним наименованием - Homo erectus (человек прямоходящий). Открытие яванского питекантропа - "недостающего" звена в родословной человека - было истинным триумфом науки. В 1891 г. антрополог Э. Дюбуа открыл на острове Ява части скелета человекоподобного существа (питекантропа). Первые яванские питекантропы возрастом 1,5-1,9 млн лет имели объем мозга 800 см3
. Большинство яванских питекантропов имеют возраст 0,5-1 млн лет. Средний объем их мозга составлял 860 см3. Более поздние питекантропы - синантропы (обнаружены в 50 км от Пекина) жили 200-500 тыс. лет назад и имели объем мозга от 915 до 1225 см3
. Средний рост мужских особей достигал 162-163 см, а женских - 152 см.НЕАНДЕРТАЛЕЦ. Занимает промежуточное положение между древнейшими людьми (архантропами типа Homo erectus) и первыми представителями Homo sapiens - кроманьонцами. Его название происходит от пещеры Неандерталь, расположенной BOXIC Дюссельдорфа (ФРГ). Неандерталец при росте 155-165 см имел объем мозга около 1300- 1600 см3
. Неандертальцы создали мустьерскую культуру каменного века (150-40 тыс. лет назад).КРОМАНЬОНЕЦ - представитель Homo sapiens (человек разумный). Впервые останки неоантропа были обнаружены в 1868 г. в пещере Кроманьон (Франция), откуда идет название первых представителей сапиенсов2. Рост кроманьонца 180-187 см, объем мозга до 1600 см3
. Преимуществом кроманьонца (неоантропа) перед неандертальцем была трудовая деятельность.Биологическое и социальное в историческом развитии человекаВ истории науки в вопросе соотношения биологических и социальных факторов в индивидуальном развитии человека, или в его онтогенезе, встречаются самые различные точки зрения. Так, немецкий биолог Э. Геккель, много сделавший для утверждения учения Дарвина, полагал, что развитие человека и общества определяется главным образом биологическими факторами, а двигателем общественного развития и эволюции человека являются борьба за существование и естественный отбор. Поэтому возникновение социал-дарвинизма, стоящего как раз на подобной точке зрения, часто связывают именно с именем Геккеля.В современной литературе существует два различных подхода к решению проблемы о роли социальных и биологических факторов в индивидуальном развитии человека. Одни авторы утверждают, что оно целиком обусловлено генами, абсолютизируя, таким образом, биологический фактор. Это направление называется панбиологизм. Вторая точка зрения состоит в том, что все люди рождаются с одинаковыми генетическими задатками, а главную роль в развитии их способностей играют воспитание и образование. Данная концепция получила название пансоциологизм.Каждый человек является носителем специфического, индивидуального набора генов, вследствие чего он, как уже говорилось, генетически уникален. Свойства человека, как и других живых существ, во многом детерминированы генотипом, а их передача от поколения к поколению происходит на основе законов наследственности. Индивид наследует от родителей такие свойства, как телосложение, рост, массу, особенности скелета, цвет кожи, глаз и волос, химическую активность клеток. Многие также говорят о наследовании способности к вычислению в уме, склонности к тем или иным наукам и т.д.На сегодняшний день господствующей точкой зрения можно считать ту, которая утверждает, что наследуются не сами способности, как таковые, а лишь их задатки, в большей или меньшей степени проявляющиеся в условиях среды.Генетическим материалом у человека, как и у других млекопитающих, является ДНК, которая находится в хромосомах. Хромосомы каждой клетки человека несут в себе несколько миллионов генов. Но генетические возможности, задатки реализуются только в том случае, если ребенок с раннего детства находится в общении с людьми, в соответствующей социальной среде.Генетический потенциал человека ограничен во времени, причем достаточно жестко. Если пропустить срок ранней социализации, он угаснет, не успев реализоваться. Ярким примером этого могут служить многочисленные случаи, когда младенцы силой обстоятельств попадали в джунгли и проводили среди зверей несколько лет. После возвращения их в человеческое сообщество они не могли уже в полной мере наверстать упущенное, овладеть речью, приобрести достаточно сложные навыки человеческой деятельности, у них плохо развивались психические функции человека. Это свидетельствует о том, что характерные черты человеческого поведения и деятельности приобретаются только через социальное наследование, через передачу социальной программы в процессе воспитания и обучения.Можно сказать, что вся цепочка предшественников современного человека и эволюция современного человека, с точки зрения сегодняшнего естествознания, может характеризоваться как эволюционный синтез биологического и социального.Эволюционная цепочка человека сочеталась с соответствующей социальной организацией, в основе которой лежала совместная трудовая деятельность (превобытно-общинный, общинный строй, рабовладельческий, феодальный, капиталистический, социалистический,....).Со способностью к труду соотносятся еще два отличительных признака человека: прямохождение, которое освободило его руки, и, как следствие развитие руки особенно большого пальца на ней. Наконец, еще два характерных признака человека, повлиявших на развитие культуры, - использование огня и захоронение трупов.Главные отличия человека от животных: понятийное мышление, речь, труд стали теми путями, по которым шло обособление человека от природы.22. Что изучает антропология?
Родиной европейской науки является древняя Греция. Возраст науки насчитывает 2000 лет и 12 тысяч научных дисциплин.
Антропология (от греч. Antropos (антрос) — человек, и logos (логос) — разум, знание) — наука о происхождении и эволюции человека. Термин введен Аристотелем.
1. Физическая антропология изучает процессы и этапы становления человека как вида, а также природу внутривидовых вариаций, их анатомические и физиологические характеристики и другие существенные биологические факты.
2. Культурная антропология занимается изучением поведения человека и результатов его деятельности:
-
Этнолингвистика. Ключом к культуре является ее понимание как системы символов.
- Доисторическая археология. Изучает антропогенез (происхождение homo sapiens) и последующее развитие человека, объясняет историческое разнообразие в образе жизни народов.
-
Культурная антропология. Третий обычно выделяемый подраздел культурной антропологии имеет одноименное с ней название. Но в разных странах (например, в Великобритании) эквивалент культурной антропологии известен там как социальная антропология (семейная, политическая, социальная организация человеческого бытия).
3. Социа́льная антрополо́гия — межпредметная научная дисциплина, изучающая современное общество с позиции антропологов.
Из вышеприведенного видно, насколько разнообразны интересы, собранные вместе под общим наименованием.
Основной задачей антропологии является изучение процесса перехода от биологических закономерностей, которым подчинялось существование первобытного предка человека, к закономерностям социальным.
Предмет включает в себя всезнание от анатомии до истории. Особенно длительная связь у антропологии отмечается с психологией и медициной.
Сам предмет имеет 2-а специфических метода: антропометрия и антропоскопия. Также широко используются смежные науки: физика, физиология, биология, генетика, медицина и ряд других наук.
Антропология состоит из 2-х разделов:
1 раздел: палеоантропология – антропология ископаемых форм;
2 раздел: антропология живых современных форм.
