МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РОССИЙСКОЙ
ФЕДЕРАЦИИ
ФГОУ ВПО «ДАЛЬНЕВОСТОЧНЫЙ ГОСУДАРСТВЕННЫЙ АГРАРНЫЙ
УНИВЕРСИТЕТ»
КУРСОВАЯ РАБОТА
По дисциплине: общая биология.
Тема: Эволюция генома
Выполнил: студент 2 курса, ИЛ
Гр. 8217
Черкасов Г.А.
Проверил: Красавина А. А.
Благовещенск, 2009
Содержание
Введение
1. Эволюция прокариотического и эукариотического генома
1.1 Геном предполагаемого общего предка про- и эукариот
1.2 Эволюция прокариотического генома
1.3 Эволюция эукариотического генома
2. Роль и мобильных генетических элементов в эволюции генома
2.1 Мобильные генетические элементы, их свойства
2.2 Отбор, температурные воздействия и динамика рисунка локализации МГЭ в геноме
2.3 Гипотеза: эволюционная роль системы МГЭ в геномах эукариот
2.4 Макроэволюция Aiu-подобных повторов
3. Роль горизонтального переноса генетического материала в эволюции генома
Заключение
Список использованной литературы
Введение
Геномика - наука, которая изучает структурно-функциональную организацию генома, представляющего собой совокупность генов и генетических элементов, определяющих все признаки организма. Термин геномика появился в 1987 году, а первый полный бактериальный геном был расшифрован в 1995 году. За прошедшие 14 лет определена нуклеотидная последовательность десятков
геномов прокариот и эукариот и стало очевидным, что традиционные представления о филогении, базирующиеся на дарвинистических представлениях о дивергенции недостаточны для понимания генеалогии видов. Филогенетические деревья, построенные на основе молекулярного анализа гомологии отдельных генов, в частности, кодирующих рибосомальные РНК, позволяют судить о степени генетического родства организмов, но не дают однозначного ответа о происхождении и эволюционной судьбе многих генов. Дело в том, что "ветвление" таких деревьев по схеме бифуркации отражает только принцип вертикальной эволюции, в ходе которой наследуется базовый набор предковых ортологичных генов (сходных у разных организмов). В результате дупликаций, мутаций и рекомбинаций из них образуются паралогичные гены, (сходные в составе одного
генома), которые увеличивают белковый арсенал и диапазон фенотипических вариаций, отражающих усложнение клеточных систем и регуляторных механизмов. В этом суть вертикальной эволюции "вверх". Другое ключевое направление эволюционного процесса связано с горизонтальным (латеральным) переносом генов между организмами, как близкородственными, так и филогенетически отдаленными, принадлежащими даже к разным царствам.
Цель данной работы:
- рассмотреть возможный механизм эволюции прокариотического эукариотического геномов;
- рассмотреть роль и мобильных генетических элементов и горизонтального переноса генетического материала в эволюции генома;
1. Эволюция прокариотического и эукариотического геномов
1.1 Геном предполагаемого общего предка про- и эукариот
Общие принципы организации наследственного материала, представленного нуклеиновыми кислотами, а также принципы записи генетической информации у про- и эукариот свидетельствуют в пользу единства их происхождения от общего предка, у которого уже была решена проблема самовоспроизведения и записи информации на основе репликации ДНК и универсальности генетического кода. Однако геном такого предка сохранял большие эволюционные возможности, связанные с развитием надмолекулярной организации наследственного материала, разных путей реализации наследственной информации и регуляции этих процессов.
Многочисленные указания на различия в организации генома, деталях процессов экспрессии генов и механизмов ее регуляции у про- и эукариот свидетельствуют в пользу эволюции названных типов клеток по разным направлениям после их дивергенции от общего предка.
Существует предположение, что в процессе возникновения жизни на Земле первым шагом явилось образование самовоспроизводящихся молекул нуклеиновых кислот, не несущих первоначально функции кодирования аминокислот в белках. Благодаря способности к самовоспроизведению эти молекулы сохранялись во времени. Таким образом, первоначальный отбор шел на способность к самосохранению через самовоспроизведение. В соответствии с рассмотренным предположением позднее некоторые участки ДНК приобрели функцию кодирования, т.е. стали структурными генами, совокупность которых на определенном этапе эволюции составила первичный генотип. Экспрессия возникших кодирующих последовательностей ДНК привела к формированию первичного фенотипа, который оценивался естественным отбором на способность выживать в конкретной среде.
