Реферат
На тему:
Риниофиты. Бесполое размножение.
Спорангии и спорогенез у древних высших растений
Риниофиты
Риниофиты (Rhyniophyta), псилофиты (Psilophyta), самая древняя и примитивная вымершая группа (отдел) высших растений. Характеризовались верхушечным расположением спорангиев и равноспоровостью, отсутствием корней и листьев, дихотомическим или дихоподиальным (псевдомоноподиальным) ветвлением, примитивным анатомическим строением. Проводящая система — типичная протостела. Протоксилема располагалась в центре ксилемы; метаксилема состояла из трахеид с кольчатыми или (реже) лестничными утолщениями. Опорные ткани отсутствовали. Р. ещё не обладали способностью ко вторичному росту (меристемы у них были только верхушечные). Спорангии примитивные, от шаровидных (диаметром около 1 мм) до продолговато-цилиндрических (длиной до 12 мм), толстостенные. Гаметофиты. достоверно не известны (некоторые авторы считают гаметофитами горизонтальные корневищеподобные органы — так называемые ризомоиды). Риниофиты. произрастали на влажных и болотистых местах, а также в прибрежном мелководье. Отдел Риниофиты. включает один класс — риниопсиды (Rhyniopsida) с двумя порядками — Rhyniales (семества Cooksoniaceae, Rhyniaceae, Hedeiaceae) и Psilophytales (семейство Psilophytaceae). Для порядка Rhyniales характерны дихотомическое ветвление и тонкая, слабо развитая стела. Ксилема из трахеид с кольчатыми утолщениями. Древнейший представитель Риниофит. — род куксония, первоначально обнаруженный в Уэльсе в отложениях конца силурийского периода (около 400 млн. лет назад). Наиболее полно изучены нижнедевонские роды — риния и отчасти хорнеофит, у которого ризомоид (вверх от него отходили стебли, вниз — многочисленные ризоиды) был расчленён на чётковидно расположенные клубневидные сегменты, лишён проводящих тканей и целиком состоял из паренхимных клеток. Полагают, что в процессе эволюции ризомоиды. дали начало корням. У обоих родов стенка спорангия была многослойной, покрытой кутикулой. Хорнеофит характеризуется своеобразной спороносной полостью, которая образует купол, сводообразно покрывающий центральную колонку стерильной ткани, представляющей собой продолжение флоэмы стебля. Этим хорнеофит напоминает современный сфагнум. В семейства риниевых включают также род тениокрада, многие виды которого образовывали подводные заросли в среднем и верхнем девоне. В отдельное семейства хедеевых иногда выделяют нижнедевонские роды хедея и яравия. Нижнедевонский род сциадофит, обычно выделяемый в отдельное семейства сциадофитовых, — небольшое растение, состоявшее из розетки простых или слабо дихотомированных тонких стеблей со стелой. Для порядка Psilophytales характерны дихоподиальное ветвление и более сильно развитая стела. У наиболее известного рода — псилофит (из нижнедевонских отложений в Восточной Канаде) — неравно развитые ветви образовывали ложную главную ось дихоподия с более тонкими боковыми ветвями: стебель был окружен кутинизированной эпидермой с устьицами; поверхность стебля была голая или покрыта шипами длиной 2—2,5 мм, концы которых дисковидно расширялись, что, вероятно, указывало на их секреторную роль. Спорангии раскрывались продольной трещиной. К псилофиту близки нижнедевонские роды тримерофит и пертика.
Бесполое размножение. Спорангии
В предыдущей главе мы попытались проследить эволюцию спорангиеносных частей у архегоняальных растений. Теперь перейдем к рассмотрению самого процесса размножения. Попытаемся уловить основные моменты эволюции в развитии спорангиев и спор.
Как известно, у папоротникообразных существует два типа развития спорангия: евспорангиатный и лептоспорангиатный. При развитии спорангия по евепорангиатному типу в образовании его участвует целый участок ткани или во всяком случае группа клеток. Самый же спорангий в большинстве случаев отличается крупными размерами и толстой, многослойной стенкой.
Лептосиорангиатный тип характеризуется тем, что спорангий возникает из одной, эпидермической клетки, имеет небольшую величину и тонкую, однослойную стенку, снабженную более или менее ясно выраженным кольцом. По евепорангиатному типу развиваются спорангии у Psilotales, плаунов, хвощей, ужовников, маратциевых, у всех голосемянных. По этому же типу, вероятно, развивались спорангии у ископаемых представителей Lycopsida и Sphenopsida.
По лептоспорангиагному типу развиваются спорангии у настоящих папоротников. Примером евшорангиатного типа может служить развитие спорангия плауна — L. clavatura (рис. 69). По исследованиям П. А. Баранова (1924) спорангий L. clavatum закладывается на верхней стороне спорофилла, ближе к его основанию. Начало ему дает группа поверхностных клеток, которые уже на очень ранних стадиях несколько отличаются своим содержимым от соседних. Клетки эти делятся стенками, параллельными поверхности спорофилла, на наружные и внутренние. Из внутренних в дальнейшем образуется археопорий, из наружных — стенки спорангия. И те и другие клетки энергично делятся, в результате чего образуется бугорок. В дальнейшем в этом бугорке дифференцируется археспорий и двуслойная стенка спорангия. Клетки археспория отличаются более крупными размерами, крупными ядрами и густой протоплазмой. Между археспорием и стенкой спорангия образуется выстилающий слой — тапетум. Он отделяется от внутреннего слоя стенки спорангия, а в нижней части спорангия дифференцируется из прилежащей к археспорию ткани. Зачаток спорангия растет неравномерно: нижняя часть его отстает от верхней, в из нее образуется ножка спорангия. Клетки археспория обособляются и превращаются в материнские клетки спор, которые обычным путем дают тетрады и опоры.