Первый раздел палеоантропология состоит их 4-х частей:
1) происхождение приматов;
2) происхождение человека - антропогенез;
3) происхождение человеческих рас – расогенез;
4) наука о болезнях ископаемого человека – палеопатология.
Второй раздел антропология живых современных форм состоит из 7-ми частей:
1) соматология (от греч. soma, родительный падеж somatos — тело и логия);
2) физическая антропология;
3) анатомическая антропология;
4) этническая антропология;
5) антропогенетика;
6) дерматоглифика (от греч. Derma (дерма), род. падеж dermatos кожа и glypho (глифика) вырезаю, гравирую, узор);
7) прикладная антропология.
23. Из каких частей состоит антропология живых современных форм?
Антропология живых современных форм состоит из 7-ми частей:
1) соматология (от греч. soma, родительный падеж somatos — тело и логия);
2) физическая антропология;
3) анатомическая антропология;
4) этническая антропология;
5) антропогенетика;
6) дерматоглифика (от греч. Derma (дерма), род. падеж dermatos кожа и glypho (глифика) вырезаю, гравирую, узор);
7) прикладная антропология.
24. Где впервые появился человек?
Самый древний из рода Homo — Homo habilis, или человек умелый, первые представители которого появились на Земле около 2 млн лет назад. До этого времени, вероятно, существовали только австралопитеки. Около 2,5 млн лет назад произошло расщепление в эволюции гоминид, в результате которого обособились массивные австралопитеки (тупиковая ветвь эволюции) и род Homo. Кроме находок из Олдувайского ущелья, к виду Homo habilis относят и так называемого рудольфского человека, Homo rudolfensis, череп которого был найден в Кении в 1972 году в районе оз. Рудольф (ныне оз. Туркана), а также находки из Эфиопии и Южной Африки. Древность этих видов от 2,4 до 1,9 млн лет. Предполагается, что эти первые люди были творцами орудий древнейшей на Земле олдувайской (галечной) культуры. Существуют находки, которые не нашли таксономического определения и частью исследователей относятся к Homo habilis, а другою частью — к группам архантропов (древнейших людей), пришедших на смену Homo habilis примерно 1,6—1,5 млн лет назад.
В группу архантропов входят два основных вида. Это вид азиатских древнейших людей, человек прямоходящий (Homo erectus), и его африканский вариант, человек работающий (Homo ergaster).
На протяжении многих миллионов лет предки человека населяли ограниченную область на востоке Африки. Здесь, на озере Виктория, 18 млн. лет назад жил проконсул – наш общий с человекообразными обезьянами предок; здесь более 4 млн. лет назад возник афарский австралопитек – наш прямоходящий предок. Здесь провел всю свою историю первый представитель рода Человек – умелый человек, появившийся более 2 млн. лет назад, и здесь 1,6 млн. лет назад возник прямостоящий человек. Где начал свой путь наш вид – разумный человек – не известно, но где-то поблизости.
Место формирования сапиенса
"Процессы сапиентации, как об этом свидетельствуют палеоантропологические данные, происходили в различных районах Старого Света, хотя и с разной скоростью. Здесь могли играть роль различные обстоятельства, особенности окружающей среды, специфика общественной структуры популяции и т.д. Таким образом, обе гипотезы — множественности центров сапиентации (полицентризм) или ограниченности ее одной достаточно обширной территорией (широкий моноцентризм) — имеют точки соприкосновения. Можно предположить, что, так сказать, "опережающими темпами" она происходила в Восточной Африке Юго-Восточной Европе и на Ближнем Востоке.
Сейчас много споров вызывает проблема приоритета Африки или Европы. Судя по некоторым данным, на древнейшей прародине человечества сапиенс мог появиться на несколько десятков тысяч лет раньше, чем на других территориях. Однако, если и принять эту гипотезу, она вовсе не исключает возможности того: что сапиентация происходила и в других районах Старого Света. Большая часть сторонников полицентризма признает умеренное число центров сапиентации: от двух ("дицентризм") до четырех-пяти.
Высказывалось, правда, и такое соображение: очагов возникновения сапиенса могло быть столько же, сколько было мест формирования культуры верхнего палеолита. Ведь мустьерская культура в основном связана с неандертальцем, а культура верхнего палеолита — с сапиенсом. <...>
Однако жесткой связи между каменным инвентарем и физическим типом не было, не было, видимо и непреодолимой границы между неандертальцем и сапиенсом в культурном отношении. <…>
В пользу полицентризма говорит и почти одновременное появление сапиенса на рубеже верхнего палеолита — примерно 40—35 тыс. лет назад, притом в таких удаленных друг от друга, а иногда даже окраинных районах, как Индонезия (Ниа на Калимантане), Западная Европа (Кроманьон, Ханёферзанд) или Южная Африка (Флорисбад). Один из самых существенных вопросов касается древности формирования рас современного человечества — негроидной, европеоидной и монголоидной <...>
Сами же большие расы современного человечества образовались только в постпалеолитическое время. То есть сапиенс древнее рас, процессы сапиентации и расогенеза совпадали лишь частично, и последние происходили уже на достаточно смешанной основе.
Заключительный этап гоминизации — процесс сапиентации — занял в основном последние 100 тыс. лет. На этом отрезке антропогенеза произошли существенные сдвиги в морфологической организации, познавательных способностях, снизилась скорость процессов старения, увеличилась продолжительность жизни (с. 99—101).
Распространение современного человека
В Африке были найдены более древние ископаемые остатки, чем на каком-либо из других континентов. Так, в Южной Эфиопии нашли Омо-I – неполный череп со многими современными признаками, возраст которого, вероятно, более 60 тысяч лет. В устье южноафриканской реки Клазис обнаружены "современные" остатки, возраст которых 100 тысяч лет, а в пещере Бордер найдена "современная" нижняя челюсть возрастом 90 тысяч лет.
Ископаемые черепа 40-тысячелетней давности, которые относятся к вполне современному типу, встречены в самых различных районах Азии – от Израиля до Явы. У всех них есть подбородочный выступ или другие явно "современные" признаки.
Впервые люди появились в Северной Америке, вероятно, между 70 и 12 тысячами лет назад. В периоды наибольших похолодании в это время море отступало и образовывалась широкая сухопутная перемычка Берингия, которая сейчас затоплена Беринговым проливом.
Следы очагов и ископаемые остатки, возраст которых установлен, свидетельствуют, что современный человек жил в Австралии по крайней мере уже 40 тысяч лет назад.
Вероятнее всего, люди впервые появились здесь в период от 55 до 45 тысяч лет назад, когда уровень океана был на 160 футов (50 м) ниже, чем сейчас, и многие острова составляли единое целое.
На этой шкале предоставлено развитие всех первобытных людей мира , начиная с того периода, когда возникли первые человекоподобные гоминиды (5 млн.лет назад) и до 700 г.до.н.э.
5 000 000- 2 000 000 до. н.э.
В Африке появляются первые гоминиды, обитавшие на деревьях.
2 000 000- 250 000 до. н.э.