Важным моментом в рассматриваемой гипотезе является предположение о том, что существенным компонентом первых клеточных геномов была избыточная ДНК, способная реплицироваться, но не несущая функциональной нагрузки в отношении формирования фенотипа. Предполагают, что разные направления эволюции геномов про- и эукариот связаны с различной судьбой этой избыточной ДНК предкового генома, который должен был характеризоваться достаточно большим объемом. Вероятно, на ранних стадиях эволюции простейших клеточных форм у них еще не были в совершенстве отработаны главные механизмы потока информации (репликация, транскрипция, трансляция). Избыточность ДНК в этих условиях создавала возможность расширения объема кодирующих нуклеотидных последовательностей за счет некодирующих, обеспечивая возникновение многих вариантов решения проблемы формирования жизнеспособного фенотипа. [1]
1.2 Эволюция прокариотического генома
По мере совершенствования и повышения надежности главных механизмов потока информации значение избыточной ДНК в повышении выживаемости организмов снижалась. В такой ситуаций одним из возможных направлений изменения генома было уменьшение его размеров за счет утраты некодирующих нуклеотидных последовательностей. Именно так можно представить эволюционный путь, пройденный геномом современных прокариот. Одновременно в качестве механизмов, поддерживающих выживаемость этих форм, в историческом развитии закреплялось свойственное им короткое время генерации, т.е. интенсивное размножение и быстрая смена поколений (кишечная палочка делится каждые 20 мин). Перечисленные особенности хорошо сочетаются с гаплоидностью прокариот, что приводит к воспроизведению в фенотипе любой мутации.
Экспрессия 95% ДНК, относительно малые размеры генома, гаплоидность, проявление в фенотипе практически каждой мутации в сочетании с коротким временем генерации обусловливают высокую приспособленность. Вместе с тем для прокариотического типа организации не свойственны обширные и разнообразные изменения структуры. Вследствие этого описанное направление эволюции, обеспечивая высокую способность к выживанию (прокариоты существуют на Земле около 3,5 млрд. лет), является тупиковым в плане прогрессивной эволюции живых существ. [1]
1.3 Эволюция эукариотического генома
В отличие от изменений прокариотического генома преобразования генома в эволюции эукариот связаны с нарастающим увеличением количества ДНК. Это увеличение наблюдается в процессе прогрессивной эволюции эукариот . На фоне такого увеличения большая часть ДНК является «молчащей», т.е. не кодирует аминокислот в белках или последовательностей нуклеотидов в рРНК и тРНК. Даже в пределах одного гена молчащие (интроны) и кодирующие (экзоны) участки могут перемежаться. В составе ДНК обнаруживаются высоко и умеренно повторяющиеся последовательности. Вся масса ДНК распределена между определенным числом специализированных структур — хромосом. Хромосомы в отличие от нуклеоида прокариот имеют сложную химическую организацию. Эукариоты в большинстве случаев диплоидны. Время генерации у них значительно больше, чем у прокариот. Отмечаемые особенности, оформившиеся в ходе эволюции генома эукариот, допускают широкие структурные изменения и обеспечивают не только адаптивную (приспособительную), но и прогрессивную эволюцию.
Среди перечисленных выше моментов увеличение размеров генома в эволюции эукариот привлекает особое внимание. Этот процесс может осуществляться различными способами. Наиболее резко размер генома изменяется в результате полиплоидизации, которая достаточно широко распространена в природе. Она заключается в увеличении количества ДНК и хромосом, кратном гаплоидному. Достигаемое в результате состояние полиплоидии приводит к увеличению дозы всех генов и создает избыток «сырого» генетического материала, который впоследствии видоизменяется в результате мутаций и отбора.