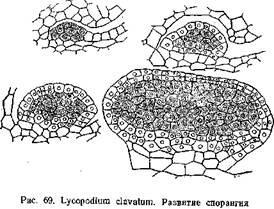
Также в общих чертах развиваются спорангии у селагинелл, хвощей и поилотов. Эволюция спорангиев выразилась в выработке приспособления для раскрывания спорангия и в возникновении особого Слоя, играющего роль в питании развивающихся спор и образовании у них сложных оболочек — тапетума или выстилающего слоя. Задачу раскрывания спорангия принимает на себя Наружный слой его стенки, превращаясь в так называемый экзотеций. Стенки его клеток утолщаются, причем утолщения эти у различных представителен папоротникообразных носят различный характер (балки, спирали и т. д.). Среди утолщенных клеток проходят ряды клеток снеутолщенными клетками (stomium). По ним проходит трещина, разрывающая стенку спорангия.
Выстилающий слой располагается между стенкой спорангия и спорогенным комплексом. Он отчленяется или от стенки спорангия (Lyoopodium) или от самого спорогенного комплекса, то есть наружные Слои этого последнего превращаются в тапетум (Ophioglossum, Equisetum). При этом различают два рода тапетума: секреторный и периплазмодиальный.
У секреторного тапетума стенки клеток сохраняются до конца его существования, и тапетум все время представляет собой слой клеток, облекающий внутреннюю поверхность спорангия. Но нередко клетки тапетума увеличиваются в размерах. При этом изменяется и характер их содержимого. Считают, что клетки секреторного тапетума выделяют теили иные вещества, которые участвуют в образовании спор и покрывающих их оболочек. Однако более подробно роль эта остается невыясненной.
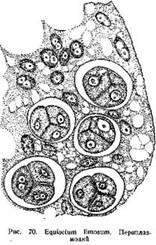
В периплаэмодиальном тапетуме стенки его клеток, иногда уже на ранних стадиях развития спорангия, расплываются к их протопласты сливаются вместе в одну сплошную массу, имеющую характер многоядерного плазмодия: его называют периплазмодием. После обособления материнских клеток спор, периплазмодий проникает в промежутки между ними, так что материнские клетки оказываются погруженными в плазматическую массу его (рис. 70). По мере развития спор и образования на них сложных оболочек периплазмодий становится все менее густым, в нем появляется много вакуолей. В конце концов, ко Времени вскрывания спорангия, периплазмодий исчезает.
Секреторный тапетум имеется у плаунов, селагинелл, у голосемянных; периплазмодиальный — у хвощей и ужовников. У псилотов также наблюдается периплазмодий, но у них он развивается своеобразно — не из тапетума, а из спорогенного комплекса. В этом последнем, на известной стадии развития спорангия наступает дифференцировка на плодущие и бесплодные клетки. Первые дают начало материнским клеткам спор и самим спорам, вторые же, после того, как разделяющие их стенки расплывутся, сливаются вместе и образуют периплазмодий.
Иной путь эволюции спорангиев выражается в слияния их между собой в синавгии. Синангии характерны для маратциевых папоротников (пор. Marattiales). При этом одни роды, как например, Angiopteris ли Archangiopteris, имеют еще отдельные спорангии, сидящие близко друг к другу. У других, как у Danaea, Christensenia или Marattia, спорангии настолько тесно срастаются, что границы отдельных, спорангиев в общем -синангии уловить нельзя. Открывается каждый спорангий отдельно продольной трещиной или круглым отверстием в общую борозду, проходящую между двумя рядами спорангиев (Marattia) или в углубление в центре синангия наблюдается и в других группах архегониат. Так оближенно сидят они, отчасти срастаясь, у Ophioglossales. У Cycas микроспоранши располагаются на нижней поверхности микро-спорофилла тесными группами. Но в наиболее, пожалуй, совершенной форме синангии представлены в ископаемой группе Whittleseyinae, относимой к Pteridospermae. У них (Christensenia) или, наконец, прямо на поверхность синангия (Danaea). Закладываются спорангии у Angiopteris отдельно, развиваясь из группы клеток У Marattia спорангии закладываются в общей плаценте, появляющейся в виде двух параллельных валиков, располагающихся вдоль жилки на нижней поверхности листа. Начало им дает целая группа клеток, которую, однако, по исследованиям Ф. Боуэра, можно свести к одной клетке.
Тенденция сливаться вместе и образовывать синангии срастаются микроопорангии, причем срастаются они настолько полно, что получается цельное тело, внешне напоминающее семя ли шлод.