Появляется человек прямоходящий – Homo erectus – и расселяются по Азии и Европе.
250 000 – 120 000 до. н.э.
В Африке – Нomo sapiens – которые постепенно переселяются на север.
80 000 – 30 000 до. н.э.
В Европе живут неандертальцы (обитатели пещер).
50 000 – 25 000 до. н.э.
Современные люди расселяются по территории Европы, Азии, Австралии, Америки. Изготавливают орудия труда из кости и камня.
25 000 – 10 000 до. н.э.
Первые круглые дома, рисунки и резьба.
10 000 – 9 000 до. н.э.
Изменение климата , конец ледниковой эпохи.
9 000 – 7 000 до. н.э.
Возникновение земледелия в отдельных районах мира. Поселения в Сирии, Палестине, на Кипре. Одомашнена собака.
7 000 – 6 000 до. н.э.
В восточном Средиземноморье одомашнены козы, овцы, свиньи. Изготавливают ткани льняные, керамики, применяется медь. Строятся первые города.
5 000 – 4 000 до. н.э.
В Анатолии обрабатывают медь и свинец. Одомашнены лошадь, осел. В районе Мексиканского залива выращивают маис, в Перу – хлопок, в Китае и Индии – рис.
4 000 – 3 000 до. н.э.
Шумерская цивилизация. Первая письменность. Изделия из золота, серебра, свинца. Орошение. Парусные суда на Ниле и Евфрате. Каменные храмы и гробницы на Мальте и в Европе.
3 000 – 2 000 до. н.э.
Первые фараоны Египта, иероглифическая письменность. В Месопотамии изобретена колесница. Возникновение цивилизации долины Индии. Ткани из хлопка . Методы обработки меди и изготовления тканей распространяются по всей Западной Европе.
2 000 – 1000 до. н.э.
Распространение методов обработки бронзы по Европе. Завершено строительство Стоунхенджа.
1 000 – 700 до.н.э.
Культкура ольмеков в Мексике . Кельты расселяются по Центральной Европе, Британским островам. К 700 г.до н.э. в Европе открыт секрет обработкаи железа. В Америке и Африке развиваются доисторические культуры.
25. Сколько лет тому назад появился человек на Земле?
Самые древние, достоверные представители рода человеческого известны примерно 2 -2,5 мл. лет тому назад. А появился человек современного вида 40-50 тысяч лет назад.
На протяжении многих миллионов лет предки человека населяли ограниченную область на востоке Африки. Здесь, на озере Виктория, 18 млн. лет назад жил проконсул – наш общий с человекообразными обезьянами предок; здесь более 4 млн. лет назад возник афарский австралопитек – наш прямоходящий предок. Здесь провел всю свою историю первый представитель рода Человек – умелый человек, появившийся более 2 млн. лет назад, и здесь 1,6 млн. лет назад возник прямостоящий человек. Где начал свой путь наш вид – разумный человек – не известно, но где-то поблизости.
Самый древний из рода Homo — Homo habilis, или человек умелый, первые представители которого появились на Земле около 2 млн лет назад. До этого времени, вероятно, существовали только австралопитеки. Около 2,5 млн лет назад произошло расщепление в эволюции гоминид, в результате которого обособились массивные австралопитеки (тупиковая ветвь эволюции) и род Homo. Кроме находок из Олдувайского ущелья, к виду Homo habilis относят и так называемого рудольфского человека, Homo rudolfensis, череп которого был найден в Кении в 1972 году в районе оз. Рудольф (ныне оз. Туркана), а также находки из Эфиопии и Южной Африки. Древность этих видов от 2,4 до 1,9 млн лет. Предполагается, что эти первые люди были творцами орудий древнейшей на Земле олдувайской (галечной) культуры. Существуют находки, которые не нашли таксономического определения и частью исследователей относятся к Homo habilis, а другою частью — к группам архантропов (древнейших людей), пришедших на смену Homo habilis примерно 1,6—1,5 млн лет назад.
В группу архантропов входят два основных вида. Это вид азиатских древнейших людей, человек прямоходящий (Homo erectus), и его африканский вариант, человек работающий (Homo ergaster).
Время появления сапиенса
"Существуют суждения о трех возможных уровнях выделения линии сапиенса: раннем, среднем и позднем (с. 97).
Более распространено мнение о средне или ранневерхнеплейстоценовом времени появления линии сапиенса. В качестве вероятного предка в этом случае у разных авторов фигурируют различные формы: или один из поздних прогрессивных эректусов (Вертешселлеш), или ранний архаический сапиенс (Сванскомб), или ранний прогрессивный неандерталец (Эрингсдорф).
Наконец существует мнение о позднем происхождении сапиенса. В этом случае предком обычно считаются прогрессивные палестинские палеоантропы или даже "классические" неандертальцы вюрма. В пользу последней точки зрения приводят как археологические аргументы (преемственность позднемустьерской индустрии "классических" неандертальцев и верхнепалеолитической культуры сапиенса), так и морфологические (например, тип людей Схул, у которых сочетаются "кроманьонские" и "неандертальские" черты).
Однако известно, что "классические" неандертальцы были поздними формами гоминид, сосуществовавшими с первыми неоантропами-сапиенсами. Чтобы разрешить это противоречие, предполагают, что темпы сапиентации на этом позднем коротком отрезке антропогенеза значительно ускорились вследствие гетерозиса при смещениях.
Процессы сапиентации протекали в разных популяциях палеоантропов, но разными темпами, и в силу ряда причин (например, условий обитания) не всегда достигали завершающего этапа.
Конечно, отдельные черты "сапиентного" комплекса могли появиться рано, даже в эоплейстоцене. Морфологическая изменчивость гоминид вообще очень широка, и, вероятно, немаловажную роль, как и у других приматов, играли в нем повторные мутации.
Видимо, некоторые признаки "сапиентного" комплекса могли возникать в эволюции отдельных групп гоминид давно и неоднократно. И в этом смысле мы вправе сказать, чтосапиентация имеет глубокие корни, хотя древнейшие настоящие" сапиенсы все же пока не известны ранее 0,1 —0,07 млн. лет назад" (с. 97—99).
Распространение современного человека
В Африке были найдены более древние ископаемые остатки, чем на каком-либо из других континентов. Так, в Южной Эфиопии нашли Омо-I – неполный череп со многими современными признаками, возраст которого, вероятно, более 60 тысяч лет. В устье южноафриканской реки Клазис обнаружены "современные" остатки, возраст которых 100 тысяч лет, а в пещере Бордер найдена "современная" нижняя челюсть возрастом 90 тысяч лет.
Ископаемые черепа 40-тысячелетней давности, которые относятся к вполне современному типу, встречены в самых различных районах Азии – от Израиля до Явы. У всех них есть подбородочный выступ или другие явно "современные" признаки.
Впервые люди появились в Северной Америке, вероятно, между 70 и 12 тысячами лет назад. В периоды наибольших похолодании в это время море отступало и образовывалась широкая сухопутная перемычка Берингия, которая сейчас затоплена Беринговым проливом.