По-видимому, в ходе эволюции в результате накопления мутаций и дивергенции нуклеотидных последовательностей полиплоидизация сопровождалась переходом к диплоидному состоянию. Само по себе увеличение дозы генов еще не означает достижения однозначно положительного биологического результата. Об этом свидетельствует развитие в эволюции эукариот механизмов компенсации возрастающей дозы генов в ходе их экспрессии путем сокращения времени жизни в клетках зрелой РНК. Так, у тетраплоидных карповых рыб в ответ на увеличение дозы генов рРНК в молекулах рРНК соматических клеток возникают скрытые внутренние разрывы, которые приводят к преждевременному их старению и сокращению содержания в цитоплазме.
Если бы увеличение объема генома происходило только в результате полиплоидизации, то в природе должно было бы наблюдаться скачкообразное изменение его размеров. На самом деле этот процесс демонстрирует плавное увеличение содержания ДНК в геноме. Это позволяет допустить возможность других механизмов, изменяющих его объем.
Действительно, некоторое значение в определении размера генома имеют хромосомные перестройки, сопровождающиеся изменением содержания ДНК в них, такие, как дупликации, делеции и транслокации. Они обусловливают повторение, утрату некоторых последовательностей в составе хромосомы или перенос их в другие хромосомы.
Важным механизмом увеличения объема генома является амплификация нуклеотидных последовательностей, которая заключается в образовании их копий, что приводит к возникновению повторяющихся участков ДНК. Особенностью генома эукариот является наличие таких повторов в большом количестве, свидетельствующее о существенном вкладе механизма амплификации в увеличение размеров наследственного материала. Амплифицированные последовательности образуют семейства, в которых они собраны вместе (тандемная организация) или же распределяются по разным хромосомам. Конкретные изменения, приводящие к амплификации, бывают различными. Появление тандемов повторяющихся последовательностей объясняется, например, неравным кроссинговером, вследствие которого возникают многократные дупликации отдельных участков ДНК. Возможна амплификация путем вырезания фрагмента с последующей его репликацией вне хромосомы и встраиванием копий в другие хромосомы. Предполагают также амплификацию, осуществляемую путем «обратной транскрипции» ДНК на РНК с участием фермента обратной транскриптазы с последующим встраиванием копий ДНК в различные локусы хромосом.
Во всех случаях амплификация некоторой последовательности приводит к возникновению в геноме более или менее многочисленных повторов и способствует некратному увеличению его объема. Наличие таких повторов в сочетании с мутационным процессом является предпосылкой дивергентной эволюции однотипных последовательностей в пределах семейства с соответствующим изменением свойств кодируемых белков или РНК.
Ярким примером эволюционной судьбы амплифицированных нуклеотидных последовательностей являются семейства глобиновых генов, широко распространенных в природе у видов разных уровней организации. У высших позвоночных известен ряд глобиновых генов, контролирующих синтез полипептидов гемоглобина. У человека в геноме имеется восемь активных глобиновых генов, образующих два семейства.
Семейство генов, определяющих синтез а-глобинов, содержит два –глобиновых гена, активно функционирующих в эмбриогенезе, и два –глобиновых гена, активно функционирующих в эмбриогенезе, и два  -глобиновых гена, которые экспрессируются у плода и взрослого человека. Это семейство генов располагается в 16-й хромосоме в следующем порядке: 5' — -глобиновых гена, которые экспрессируются у плода и взрослого человека. Это семейство генов располагается в 16-й хромосоме в следующем порядке: 5' — 2 —1— 2— 2 —1— 2— 1—3'. Семейство генов, определяющих синтез 1—3'. Семейство генов, определяющих синтез  глобинов, расположенное в 11-й хромосоме, содержит глобинов, расположенное в 11-й хромосоме, содержит  –глобиновый ген эмбриона, два сходных –глобиновый ген эмбриона, два сходных –глобиновых гена плода –глобиновых гена плода  и и  , малый , малый  - и большой - и большой  -глобиновые гены взрослых: 5' — — — — -глобиновые гены взрослых: 5' — — — — — — — 3'. — 3'.