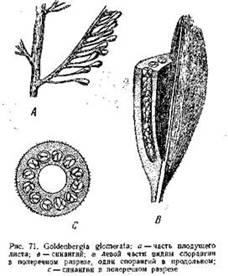
У Goldenbergia glomerata, например, (рис. 71) такой плод представлял собой овальное тело около 6—8 -мм длиной, сидящее на длинной тонкой ножке (10—15 мм). Внутри •его находилась обширная полость, а стенка была построена из вегетативной ткани, в толще которой заключалось 12—18 трубчатых спорангиев. Они располагались в один ряд вокруг центральной полости и содержали в себе довольно крупные микроспоры (0,3—0,4 мм). Внешняя поверхность плода была покрыта короткими шипиками.
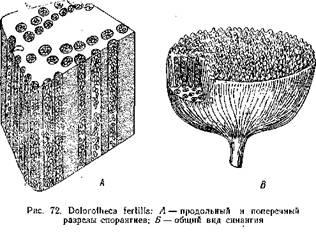
Подобное же строение имели и синангии Aulotheca. У Whittleseya elegans синангии имел форму бокала на тонкой ножке. Стенку бокала образуют сросшиеся между собой трубчатые спорангии с очень толстыми стенками. Dolorotheca мела чашевидные синангии, причем полость чаши была заполнена вегетативной тканью, в которую были вкраплены трубчатые спорангии (рис. 72). Т. Галле, подробно изучивший плодоношение Whittleseyinae, считает, что синангии их могли возникнуть путем тангентальвого сращения трубчатых спорангиев, подобно тому, как листья могли возникнуть ив дихотомически разветвленных теломов благодаря срастанию и с краями в одной плоскости.
Ход эволюции синангиев Whittleseyinae Т. Галле иллюстрирует следующей схемой (рис. 73). На ней первая фигура изображает гипотетическую исходную форму — плодущий синтелом. Наличие групп спорангиев Psilophyton, сидящих на концах теломов, подтверждает построение такой гипотетической формы. На следующей стадии спорангии срослись в нижней своей половине. Вершины их остаются свободными.
Это плодоношение типа Codonotheca. Далее срастание спорангиев идет до их вершин, получается бокаловидный синангии, как у Whittleseya elegans. Наконец, спорангии смыкаются и срастаются своими вершинами —образуется плод, полый внутри со спорангиями, находящимися в его стенке—Aulotheca, Goldenbergia.
Лептоспорангиатный тип развития спорангия характеризует типичные папоротники (пор. Filioales). Он отличается тем, что спорангий здесь возникает из одной эпидермической клетки, и сам спорангий имеет однослойную стенку и относительно малые размеры. Развитие его происходит по следующей схеме (рис. 74).

Клетка эпидермиса плаценты разрастается, выпячивается над общим уровнем эпидермиса и отделяется поперечной перегородкой от остальной ткани плаценты. Эта отделившаяся клетка и дает начало всему спорангию. В ней появляется горизонтальная перегородка, разделяющая ее на 2 клетки: нижнюю — меньшую, плоскую и верхнюю — большую, выпуклую. Нижняя в дальнейшем образует ножку спорангия, из верхней же развивается самый спорангий. В верхней клетке появляются последовательно три косые перегородки, пересекающие друг друга под некоторым углом. Первая перегородка проходит почти вертикально под небольшим углом к горизонтальной стенке, вторая пересекает под острым углом первую, а третья (на рисунке невидимая) пересекает тоже под острым углом две первые. Эти перегородки разделяют одноклетный зачаток спорангия на четыре клетки: одна из них занимает верхушку его и имеет форму трехгранной пирамиды, обращенной вершиной вниз, а выпуклым основанием наружу; три остальные лежат ниже ее, примыкая к ее граням. Далее, в пирамидальной клетке образуется горизонтальная перегородка.

Теперь весь зачаток состоит из пяти клеток— одной пирамидальной, внутренней, и четырех наружных, окружающих ее. Внутренняя клетка является археспориальной клеткой, и в нее развивается содержимое спорангия — опоры и выстилающий слой. Наружные же дают начало стенке спорангия. Они делятся только радиальными перегородками, так что стенка спорангия все время остается однослойной. В археспориальной же клетке появляются перегородки, тангенгальные, параллельные ее граням. Отделившиеся таким образом четыре таблитчатые клетки дают начало тапетуму, внутренняя же является опорообразующей, из нее развиваются споры. Клетки тапетума снова делятся тангентальными перегородками, вследствие чего он становится двуслойным. В то же время делится и опорообразующая клетка, постепенно распадаясь на 16 клеток. Вскоре стенки клеток тапетума расплываются и протопласты их сливаются, образуя периплазмодий. Клетки же археспория обособляются друг от друга и округляются, превращаясь в материнские клетки опор, которые обычным путем дают начало спорам.
Евопорангиатный тип спорангиев встречается у наиболее древних и примитивных групп среди ныне живущих папоротникообразных. Он же характерен) и для ископаемых папоротникообразных. Это заставляет нас смотреть на евшорангий как на древний, примитивный тип спорангия, от которого уже произошел лептосггорангий, свойственный к тому же более молодым группам папоротников (Filices). Об этом говорит и то, что, как показывают сравнительные исследования, между евопорангиатный я лептоспорангиатным типами развития существует ряд постепенных переходов, связывающих их между собой. Вместе с тем эти переходы указывают на ход эволюции лептоспорангия.