Следы очагов и ископаемые остатки, возраст которых установлен, свидетельствуют, что современный человек жил в Австралии по крайней мере уже 40 тысяч лет назад.
Вероятнее всего, люди впервые появились здесь в период от 55 до 45 тысяч лет назад, когда уровень океана был на 160 футов (50 м) ниже, чем сейчас, и многие острова составляли единое целое.
На этой шкале предоставлено развитие всех первобытных людей мира , начиная с того периода, когда возникли первые человекоподобные гоминиды (5 млн.лет назад) и до 700 г.до.н.э.
5 000 000- 2 000 000 до. н.э.
В Африке появляются первые гоминиды, обитавшие на деревьях.
2 000 000- 250 000 до. н.э.
Появляется человек прямоходящий – Homo erectus – и расселяются по Азии и Европе.
250 000 – 120 000 до. н.э.
В Африке – Нomo sapiens – которые постепенно переселяются на север.
80 000 – 30 000 до. н.э.
В Европе живут неандертальцы (обитатели пещер).
50 000 – 25 000 до. н.э.
Современные люди расселяются по территории Европы, Азии, Австралии, Америки. Изготавливают орудия труда из кости и камня.
25 000 – 10 000 до. н.э.
Первые круглые дома, рисунки и резьба.
10 000 – 9 000 до. н.э.
Изменение климата , конец ледниковой эпохи.
9 000 – 7 000 до. н.э.
Возникновение земледелия в отдельных районах мира. Поселения в Сирии, Палестине, на Кипре. Одомашнена собака.
7 000 – 6 000 до. н.э.
В восточном Средиземноморье одомашнены козы, овцы, свиньи. Изготавливают ткани льняные, керамики, применяется медь. Строятся первые города.
5 000 – 4 000 до. н.э.
В Анатолии обрабатывают медь и свинец. Одомашнены лошадь, осел. В районе Мексиканского залива выращивают маис, в Перу – хлопок, в Китае и Индии – рис.
4 000 – 3 000 до. н.э.
Шумерская цивилизация. Первая письменность. Изделия из золота, серебра, свинца. Орошение. Парусные суда на Ниле и Евфрате. Каменные храмы и гробницы на Мальте и в Европе.
3 000 – 2 000 до. н.э.
Первые фараоны Египта, иероглифическая письменность. В Месопотамии изобретена колесница. Возникновение цивилизации долины Индии. Ткани из хлопка . Методы обработки меди и изготовления тканей распространяются по всей Западной Европе.
2 000 – 1000 до. н.э.
Распространение методов обработки бронзы по Европе. Завершено строительство Стоунхенджа.
1 000 – 700 до.н.э.
Культкура ольмеков в Мексике . Кельты расселяются по Центральной Европе, Британским островам. К 700 г.до н.э. в Европе открыт секрет обработкаи железа. В Америке и Африке развиваются доисторические культуры.
26. С появлением, какого доисторического человека стала развиваться культура?
Homohabilis(человек умелый), по общему мнению, представлял собой первый известный вид нашего рода Homo. Homohabilis представлял собой двуногое существо ростом 120—140см. У этого вида сравнительно более крупный мозг, чем у австралопитека; таким образом, мозговая коробка была больше, а лицо меньше и менее выдвинуто вперед. Верхняя и нижняя челюсти были меньше, чем у австралопитека бойсеи (зинджа), но почти не отличались от челюстей питекантропа и современного человека, зубной ряд имел открытую форму, похожую на латинскую букву U. Руки были короче по сравнению с ногами. Кисть руки человека "умелого" была способна к силовому захвату большой мощности; об этом свидетельствовали широкие ногтевые фаланги и массивные трубчатые кости кисти. Форма тазовых костей позволяла как ходить на двух ногах, так и рожать детей с крупными головами. Морфологически Homohabilis тесно примыкает к австропитековым.
Первоначально Homohabilis назвали "презинджантроп" (то есть предшественник зинджантропа), поскольку останки его обнаружили в немного более глубоком горизонте. Однако в дальнейшем выяснилось, что презинджантроп был современником зинджантропа.
На месте нынешнего Олдувайского ущелья, у озера (или водного потока) существовавших около 2 млн лет назад, в самом низу — слой (I) толщиной до 40 м, были обнаружены останки австралопитековых — зинджантропа и человека "умелого" и вместе с ним самые древние орудия человека, названные олдувайскими. Возраст этих находок по калий-аргону — 1,75-1,85 млн лет. Примитивное "праорудие" получило название "чоппер", то есть "ударник", а сама эта древнейшая каменная культура стала именоваться галечной, или олдувайской, — по имени ущелья.
Заслужить высокое звание Homo habilis (человек умелый) - значило овладеть рядом совершенно необходимых умений, приобрести сумму знаний. Во мраке пещер одним из первых понадобилось умение сохранять и добывать огонь. Постоянные очаги палеолитических убежищ использовались для обработки продуктов питания и создания комфортной среды обитания. Но одновременно они были своеобразным социокультурным центром, символом бытового, производственного и идеологического единства древних микроколлективов. Вероятно, отсюда берут начало многие культы и обряды, в том числе и неугасимый огонь на захоронениях павших в сражениях XX века...
Культура
Строение голосового аппарата и мозга неандертальцев позволяют сделать вывод о том, что они могли говорить. Самый ранний известный музыкальный инструмент — костяная флейта с 4 отверстиями (англ.) — принадлежит неандертальцам. Неандертальцы умели использовать самодельные инструменты и оружие, но, по-видимому, у них не было никакого метательного оружия.
Неандертальцы занимались собирательством и охотой. Хотя существуют версии, что они не столько охотились, сколько подбирали падаль, убитую другими хищниками. Неандертальцы хоронили своих умерших. В гроте Ля-Шапель-о-Сен во Франции было обнаружено неглубокое захоронение со скелетом в позе эмбриона, покрытым красной накидкой. Рядом с телом были оставлены инструменты, цветы, яйца и мясо, что свидетельствует о вере в загробную жизнь и наличии религиозно-магической практики.
Европейские неандертальцы практиковали каннибализм, однако также они употребляли в пищу дельфинов, тюленей и морских моллюсков.
Культура неандертальцев (т. н. "мустьерская", или, что то же самое, среднепалеолитическая культура) — это, прежде всего, обоюдоострые рубила, заточенные более качественно, чем аналогичные орудия H.erectus; а также разнообразные отщепы, использовавшиеся для разделки туш. У неандертальцев имелись также деревянные копья для ближнего боя с каменными наконечниками. В более позднее время, уже во время контактов с сапиенсами (см. ниже) у неандертальцев появляются зачатки искусства (ожерелье из медвежьих когтей, нечто вроде "флейт" — кости с просверлёнными дырочками, которые, впрочем, могли служить для разведения огня, а не для музыкальных упражнений.

Одна из первых попыток восстановить облик неандертальца
Неандертальцы были носителями Мустьерской культуры.