Изучение гомологии продуктов указанных генов и генов миоглобина у разных видов организмов позволило предположить общность происхождения этих семейств. Вероятно, около 1100 млн. лет назад произошла дупликация гена-предшественника, давшая начало гемоглобиновым и миоглобиновым генам. Позднее, около 500 млн. лет назад, на ранней стадии эволюции позвоночных произошла дупликация, давшая начало двум ( и ) семействам глобиновых генов, сопровождавшаяся транслокацией. Примерно 200 млн. лет назад очередная дупликация привела к возникновению в семействе –глобиновых генов генов –глобинов плодов и взрослых. Около 100 млн. лет назад произошло образование –  -глобиновых генов и, наконец, 40 млн. лет назад появились – и –глобиновые гены. -глобиновых генов и, наконец, 40 млн. лет назад появились – и –глобиновые гены.
Семейства – и –глобиновых генов организованы в генные кластеры, возникшие, вероятно, в результате тандемной дупликации генов. В составе указанных кластеров наряду с активно функционирующими на разных стадиях онтогенеза генами обнаружены неактивные, или псевдогены. Последние возникли, вероятно, в результате появления в них изменений, несовместимых с возможностью их экспрессии. В семействе –глобиновых генов содержится два псевдогена: 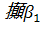 и и 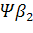 . В -семействе имеется один псевдоген. Дивергенция амплифицированных последовательностей с образованием разных генов или их семейств обусловлена накоплением в них различных изменений в виде замен оснований или других генных мутаций. О гомологии глобиновых генов обоих семейств свидетельствует наличие во всех активных глобиновых генах позвоночных двух интронных участков, занимающих в них строго одинаковое положение. Такую же организацию имеют и псевдогены . В -семействе имеется один псевдоген. Дивергенция амплифицированных последовательностей с образованием разных генов или их семейств обусловлена накоплением в них различных изменений в виде замен оснований или других генных мутаций. О гомологии глобиновых генов обоих семейств свидетельствует наличие во всех активных глобиновых генах позвоночных двух интронных участков, занимающих в них строго одинаковое положение. Такую же организацию имеют и псевдогены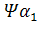 у человека, у человека, 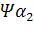 у кролика. Однако в у кролика. Однако в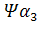 псевдогене мыши в ходе эволюции оба интрона оказались точно вырезанными. псевдогене мыши в ходе эволюции оба интрона оказались точно вырезанными.
Результатом амплификации небольших последовательностей ДНК в пределах функциональной единицы является удлинение гена, при котором из простых генов могут возникать более сложные. Это может происходить за счет тандемных дупликаций. Например, в генах, кодирующих вариабельные участки иммуноглобулинов мыши, последовательности из 600 п.н. образуются в результате 12 тандемных повторов исходной предковой последовательности в 48 п.н. Другим примером удлинения гена посредством тандемных дупликаций служит ген коллагена , который у курицы состоит из 34 000 т.н. и содержит больше 50 экзонов. Длина таких участков во всех случаях кратна девяти нуклеотидным парам. Эволюция этих экзонов, очевидно, шла от гипотетического исходного строительного блока длиной в 54 пары нуклеотидов. , который у курицы состоит из 34 000 т.н. и содержит больше 50 экзонов. Длина таких участков во всех случаях кратна девяти нуклеотидным парам. Эволюция этих экзонов, очевидно, шла от гипотетического исходного строительного блока длиной в 54 пары нуклеотидов.
Таким образом, амплификация нуклеотидных последовательностей, происходившая в процессе эволюции генома, обеспечивала не только его количественное увеличение, появление семейств генов, но и создавала предпосылки для накопления в них изменений, дивергенции генов, увеличения разнообразия контролируемых ими продуктов. [1]
2. Роль и мобильных генетических элементов горизонтального переноса генов в эволюции генома
2.1 Мобильные генетические элементы, их свойства
Мобильные генетические элементы (МГЭ, подвижные элементы, транспозируемые элементы, транспозоны и т.д.) повсеместно распространены в живой природе от плазмид, фагов и бактерий до высших животных и растений. Будучи нестабильными по своей локализации в геномах, они создают мощный источник изменчивости генов, систем их управления и геномов. Будучи сами последовательностями нуклеотидов, они тоже подвержены эволюции. Поэтому МГЭ выступают и как факторы эволюции содержащих их геномов, и как эволюционирущие объекты.