На рис. 75 дана схема заложения опорангия у различных папоротников: рис. а относится к типичному лаптоспорангию (сем. Polypodiaeeae), а рис. g— к типичному евспорангию (Angiopteris). Между ними располагаются промежуточные типы, принадлежащие к различным родам. Понятно, эволюция лептоспорангия совершалась в направлении, обратном расположению рисунков. У Angiopteris археспорий закладывается глубоко под поверхностью и самая археспориальная клетка имеет форму куба: ограничивающие ее стенки пересекаются под прямым углом. У Todea (сем. Osmundaceae) наблюдаются два варианта развития опорангия. В одном случае ([) археспорий имеет в разрезе трапециевидную форму, благодаря тому, что антиклины, ограничивающие археспорий, наклонены под некоторым углом друг к другу. В другом случае (более мелкие спорангии) эти антиклины пересекаются, и археспорий принимает форму четырехгранной пирамиды, обращенной вершиной вниз, то есть форму, характерную для археспория лептоепорангия. И у Angiopteris и у Todea в образовании спорангия принимают участие и прилежащие к археспорию клетки. Развитие спорангия у Schiziaea (d), Alsophila(с) и Ceratopteris (Ь) представляет переход к типичному лептоспорангиатному типу (а). Археспорий окончательно принимает форму четырехгранной пирамиды и все меньшее число эпидермических клеток идет на развитие спорангиев. У Polypo-diaceae (а) весь спорангий развивается из одной эпидермической клетки.
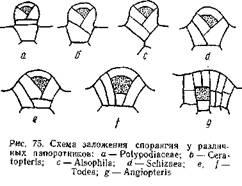
Таким образом, эволюция спорангия у папоротников шла в направлении редукции, которая и приведет к образованию лептоспорангия. Эволюция последнего, кроме того, выразилась в создании длинной и тонкой ножки, выработке кольца и в общем уменьшении количества производимых спорангием спор. В результате получился типичный лептоспорангий малой величины с длинной ножкой и однорядным кольцом. Число спор, образующихся в спорангии, невелико, но число спорангиев сильно возрастает, благодаря чему общее количество спор, производимых растением, не только не уменьшается, но и весьма увеличивается.
По характеру спор среди археголиальных мы различаем две труппы растений — равноепоровые и разноюлоровые (гомо- и гегероспоровые) формы. И те и другие встречаются во всех классах папоротникообразных; голосемянные—исключительно тетероспоровые. Это показывает, что гетероспория возникала в разных типах и классах самостоятельно я независимо. Естественно встает вопрос, какой тип спор мы должны считать первоначальным, исходным, иначе, развилась ли гетероспория из гомоспории или, наоборот, гетероспория дала начало гомоспории. Прямых указаний на то или иное решение вопроса мы не имеем, теоретически же возможно допустить и тот и другой ход эволюции. Однако, исходя из того факта, что гетероспория всегда связана с сильной редукцией гвметофита, в принимая во внимание то, что примитивный гаметофит должен был быть хорошо развитым образованием, правильнее будет предположить, что гомоопория является примитивным признаком и первоначально все опоры были гомоспорами, то есть при прорастании давали обоеполые заростки, несущие и антеридии и архегонии. В дальнейшей эволюции произошла дифферевцировка их на микроспоры и мегаспоры — мужские и женские опоры. Указание на такого рода дифференцирозку мы находим и среди мхов, согласно исследованиям сравнительно недавнего времени.
У Utota bruchii, например, величина спор, образующихся в одном и том же спорангии, сильно колеблется (от 21 до 32 (х). Однако связана ли эта разница в величине с половой дифференцировкой, остается невыясненным. Но для форм, близких к Ulota bruchii, из того же семейства Orthotriehaeeiae, именно для Macromitrium blumeivH Schlotheimia konigsbergii таковая дифференциревка была доказана М. Ф. Фдейшером. Споры у этик мхов также сильно разнятся в размера и, как показал М. Флейшер, яри ирорастании их из более крупных спор вырастают всегда карликовые мужские растения. Из мелких же спор развиваются нормальные женские особи.
М. Флейшер нашел также карликовые мужские и женские растения у Trismegistia brauniana (из Hypnobryales), хотя споры у этого мха внешне не отличаются друг от друга по размерам. Внутренняя половая дифференцировка, таким образом, у них имеется. Карликовые мужские растения Флейшер нашел и у многих других мхов.
Позднее гетероспория была обнаружена М. Эрнет-Шварценбах (Ernst-Schwarzenbach,1939) у ряда видов Macromitrium. Этим автором было исследовано 35 видов p. Macromitrium. Из них 16 видов оказались гомостюровыми, 14— явно гетероспоровыми, 5—неясно гетероспоровыми. Например у salakense микроспоры имеют в- среднем 19,6 ц в диаметре, мегаспоры — 36,7 ц. Микроспоры имеют желтую окраску и одеты гладкой оболочкой, мегаспоры — зеленые, оболочка их — бородавчатая. Отношение тех и других— 1 : 1. На выросшей из микроспор протонеме через 3—4 месяца после посева образовались карликовые растения с антеридиями. На протонеме, развившейся ив мегаспор, появились Стебля, но к моменту окончания опытов половых органов на них еще не было. На них, по-видимому, при дальнейшем развитии -должны были появиться архегонии.
Как видно из сказанного, между данными М. Флейшера и М. Эрнст-Шварценбах имеется известное противоречие. По М. Флейшеру, микроспоры являются женскими спорами, мегаспоры — мужскими, по М. Эрнст-Шварценбах, наоборот, то есть мхи ведут себя в этом отношении так же, как папоротникообразные. Вопрос о гегероспории у мхов требует, следовательно, дальнейшего исследования. Однако и в настоящий момент мы можем констатировать, что у лиственных мхов гетероспория в той или иной форме существует. Указаний на гетероепорию у печеночников нет.