Кроманьонский человек и человек современного физического типа (неоантропы)
Под этим названием понимают как ископаемые формы человека современного физического типа, так и ныне живущих людей. В 1868 г. в пещере Кро-Маньон (Франция) был найден скелет человека, объем черепной коробки которого составлял 1800 мл. Для него было характерно отсутствие надбровных валиков (они разделились на две надбровные дуги), наличие подбородка, прямой лоб. Рост его составлял около 180 см. Этого человека назвали кроманьонцем. В дальнейшем сходные ископаемые формы были найдены также на территориях Китая, Палестины, Чехии, России и в других странах Европы, Африки и Азии. Средний возраст кроманьонцев составлял около 30 лет.
Считалось, что человек современного физического типа появился 40—50 тыс. лет назад. В соответствии с новыми данными этот срок определен в пределах 100-200 тыс. лет назад. Кроманьонцы не отличаются от людей современного физического типа. Если бы кроманьонец, одетый в современную одежду, шел по улице или находился бы в учебной аудитории, его было бы трудно выделить среди окружающих людей. В их пещерах были найдены стрелы, гарпуны, иглы, что указывает на усовершенствованные методы изготовления орудий. Они были искусными охотниками и ответственны за истребление мамонтов, саблезубых тигров, бизонов и др. Примерно около трети родов млекопитающих было уничтожено еще 50 тыс. лет назад. Предполагают, что резкое уменьшение диких животных способствовало переходу людей от охотничьего сообщества к сельскохозяйственному. На стадии существования неоантропа на развитие человека стали оказывать все большее влияние культура и социальные закономерности.
27. Кто был предком неандертальца?
Неандерта́лец, человек неандертальский (лат. Homo neanderthalensis или Homo sapiens neanderthalensis) — ископаемый вид людей (палеоантропов), обитавших 300—24 тысячи лет назад.
Древние люди (палеоантропы)
В 1856 г. в долине Неандерталь реки Дюссель в ФРГ были обнаружены части скелета (череп и другие костные фрагменты) древнего человека, названного в 1864 г. У.Кингом неандертальцем (homosapiensneanderthalensis). Останки этого существа были описаны К.Фульроттом.
Череп этих древних людей был значительно удлинен, с характерным низким сводом и выступающими надбровными валиками. Объем мозга - 1400 мл, рост - около 160 см. Им были свойственны широкие зубы, они обладали речью, но, вероятно, нечленораздельной.Считают, что было две группы древних людей - ранняя и поздняя. Ранние неандертальцы произошли от одной из популяций homoerectus и существовали в течение 300-250 тыс. лет. Они считаются предками как поздних неандертальцев, так, возможно, и современных людей. Жили поздние неандертальцы 200-135 тыс. лет назад на территории, ныне занимаемой Францией, Бельгией, Испанией, Италией. Объем головного мозга -до 1450 мл, средний рост 155-165 см. Средний возраст составлял 30 лет. Неандертальцы хоронили умерших или погибших соплеменников.40-30 тыс. лет назад неандертальцы внезапно исчезли. Поскольку у них были недоразвиты лобные доли головного мозга, которые обычно ответственны за поведение, то предполагают, что конфликты между ними вели к самоистреблению. По другой версии неандертальцы были либо поглощены, либо частично истреблены людьми современного физического типа.Новейшие данные не подтверждают предположения о том, что неандертальцы были предками современного человека. Считают, что эволюционные расхождения между неандертальцами и людьми современного физического типа является очень большим. Более того, известно мнение о том, что homosapiensneanderthalensis, homoerectus, homosapiens являются разными видами рода homo.
28. Кто составил переходную ступень от обезьяны к человеку?
Homohabilis - человек умелый
В 1960 г. в Африке Л.Лики нашел останки другого существа, более близкого к человеку, чем австралопитек, древность которого составляла 2 млн лет. Его рост был равен 120 см, объем головного мозга - около 650 мл. Первый палец его стопы не был отведен в сторону, что указывает на полное завершение формирования прямохождения. Кости кисти этого существа сочетали как обезьяньи, так и человеческие черты. Рядом с костными останками были найдены грубые галечные орудия и расколотые кости (для добывания мозга). Исходя из этого Л.Лики заключил, что существо было творцом этих орудий. Чтобы подчеркнуть способность изготавливать простейшие орудия, ученый назвал его homohabilis - человек умелый.
Предполагается, что австралопитек (africanus) эволюционировал независимо от австралопитека (robustus) и австралопитека (boisei), которые родственны между собой, и что эволюционная линия, ведущая от австралопитека к роду homo, начинается с австралопитека (africanus).
Представители этого вида предков человека пересекли границы звериной сущности. Можно сказать, что они уже не были обезьянами, но пока еще не были и людьми. Их рост составлял 120-130 см, вес - 50 кг, объем головного мозга - около 500 мл.
Древнейшие люди (китайский синантроп, яванский питекантроп), или архантропы
Архантропы занимают среднее положение между австралопитеком и современным человеком. С зоологической точки зрения они входят в один вид homoerectus (человек прямоходящий). Открытие архан-тропов произошло в 1891 г. на острове Ява врачом из Голландии Э.Дюбуа. На глубине 15 м плейстоценовых отложений он обнаружил коренной зуб, бедренную кость и часть черепа ископаемого существа, названного им pithecanthropuserectus (от греч. pithecus - "обезьяна", ап-thropos - "человек"), или яванским человеком.
Для питекантропа были характерны следующие признаки: головной мозг объемом 900 мл, вертикальное положение тела, скелет и зубы, сходные с человеческими, преобладание черепной коробки над лицевым отделом черепа. Но кости черепа были утолщены, а рост его составлял 170 см, масса - около 70 кг, возраст - около 650 тыс. лет. Позже на острове Ява были обнаружены грубые каменные орудия, что свидетельствует о способности яванского питекантропа к трудовой деятельности.
В 1927 г. в Китае, недалеко от Пекина, были найдены останки ископаемого существа, названного синантропом (китайским человеком). Объем его мозга был равен 1200 мл, рост - 150 см, возраст - 400 тыс. лет. Синантропы производили орудия труда и пользовались огнем. Предполагается, что они были каннибалами. Древнейшие люди умели изготавливать орудия из камня и гальки. У всех древнейших людей над глазницами проходил сплошной, сильно выдающийся вперед костный валик (надглазничный). Пищу, они добывали путем коллективной охоты на крупных животных, включая мамонтов. В этот период возникает разделение труда между теми, кто охотился, и теми, кто оставался в пещере для наблюдения за детьми, стариками и для выполнения других работ.
Предполагают, что древнейшие люди берут начало от одной из ветвей австралопитеков, возможно, homohabiiis, которые 2-1,5 млн лет назад начали расселяться по Африке, а затем проникли в Средиземноморье, Южную, Центральную и Юго-Восточную Азию.
Существует гипотеза, что древнейшие люди были прямыми предками ранних представителей древних людей (палеоантропов), хотя время, способ эволюции и география процесса остаются дискуссионными.
29. Какой объем головного мозга был у китайского синантропа?