Большинство гомоепоровых папоротникообразных обладает обоеполыми заростками. Это указывает на то, что в спорах их нет половой дифференцировки и все споры и внешне и внутренне вполне равноценны. Однако зачатки гегероспории мы можем уловить и у них. Так, по свидетельству К. Гебеля, споры типичного папоротника Dennstedtia pimctilobula при посеве дают однополые заростки, то есть часть их развивается в мужские, часть в женские заростки. То же самое наблюдается и у некоторых хвощей, как например, у Equisetum limosum, E. hiemale, E. silvestre. Споры стих. хвощей дают мужские и женские заростка, отличающиеся друг от друга размерами, строением и даже окраской. Мужские заростки — меньшей величины, более - простого строения по сравнению с женскими и более светлой, желтоватой окраски. И у Dennstedtia, и у названных выше хвощей пол споры, возможно, определяется генотипически, так что если все споры -по величине и форме одинаковы, то внутреннее их различие — весьма резкое. У других хвощей, как у Е. debile, E. laevigatum или Е. arvense, заростки — обоеполые, причем архегонии на них развиваются раньше антеридиев. Но наряду с обоеполыми у этих хвощей образуются и чисто мужские заростки; количество их весьма зависит от тех условий, при которых прорастают споры и развиваются заростки.
В этом отношении подробно -изучен Е. arvense E. Шратдем (Schratz, 1928). Названный автор исследовал, насколько внешние условия определяют у Е. arvense пол заростка. Результаты его исследования сводятся к следующему. При неблагоприятных условиях, например при очень густом посеве большинство заростков становится чисто мужскими. Отсюда можно сделать вывод, что все споры могут дать мужские заростки и что пол заростков определяется внешними условиями. При благоприятных условиях получается не менее 50%-мужских заростков. Остальные — сначала являются женскими, впоследствии же становятся обоеполыми: на них появляются антеридии.
Переход женских заростков в обоеполые особенно хорошо происходит тогда, когда не бывает оплодотворения, и жизнь заростка тем самым затягивается. Ухудшая условия существования, можно заставить женские заростки образовать антеридии. Заставить же женские заростки, улучшая внешние условия, перейти к образованию архегониев не удается. Отношение мужских заростков к женским (соответственно обоеполым), как 1:1, -наблюдается и среди заростков, находимых в природе.
Вое это позволяет думать, что однодомность является нормальным распределением полов у Е. arvense; если не удается все заростки довести до .обоеполого состояния, то это зависит от того, что пол заростка -определяется очень рано, при развитии спор, и раз определившись, -он сохраняется уже и в дальнейшей жизни заростка, не изменяясь. Интересно отметить, что в спорангиях некоторых видов ископаемого рода Calomosta-chys, наблюдались споры различной величины, что позволяет допускать у них фенотипическое определение пола.
Таким образом, у одних хвощей (Е. arvense, E. debile в др.) половая дифференцировка спор еще не установилась, и определение их пола находится в сильной зависимости от внешних условий, у других же видов (Е. limosum, E. hiemale и др.) половая дифференцировка уже совершилась и можно говорить о мужских и женских спорах. Гетероспорией эту дифференцировку назвать еще нельзя, так как внешнего различия между опорами здесь нет. Поэтому многие исследователи называют ее гетероталлизмом.
Ф. Жуйе-Лавернь показал, что споры хвощей различаются по своему химическому составу. Применяя различные прижизненные окраски: нейтральрот, крезильблау, митиленовую синьку, а также обрабатывая споры хвощей осмиевой кислотой и некоторыми другими реактивами, он обнаружил среди спор хвощей две группы, -отличающиеся друг от друга величиной окислительно-восстановительного потенциала — гН: одни из них — группы А, — имеют более низкий гН, чем другие (группы В). Исходя из того факта, что семяпочки покрытосемянных ведут себя так, как споры группы А, а пыльцевые зерна, как опоры группы В, Ф. Жуйе-Лавернь делает заключение, что споры группы А являются женскими, а группы В—мужскими опорами. Однако Е. Шратц не подтвердил результатов Ф. Жуйе-Лавернь.
Гетероталлизм в дальнейшей эволюции влечет за собой адтероспорию: споры начинают различаться между собой не только то внутреннему строению, но и внешнему — по величине, по строению оболочек и т. д. При этом мужские споры всегда гораздо мельче и проще устроены, чем женские. Вместе с тем гетероспория всегда, как это показывает изучение разноспоровых форм, связана с редукцией гаметофита. Редукции особенно сильно подвергается мужской гаметофит. Меньшая величина микроспор и большая редукция мужского гаметофита стоит, по-видимому, в связи с кратковременностью существования и функционирования его, с коротким сроком, в который совершается половой акт. Для развития немногих сперматозоидов, образующихся из микроспоры, не требуется .большого количества питательных веществ и отпадает вместе с тем необходимость в вегетативной ткани заростка; Немногочисленность Сперматозоидов, даваемых каждой микроопорой, восполняется большим числом микроспор, так что .общее количество сперматозоидов, образуемых особью, громадно и вполне обеспечивает половой процесс. Сильную редукцию мужского гаметофита мы видим у всех разноспоровых папоротникообразных. У оелагинелл, Isoetes, и водных папоротников вегетативная часть мужского заростка сводится, в сущности, к двум клеткам—ризоидальной и базальтом. Кроме них мужской заросток содержит один (Isoetes) или два антеридия (оелагинелли, водные папоротники), дающие немногие сперматозоиды. В еще большей степени редуцирован мужской гаметофит у голосемянных и покрытосемянных. У голосемянных при прорастании микроспор образуется одна (Cyoadales) или две Вегетативные клетки мужского заростка (Ginkgo, хвойные). Они очень скоро разрушаются и ко времени образования половых элементов от них не остается ни следа. Остальные клетки мужского заростка (клетка-ножка, клетка, дающая пыльцевую трубку) надо считать принадлежащими к антеридию.
У покрытосемянных же стерильных клеток мужского заростка совсем не образуется, имеется лишь клетка, вырастающая в пыльцевую трубку; ее следует считать принадлежащей антеридию. Но самый микроспорангий при этом не подвергся никакой особой эволюции, сохраняв свой характер или лептоспорангия (водные папоротники), или евспорангия (селагинелли, голо- и покрытосемянные).
Мегаспоры (женские споры) много крупнее микроспор и имеют более сложное строение, обладая толстыми, нередко многослойными оболочками со сложной структурой. У папоротникообразных, кроме того, в мегаспорах откладываются в большем или меньшем количестве запасные питательные вещества, за счет которых развивается женский заросток и зародыш на первых стадиях своей жизни. У голосеменные, у которых мегаспора не покидает мегаспорангия, питание развивающемуся заростку и зародышу доставляет спорофит.:
Женский заросток редуцирован значительно меньше, чем мужской. Он почти всегда представляет собой тканевое образование, и связан с оболочкой мегаспоры. Женский заросток или имеет вид небольшой зеленой пластинки (Salvinia, Azolla) или образует комплекс тонкостенных, плотно соединенных между собой клеток, заполняющих полость мегаспоры (селагинелла, Isoetes, голосемянные). Наибольшей редукции подвергся женский заросток у Marsiliaceae, где он сводится к двум-трем клеткам и одному архегонию.
Самый мегаспорагий у папоротникообразных не претерпел сложной эволюции и сохранил характер спорангия (ев спорангия или лептоспорэнгия), свойственный той или иной их группе. У семенных же растений (голо- или покрытосемянные) мегаспорангия превратился в семяпочку, пройдя сложную эволюцию в связи с тем, что мегаспора стала развиваться внутри мегаспорангия на Материнском растении и тем, что мегаспорангий был Образованием, из которого возникло ceмя. С семяпочками мы впервые встречаемся у семенных папоротников.
Во всех отделах голосемянных растений семяпочки построены в общем по одному плану. Семяпочка состоит из шуцеялуса, образующего центральную часть ее. Внутри нуцеллуса помещается эндосперм, построенный из тонкостенных, плотно соединенных между собой клеток. В эндосперме находятся архегогии. Снаружи нуцеллус одет покровом или интегументом, имеющим на вершине микропиле или семявход. -
Еще исследованиями В. Гофмейстера (1851) доказано, что нуцеллус морфологически представляет собой мегаспорам, а эндосперм — женский заросток, который развился внутри мегаспорангия из мегаспоры. Эта последняя здесь не высеивается из мегаспорангия, а остается внутри его, тут же про растает в заросток, который всю жизнь связан с спорофитом паразитируя, так сказать, на нем.:
Что же касается интегумента, то он является новым образованием, не имеющим гомолога у папоротникообразных. Но если ясна морфологическая природа нуцеллуса и эндосперма, то нельзя того же сказать про интегумент. Морфологическое значение и происхождение покрова остается неясным, и вполне возможно, что интегументы в разных порядках голосемянных растений не гомологичны.
В. Циммермавн приписывает интегументу листовое происхождение, думая, что он возник из сросшихся между собой в виде чашечки филлоидов. В пользу этою говорит, по его мнению, то, что у Lyginopteris, например интегумент не срастается с нуцеллусом, а свободно облегает его; далее у некоторых птеридоспермов, в особенности у Physostoma elegans, сосудистые пучки, пронизывающие интегумент, свободно оканчиваются в нем так же, как это наблюдается в чашечке. Однако отношение этих филлоидов к спорофиллам остается неясным.
Р. Пильгер (1927) .по поводу интегументов цикадовых говорит, что многое указывает на то, что толстый интегумент, свойственный этой группе, произошел из отдельных, сросшихся по длине частей, морфологическая природа которых неясна. Так, у Macrozamiia после удаления внешнего, сочного слоя интегумента можно видеть, что мииропиле окружено 8—И ребрами, разделенными короткими, глубокими ложбинками; у Geratoaamia и Encephalartos по каменистому слою семени проходят от основания его до микропиле хорошо выраженные ребра. В пользу того же предположения говорит и строение семян некоторых семенных папоротников, как например Physostoma elegans, упомянутой выше. Семена ее достигают 6 мм в длину и 2 мм в поперечнике. По оболочке вдоль их проходят 10 ребер, переходящие в свободные лопасти, окружающие венчиком верхушку нуцеллуса. Они играют роль микропилярной трубочки. Сосудистый пучок, войдя в халазу, дает десять ветвей, из которых каждая входит в ребро и продолжается в лопасть. Интегумент и нуцеллус срастаются почти до того места, где начинаются свободные лопасти покрова.