В 1927 г. в Китае, недалеко от Пекина, были найдены останки ископаемого существа, названного синантропом (китайским человеком). Объем его мозга был равен 1200 мл, рост - 150 см, возраст - 400 тыс. лет. Синантропы производили орудия труда и пользовались огнем. Предполагается, что они были каннибалами. Древнейшие люди умели изготавливать орудия из камня и гальки. У всех древнейших людей над глазницами проходил сплошной, сильно выдающийся вперед костный валик (надглазничный). Пищу, они добывали путем коллективной охоты на крупных животных, включая мамонтов. В этот период возникает разделение труда между теми, кто охотился, и теми, кто оставался в пещере для наблюдения за детьми, стариками и для выполнения других работ.
30. Кто впервые выделил четыре типа темперамента?
Гален - впервые описал четыре типа темперамента.
Темперамент - характеристика индивида со стороны динамических особенностей его психической деятельности. Основные компоненты:
- общая активность индивида.
- моторика индивида (двигательные особенности);
- эмоциональность.
Темперамент проявляется уже в раннем детстве, он относительно устойчив и слабо поддается воспитанию. Основными разновидностями темперамента считаются четыре классических типа, предложенные римским врачом Галеном (ок. 130 - ок. 200 гг. н.э.). Чаще всего встречается темперамент смешанного типа.
| Тип темперамента, по Галену |
Краткая характеристика |
Свойства нервной системы, по И.П.Павлову |
Выдающиеся личности |
| флегматик |
пассивный, очень трудоспособный, медленно приспосабливающийся. Эмоции внешне проявляются слабо |
спокойный, сильный, уравновешенный, мало
подвижный
|
И.А.Крылов М.И.Кутузов И.Ньютон |
| сангвиник |
активный, энергичный, легко приспосабливающийся |
живой, сильный, уравновешенный, подвижный |
М.ЮЛермонтов Наполеон I В.А.Моцарт |
| холерик |
активный, очень энергичный, настойчивый. Эмоции не управляемые |
легко возбудимый, сильный, неуравновешенный, подвижный |
Петр1 А.С.Пушкин А.В.Суворов М.Робеспьер |
| меланхолик |
пассивный, легко утомляющийся, тяжело приспосабливающийся, очень чувствительный |
слабый, неуравновешенный, сдержанный, малоподвижный |
Н. В. Гоголь П.И.Чайковский |
31. По какой формуле исчисляется индекс Пинье?
Классификация конституции человека по М.В.Черноруцкому. Индекс Пинье
М.В.Черноруцкий выделил 3 типа конституции человека:
1) астенический;
2) нормостенический;
3) гиперстенический.
Для того чтобы определить, какой тип конституции у данного человека, нужно определить индекс Пинье по формуле L - ( Р + Т ), где L -длина тела, Р - вес тела в кг, Т - окружность грудной клетки в см.
Если индекс Пинье больше 30 - астенический тип сложения, меньше 10 - гиперстенический. От 10 до 30 - нормостенический.
Конституцию можно определить как основную биологическую характеристику целостного организма человека, проявляющуюся в особенностях его реактивности и резистентное™ (устойчивости), в основе которых лежит специфическая норма реакций индивидуального генотипа.
32. Сколько существует больших рас?
Формирование больших рас
Примерно 100 тыс. лет назад на континенте Азия с прилегающими областями Африки и Европы обозначились две основные расовые группы: юго-западная и северо-восточная. Их разделяли горные цепи Гималаев, Гиндукуша и хребтов Индокитая.
Первая ветвь дала начало европеоидной и негроидно-австроло-идной большим расам.
Вторая ветвь образовала монголоидную расу.
Понятие о расе. Большие расы
Расы - это территориальные группы людей, выделяемые на основе их генетического родства, которое проявляется в определенном физическом сходстве. Большинство отечественных антропологов различают три большие расы человечества - монголоидную ("желтую"), европеоидную ("белую") и негроидно-австролоидную ("черную").
Европеоиды светлокожи, для них характерны светло-русые или темно-русые, мягкие или средней жесткости волосы, серые, серо-зеленые или каре-зеленые широко открытые глаза, умеренно развитый подбородок, широкий таз, узкий и сильно выступающий нос, нетолстые губы, обильный волосяной покров тела и лица.Негроиды темнокожи, у них курчавые или шерстистые темные волосы, толстые губы, очень широкий и плоский нос, крупные зубы, карие или черные глаза, удлиненная форма головы, редкая растительность на лице и теле, узкий таз, большие ступни.Монголоиды обладают желтой или желто-коричневой кожей. У них прямые жесткие черные волосы, плоское крупное скуластое лицо, узкие раскосые карие глаза со складкой верхнего века (эпикантус) во внутреннем углу глаза, плоский широкий нос, редкая растительность на лице и теле.Австролоиды имеют шоколадный цвет кожи. У них темные волнистые волосы, крупная голова и массивное лицо с широким и плоским носом, выступающим подбородком, значительный рост волос на лице и теле.Большие расы делятся на более мелкие. Последние распадаются на группы антропологических типов. Между расами существуют переходные группы, поэтому современное человечество можно рассматривать как переплетение множеств антропологических типов и рас, составляющих единое биологическое целое.
33. Где началось формирование больших рас на Земле?
Формирование больших рас
Примерно 100 тыс. лет назад на континенте Азия с прилегающими областями Африки и Европы обозначились две основные расовые группы: юго-западная и северо-восточная. Их разделяли горные цепи Гималаев, Гиндукуша и хребтов Индокитая.
Первая ветвь дала начало европеоидной и негроидно-австроло-идной большим расам.
Вторая ветвь образовала монголоидную расу.
34. В какой структуре клетки содержится генетическая программа человека?
Наследственность человека
Генетическая программа homosapiens определяет его как биологический вид. Элементарной единицей наследственности является ген, характеризующийся рядом признаков. Гены располагаются в ядрах клеток. По своему уровню ген - внутриклеточная молекулярная структура. По химическому составу это нуклеиновые кислоты, в составе которых основную роль играют азот и фосфор. Гены - это "мозговой центр" клеток. Генетическая программа записана в молекулах ДНК и является достаточно консервативной.
35. Когда возник человек современного типа на Земле?
Считалось, что человек современного физического типа появился 40—50 тыс. лет назад. В соответствии с новыми данными этот срок определен в пределах 100-200 тыс. лет назад. Кроманьонцы не отличаются от людей современного физического типа. Если бы кроманьонец, одетый в современную одежду, шел по улице или находился бы в учебной аудитории, его было бы трудно выделить среди окружающих людей. В их пещерах были найдены стрелы, гарпуны, иглы, что указывает на усовершенствованные методы изготовления орудий. Они были искусными охотниками и ответственны за истребление мамонтов, саблезубых тигров, бизонов и др. Примерно около трети родов млекопитающих было уничтожено еще 50 тыс. лет назад. Предполагают, что резкое уменьшение диких животных способствовало переходу людей от охотничьего сообщества к сельскохозяйственному. На стадии существования неоантропа на развитие человека стали оказывать все большее влияние культура и социальные закономерности.