Оригинальный взгляд на происхождение интегумента голосемянных был высказан М. Бенсон (Benson, 1904). На основании изучения семян Telangium, она пришла к выводу, что семяпочка семенных папоротников возникла из еинангия. Произошло это таким образом, что расположенный в центре еинангия мегаспор авгий сохранил свою функцию и превратился в нуцеллус. Окружающие же его мегаспорангии стерилизовались и, срастаясь между собой, образовали интегумент. Этот последний является, таким образом, гомологом мегаопорангиев, а в конечном счете плодущих теломов В. Циммерманна.
К аналогичному выводу пришел С. Уольтон (Walton, 1953). Он на основании сравнительно морфологического анализа семян Pteridospermae считает, что интегумент и плюска семенных папоротников произошли из некоторого числа листовых образований путем их срастания между собой и обрастания ими нуцеллуса.
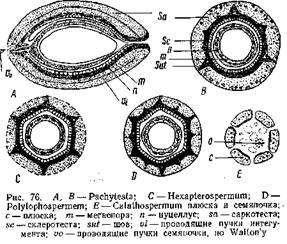
При этом интересно отметить, что число этик образований и в группе Lagenostomales и Trigonocarpales обычно кратно трем. У Calathospermqm плюсна состоит из 6 отдельных частей, окружающих мегаспорангии. У Hexapterospermum и Polylophospermum части эти срастаются и образуют общий покров, достигающий значительной толщины и дифференцирующийся натри Слоя — наружный мясистый (sarcotesta), второй — каменистый (solerotesta) и внутренний — тонкостенный. В местах сращения на склерютесте возникли ребра, вдающиеся в саркотесту (рис. 76). Этот покров семян Trigonacarpales соответствует плюске- Lagenostoma, а собственно интегумент, хорошо выраженный у Lagenostoma и свободно лежащий в плюске, в семенах Trigonocarpus срастается с сильно редуцированным нуцеллусом, образуя то, что называют у него нуцеллусом. И у Trigonocarpus и у Lagenostoma в семени находится только один мегаспорангии. Таким образом, Trigonocarpus по сравнению с Lagenostoma отличается более простым строением, и оба он более просты по сравнению с Caiiathospermum, у которого в купле находится несколько мегаслорангиев. Эволюция в семенах Pteridospermae шла, по мнению С. Уольтона, в направлении редукции числа мегаепорамгиев и упрощения строения их. Это упрощение выражается и в устройстве вершины нуцеллуса, довольно "сложного у Lagenostoma.
Выше изложенные соображения о возникновении интегумента у цикадовых могут быть так или иначе приложены и к хвойным. Однако в последнее время О. Гагеруп (Hagerup, 1933) выдвинул новое толкование интегумента у хвойных. На основании весьма подробного изучения истории развития женских шишек у ряда хвойных различных семейств он пришел к выводу, что интегумент представляет собой мегаепорофилл, несущий мегаопорангий, то есть пуцеллус с заключающейся в нем мегаспорой, а впоследствии — женским заростком. Это заключение подтверждают и явления позеленения семяпочек, наблюдаемые на женских шишках у многих хвойных. В таких случаях на месте семяпочек появляется вегетативный" лист (хвоя). Кроме того, можно встретить и переходы между хвоей в покровом семяпочки.
Напомним, что к такому же толкованию интегумента у кордаитов и хвойных пришел Р. Флорин (1951), на основания изучения ископаемых голосемянных. Речь об этом была выше, когда мы разбирали морфологию женских шишек у кордаитов и хвойных. У них, по толкованию Р. Флорина, интегумент является гомологом двух конечных лопастей мегаспорофишла, между которыми помещался мегагаюрангий. Но у тиссовых интегумент возник путем срастания двух или нескольких чешуи, соответствующих по своему положению мегаспорофиллам.
Неясным остается и Происхождение покровов у покрытосемянных растений. История развития не дает нам никаких указаний на филогению интегумента. По данным же сравнительной морфологии, согласно воззрениям одних авторов, они стеблевого (осевого) происхождения, по взглядам других — листового.
В пользу последнего взгляда, развиваемого, между прочим, А. Кернером (1900) говорят, прежде всего, явления озеленения семяпочек — превращения в листообразный орган внешнего интегумента. А. Кернер в позеленевших цветках Primula japonica нашел семяпочки, у которых внешний покров принял форму листа. Па нем помещался бугорок, зачаток нуцеллуса, а вокруг этого рудиментарного нуцеллуса находился кольцеобразный выроет, соответствующий внутреннему покрову. То же самое наблюдалось и у Trifolium repens, Rumex scutatus. Здесь также внешний покров превращается в лист, на котором сидит рудимент нуцеллуса с рудиментарным же внутренним покровом. И. Лотси у Hesperis находил семяпочки с превращенным в лист наружным покровом: на жилках его помещались зачаточные внутренние интегументы.
Эти факты подтверждают, листовое происхождение внешнего покрова. Но если это так, то внешний покров у покрытосемянных представляет собой мегасворофилл, несущий мегаопорангий (нуцеллус). Внутренний же интегумент —новообразование, вырост мегаспор ангия. Не будучи .генетически связан с листом, он листовидной формы при антолизе не принимает. В случае одною покрова семяпочки, можно принять, что он гомологичен мегаепорофиллу. Внутренний же покров или не развился или редуцировал. Но возможно, что произошло сращение обоих покровов в один.