36. В какой части света был впервые обнаружен австралопитек?
Австралопитек - непосредственный предок человека
В 1924 г. Р.Дарт, профессор Иоганнесбургского университета, обнаружил в Южной Африке череп ископаемого гоминида, который по строению костей лицевого отдела черепа и зубов принадлежал существу, занимавшему среднее положение между обезьяной и человеком. Р.Дарт назвал его австралопитеком, или южной обезьяной (от лат. australis - "южный", pithecus - "обезьяна"). Возраст южноафриканского австралопитека составляет около 3,0—2,3 млн лет.
- Австралопитек (africanus), 1924 г. Южная Африка
- Австралопитек (afarensis) - "Люси", 1978 г. Восточная Африка. Возраст 3,9-3,4 млн лет
- Австралопитек (ramidis), 1994 г. Эфиопия. Возраст 4,4 млн лет
- Австралопитек (robustus), 1938 г. Южная Африка. Возраст 1,9—1,6 млн лет
- Австралопитек (boisei), 1959 г. Танзания, первое название зинджантроп
На основании изучения тазовых и бедренных костей австралопитеков считается, что они были двуногими существами, способными стоять и передвигаться подобно человеку. Жили они в пещерах, которые использовали в качестве убежищ, и изготавливали простейшие орудия. Австралопитеки были охотниками на мелких животных, но также использовали в пищу зерна, плоды диких деревьев, корни растений.
37. В какие генетические процессы вовлекается ДНК человека?
Генетические процессы
От поколения к поколению ДНК человека вовлекается в разнообразные генетические процессы:
1. Мутационный процесс, непосредственно изменяющий структуру ДНК.
2. Миграция генов - их отток или приток из других популяций.
3. Дрейф генов - случайные колебания частот генов.
4. Естественный отбор — процесс, направленно изменяющий частоту генетических признаков.
Мутационный процесс в условиях естественного радиационного фона Земли не может повлиять на жизнь популяций, но повышенный уровень радиации обладает исключительной мутационной активностью, как и химические мутагены.
Случайный дрейф и миграция генов приводит к образованию эко-типов, которые, развиваясь изолированно, могут образовать в результате эволюции новую расу или популяцию.
Естественный отбор сыграл решающую роль в эволюции всех видов, в том числе и homosapiens. Человек современного типа возник в последнюю ледниковую эпоху — примерно 40-50 тыс. лет тому назад. В этот период он занимался охотой, собирательством, позже - скотоводством, земледелием и ремеслами. За последние 2-3 века получило бурное развитие промышленное производство, повлиявшее на величину и качественный характер давления естественного отбора. Человек, являясь биосоциальным существом, не освободился от действия общебиологических закономерностей, универсальных для всего живого. По данным научной литературы, 5 % эмбрионов человека погибает на ранних стадиях онтогенеза (спонтанные аборты), 3 % - смертность до наступления репродуктивного возраста, 20 % взрослых не вступают в брак и 10 % браков бесплодны.
Таким образом, примерно 50 % первичного генофонда не воспроизводится в следующем поколении, значительная часть этих явлений генетически обусловлена.
38. Назовите природные факторы расообразования
Природные факторы расообразования
Природные факторы классифицируются на 3 группы:
1. Космические факторы воздействуют на наследственную изменчивость человека, вызывают мутацию ДНК
2. Физико-географические агенты выражают соотношение балансов тепла и влаги, индекс сухости и т.д.
3. Адаптация человека к биосфере планеты как защитная реакция против вирусных и микробных инфекций
Географическая изменчивость является основой расообразования. Согласно популяционной концепции раса есть совокупность популяций.
Согласно типологической концепции раса есть сумма индивидуумов.
Расовые вариации. При изучении антропогенеза, т.е. происхождения Homo sapiens, неизменно встает вопрос и о происхождении расовых типов: развились ли они после того, как эволюция достигла ступени H.sapiens sapiens, или на более ранних стадиях эволюции гоминид. Когда первые H.sapiens sapiens появились в Европе, Африке и Азии (конец Старого Каменного века), им уже были свойственны определенные расовые вариации. Так, кроманьонский человек в Европе четко отличался от сопоставимых ранних форм человека в Австралии, Африке и Азии. Вместе с тем нужно отметить, что в общем ни в одном регионе мира анатомические характеристики ранних ископаемых H.sapiens не были полностью идентичны современным параметрам расовых типов.
Некоторые авторы полагают, что расовые вариации – относительно недавнее адаптивное явление. Другие же – с учетом широкого распространения представителей более ранних гоминид, их относительной изоляции друг от друга и длительности того периода времени, который необходим для осуществления такой дифференциации, – относят происхождение расовых различий к более древним H.sapiens и даже к уровню, предшествующему H.sapiens. Теория о том, что расовые типы современного человека начали развиваться еще на стадии H.erectus, была убедительно разработана Ф.Вейденрейхом в результате изучения ископаемых остатков пекинского человека. В этой популяции, обитавшей в Китае, Вейденрейх идентифицировал ряд признаков, которые не только отличают пекинского человека от живших в иных регионах H.erectus, но и обнаруживаются также у монголоидной ветви современного человека.
39. В какой стране впервые был обнаружен неандерталец?
Древние люди (палеоантропы)
В 1856 г. в долине Неандерталь реки Дюссель в ФРГ были обнаружены части скелета (череп и другие костные фрагменты) древнего человека, названного в 1864 г. У.Кингом неандертальцем (homosapiensneanderthalensis). Останки этого существа были описаны К.Фульроттом.
Череп этих древних людей был значительно удлинен, с характерным низким сводом и выступающими надбровными валиками. Объем мозга - 1400 мл, рост - около 160 см. Им были свойственны широкие зубы, они обладали речью, но, вероятно, нечленораздельной.
Считают, что было две группы древних людей - ранняя и поздняя. Ранние неандертальцы произошли от одной из популяций homoerectus и существовали в течение 300-250 тыс. лет. Они считаются предками как поздних неандертальцев, так, возможно, и современных людей. Жили поздние неандертальцы 200-135 тыс. лет назад на территории, ныне занимаемой Францией, Бельгией, Испанией, Италией. Объем головного мозга -до 1450 мл, средний рост 155-165 см. Средний возраст составлял 30 лет. Неандертальцы хоронили умерших или погибших соплеменников.
40-30 тыс. лет назад неандертальцы внезапно исчезли. Поскольку у них были недоразвиты лобные доли головного мозга, которые обычно ответственны за поведение, то предполагают, что конфликты между ними вели к самоистреблению. По другой версии неандертальцы были либо поглощёны, либо частично истреблены людьми современного физического типа.
Новейшие данные не подтверждают предположения о том, что неандертальцы были предками современного человека. Считают, что эволюционные расхождения между неандертальцами и людьми современного физического типа является очень большим. Более того, известно мнение о том, что homosapiensneanderthalensis, homoerectus, homosapiens являются разными видами рода homo.