Таким образом, для покрытосемянных мы получаем то же толкование природы интегумента, какое дают О. Гагеруп и Р. Флорин покрову голосемянных. Вместе с тем с высказанной выше точки зрения внешний и внутренний покровы семяпочки покрытосемянных не гомологичны.
Каково бы ни было происхождение интегументов, они представляют собой новое образование, свойственное только семенным растениям и возникшее в связи с образованием семян. Значение их сводится на ранних стадиях к защите нуцеллуса. Впоследствии у них развиваются покровы семени, защищающие находящийся внутри семени зародыш, а в некоторых Случаях способствующие распространению семян, как например сочные покровы семян Цикадовых и др. Интересно отметить, что наибольшего развития, наибольшей толщины интегументы достигают там, где семяпочки слабо защищены спорофиллами или другими образованиями (чешуями и. пр.). Так, у Cycas и других Cycadales, у Podocarpus, Taxus, покровы отличаются толщиной и нередко становятся сочными. У сосновых же, где семяпочки спрятаны среди семенных чешуи, покровы тонкие. Тонкие покровы и у Cypressaeeae и Taxodiaceae. Тонкими же относительно интегументами характеризуются и семяпочки покрытосемянных растений.
Спорангии и спорогенез у древних высших растений
У многих древних высших растений спорангии образовывались верхушкой побега (как у
мхов). Позже спорангии стали формироваться также на листьях, что и наблюдается почти у всех современных ныне живущих высших растений, за исключением мхов и некоторых
плаунов. Листья, на которых развиваются спорангии, получили название
спорофиллов . Они обычно собраны в колосья, так называемые
стробилы , и часто значительно видоизменены по сравнению с обычными листьями растения. Очень редко, например у
псилотовых и
саговников, спорангии срастаются по три в сложные спорангии -
синангий. У псилотовых синангий располагаются поодиночке с верхней стороны чешуевидного вильчато-разветвленного
листа
. У саговников они собраны в большом числе на нижней стороне чешуевидных видоизмененных листьев, образующих стробил .
У плаунов и
хвощей спорофиллы собраны в более или менее компактные группы, чаще всего в
колоски , или стробилы , и несут от 1 до 14 спорангиев. У плаунов спорофиллы часто сохраняют большее или меньшее сходство с вегетативными листьями, а у хвощей они видоизменены в щиток, или
спорангиофор, на нижней стороне которого и прикрепляются мешковидные спорангии. У ближайших родственников плаунов -
селягинелл и
полушников наблюдается разноспоровость, когда в одних спорангиях образуются
микроспоры, а в других -
мегаспоры. Такие спорангии называются соответственно
микроспорангиями и
мегаспорангиями, а спорофиллы, на которых они развиваются, -
микроспорофиллами и
мегаспорофиллами . Если в микроспорангии образуется множество мелких спор, то в мегаспорангии развивается чаще всего лишь 4 крупные мегаспоры. На гаметофите, развивающемся из мегаспор, образуются женские
гаметангии -
архегонии, а гаметофит, развивающийся из микроспоры, образует обычно только
антеридии. Гаметофит равноспоровых растений чаще всего обоеполый. Встречается разноспоровость и у некоторых
папоротников, хотя в подавляющем большинстве это равноспоровые организмы. Спорангии папоротников развиваются на нижней стороне листа. Спорофиллы часто совсем не отличаются от обычных листьев, но иногда они высоко специализированы и даже утрачивают ассимиляционную функцию. Между этими крайними примерами наблюдаются различные промежуточные варианты.
Спорангии у папоротников обычно собраны группами, расположены линейно или покрывают полностью определенные участки листа с его нижней стороны. Более или менее компактные собрания спорангиев называют
сорусами. Сидят спорангии обычно на толстом выросте - плаценте, или ложе соруса, из которого часто развивается
покрывальце, или индузий. Покрывальце бывает различной формы и несет защитную функцию.
Спорангии большинства папоротников имеют механизм раскрывания в виде кольца толстостенных клеток, идущих по гребню сжатого с боков спорангия. При подсыхании это кольцо из-за неравномерного высыхания клеточных оболочек распрямляется и разрывает тонкие стенки спорангия, освобождая споры. Строение спорангиев и сорусов у папоротников очень разнообразно и в деталях может сильно различаться.
Вывод
Изучение строения Риниофит и их эволюционных взаимоотношений имеет большое значение для эволюционной морфологии и филогении высших растений. По-видимому, первоначальным органом спорофита высших растений был дихотомически ветвящийся стебель с верхушечными спорангиями; корни и листья произошли позже спорангия и стебля. Есть все основания считать Риниофиты исходной предковой группой, от которой произошли мохообразные, плауновидные, хвощевидные и папоротники. Согласно другой точке зрения, мохообразные и плауновидные имеют лишь общее происхождение с Риниофит.
Литература
1. Основы палеонтологии. Водоросли, мохообразные, псилофитовые, плауновидные, членистостебельные, папоротники, М., 1963; Traite de paleobotanique, t. 2, Bryophyta. Psilophyta. Lycophyta, P., 1967.
|