40. Назовите ответственные этапы в развитии человека
Индивидуальное развитие человека
Рост развития человеческого организма
На процессы роста и развития оказывает влияние большое число самых разнообразных эндо - и экзогенных факторов. "Рост" и "развитие" обычно употребляются как понятия тождественные, неразрывно связанные между собой. Между тем, биологическая природа этих процессов различна, отличаются их механизмы и последствия.
Рост - это количественное увеличение биомассы организма за счет геометрических размеров и массы отдельных клеток.
Развитие — это качественные преобразования в многоклеточном организме за счет дифференцированных процессов (увеличения разнообразия многоклеточных структур), которые приводят к качественным и количественным изменениям функций организма.
Процесс роста и развития характеризуется следующими закономерностями:
1. Дифференциация и интеграция частей и функций, автономиза-ция развития, возрастающая в ходе филоонтогенеза.
2. Диалектическое единство непрерывного и прерывистого; постепенность и цикличность. На кривой роста человека можно отметить три основных цикла в постнатальном развитии:
а) от рождения до 10—13 лет при постоянном снижении скорости;
б) пубертатный спурт;
в) падение скорости ниже уровня до пубертатного периода и прекращение роста.
Сочетание периодов ускоренного развития и относительной стабилизации можно выявить даже на коротких отрезках онтогенеза. Например, при долговременном наблюдении с недельными интервалами детей от рождения до 2-х лет. Нелинейность роста (мини-скачки) наблюдается примерно у 70 % детей и подростков при еженедельном обследовании в течение 6-10 месяцев.
3. Гетерохрония (разновременность) в созревании разных систем организма (или тканей) и разных признаков в пределах одной системы. Согласно концепции системогенеза П.К.Анохина определяющими темпами созревают жизненно важные функции, обеспечивающие первоочередное формирование комплексных адаптивных реакций, специфических для каждого конкретного этапа взаимоотношений организма с внешней средой.
4. Значительное индивидуальное разнообразие возрастной динамики по отдельным этапам онтогенеза, зависящее от генетической программы.
Периодизация индивидуального развития (онтогенеза)
Онтогенез - комплекс последовательных преобразований организма, начиная со стадии оплодотворения яйцеклетки и до окончания жизненного цикла.
Пифагор (VI в. до н.э.) выделял 4 периода человеческой жизни: весну (до 20 лет), лето (20-40 лет), осень (40-60 лет) и зиму (60-80 лет), которые соответствовали становлению, молодости, расцвету сил и угасанию.
Гиппократ разделил весь постнатальный онтогенез на десять семилетних циклов.
Онтогенез подразделяется на 2 периода: эмбриональный и фетальный (плодный). В течение первого периода, который продолжается 8 недель, происходит формирование органов и частей тела, свойственных, взрослому человеку. В фетальный период увеличиваются размеры и завершается органообразование. Скорость роста плода возрастает до 4-5 месяцев. После 6 месяцев скорость роста линейных размеров уменьшается.
На VII Всесоюзной конференции по возрастной морфологии, физиологии и биохимии, состоявшейся в 1965 г. в Москве была принята следующая схема возрастной периодизации онтогенеза человека. Эта схема нашла широкое применение в антропологии, педиатрии, педагогике.
Схема возрастной периодизации онтогенеза человека
| Новорожденные |
1-10 дней |
| Грудной возраст |
10 дней—1 год |
| Раннее детство |
1-3 года |
| Первое детство |
4-7 лет |
| Второе детство |
8-12 лет (мальчики) 8-11 лет (девочки) |
| Подростковый возраст |
13-16 лет (мальчики) 12-15 лет (девочки) |
| Юношеский возраст |
17-21 лет (юноши) 16—20 лет (девушки) |
| Зрелый возраст (1 период) |
22-35 лет (мужчины) 21-35 лет (женщины) |
| Зрелый возраст (2 период) |
36-60 лет (мужчины) 36-55 лет (женщины) |
| Пожилой возраст |
61-74 лет (мужчины) 56-74 лет (женщины) |
| Старческий возраст |
75-90 лет |
| Долгожители |
90 лет и выше |
С момента рождения наступает период новорожденности. В это время ребенок вскармливается молозивом в течение 8-10 дней. Следующий период - грудной - продолжается до 1 года. Начало его связано с переходом к питанию "зрелым молоком". Длина тела увеличивается от рождения до года примерно в 1,5 раза, а вес утраивается. С 6 месяцев начинают прорезываться молочные зубы, и на 2-3 году жизни заканчивается прорезывание молочных зубов. В период первого детства, начиная с 6 лет, появляются первые постоянные зубы. В период второго детства выявляются половые различия в размерах и форме тела, а также начинается усиленный рост в длину. В среднем к 12—13 годам у мальчиков и девочек заканчивается смена зубов. Начинают развиваться вторичные половые признаки. Следующий период - подростковый — называют периодом полового созревания, или пубертатным, к концу которого размеры тела составляют 90-97 % своей окончательной величины, а основные функциональные характеристики подростков приближаются к характеристикам взрослого организма. В период юношеского возраста заканчивается процесс роста и формирование организма, все основные размерные признаки достигают дефинитивной (окончательной) величины. В зрелом возрасте форма и строение тела изменяются мало. Между 30-ю и 45-50-ю годами длина тела остается постоянной, а потом начинает уменьшаться. В пожилом и старческом возрасте происходят инволютивные изменения организма.
Пубертатный скачок роста
Важным событием пубертатного периода является спурт - скачкообразное увеличение роста, наблюдающееся у мальчиков в среднем в 13-15 лет, а у девочек - в 11-13 лет. В течение этого времени абсолютная скорость роста непостоянна и постепенно снижается: у мальчиков - от 12 до 7 см / год, у девочек - от 11 до 6 см / год.
В осуществлении ростового спурта задействованы многие факторы, и в первую очередь половые гормоны и гормон роста - СТГ, оказывающий свое воздействие посредством соматомединов - пептидов, синтезирующихся в печени. Помимо этих гормонов, существенное значение в данном периоде имеет универсальный анаболический гормон инсулин и гормоны щитовидной железы - тироксин и трийодтиронин. Инсулин регулирует белковый и жировой обмен, а тироксин и трийодтиронин влияют на рост и дифференцировку тканей, повышение интенсивности основного обмена и теплопродукции. Созревание репродуктивной функции завершается к 18—20 годам.
Морфологическое созревание лобной корки достигается только к 12 годам, а окончательное формирование полушарий конечного мозга - в 20-22 года. Для пубертатного периода характерными считаются: усиление подкорковых влияний и ослабление деятельности коры головного мозга, нарушение вегетативной сферы, повышенная эмоциональность, особенно у девочек.
41. Какова была средняя продолжительность жизни первобытного человека?
Древние люди (палеоантропы) неандертальцы - Средний возраст составлял 30 лет.
Кроманьонский человек и человек современного физического типа (неоантропы) - Средний возраст кроманьонцев составлял около 30 лет.
|