В настоящее время МГЭ найдены у бактерий (включая их фагов и плазмид), низших грибов, насекомых, растений, животных и многих других объектов. Число известных семейств МГЭ, вероятно превышает 100. У хорошо изученных объектов найдены многие десятки семейств МГЭ: например у дрозофилы их число, вероятно, достигает 50. Некоторые семейства относятся к умеренным повторам, имея десятки-сотни копий на геном, другие - к повторам высокой множественности (Alu у человека - 5—6 млн копий на геном). В сумме МГЭ различных семейств могут составлять значительную часть генома (до 10% у дрозофилы). МГЭ нестабильны, т.е. с определенной вероятностью способны к транспозициям и исключению из отдельных позиций генома (табл.1).
Таблица 1. Некоторые свойства МГЭ (Хесин, 1984; Fennegan,1985)
| МГЭ |
Частота транспозиций на позицию, за поколение |
Количество копий МГЭ в геноме |
МГЭ |
Частота транспозиций на позицию, за поколение |
Количество копий МГЭ в геноме |
| FB |
10-3 |
30 |
Te |
10-3 |
10 |
| P |
10-2-10-5 |
30-50 |
MU |
10-1-10-3 |
10-60 |
| Ty |
10-7 |
30-35 |
MDG |
10-2-10-5 |
10-50 |
МГЭ способны к воспроизведению в клетке либо через репликацию ДНК, либо через прямую и обратную транскрипцию (ретротранспозоны).В случаях, когда МГЭ не содержит генов, выполняющих клеточных функции, их часто считают "эгоистическими ДНК". Транспозиции обычно связаны с размножением копий МГЭ. В своей структуре МГЭ содержат гены транспозиции (ферментов-транспозаз - Тп-3, Тп-5 ) и др., ревертаз - ретропозоны (Ананьев, 1989). Поэтому фактически они являются отдельными репликонами. В некоторых случаях синтез транспозазы репрессируется при избыточной ее концентрации по механизму отрицательной обратной связи (Тп-3), Р-фактор дрозофилы). МГЭ содержат также разнообразные функциональные сайты - знаки пунктуации и управления (промоторы, терминаторы, операторы, репликаторы, энхансеры, регуляторные сайты теплового шока), которые существенны для окружающих участков генома. Инсерции МГЭ в кодирующие зоны генов приводят к нарушению или резкому изменению их функций. Это связано с прямым нарушением генов и с влиянием знаков пунктуации (промоторов, терминаторов и др.,) на процессы считывания. Доля таких мутаций особенно велика у прокариот, которые имеют высокую плотность кодирования информации в геноме. Инсерции МГЭ в некодирующие области (спейсеры, интроны, фланговые участки др.) приводят к более "мягким" последствиям: усилению или ослаблению активности близлежащих генов, изменению их регуляции и т.п. Такие последствия преобладают у высших эукариот, у которых кодирующая часть генома составляет ~3-5%. Показано также, что среди видимых мутаций у дрозофилы и других объектов наиболее значительную долю составляют не замены нуклеотидов, а именно инсерции МГЭ (табл. 2).
Таблица 2 Доля спонтанных мутаций, вызванных инсерциями МГЭ, в различных локусах Drosophilla melanogaster (Sankaranarayanan, 1988; McDonald, 1989)
| Локус |
Доля инсерций МГЭ среди видимых спонтанных мутаций |
МГЭ (число мутаций) |
| V (vemillion) |
4/5 |
mg-2-(3), BI04 (1) |
| ct (cut) |
28/28 |
mdg-4 (27), copia (1) |
| ry (rosy) |
3/5 |
calypso (1), BI04 (1) |
| f (forked) |
¾ |
mdg-4 (3) |
| su (s) |
5/7 |
mdg-4 (5) |
| Bx (Beadex) |
4/4 |
mdg-4 (2) ,BI04 (2) |
| bx (bithorax) |
8/9 |
mdg-4 (7) ,BI04 (2) |
| sc (scute) |
2/2 |
mdg-4 (2) |
| Antr (Antennapedia) |
2/5 |
BI04 (2) |
2.2 Отбор, температурные воздействия и динамика рисунка локализации МГЭ в геноме
Гибридизация in situ ДНК МГЭ с политенными хромосомами дрозофилы была тем методом, который позволил исследовать популяционную динамику рисунков локализации МГЭ. Гвоздев, Кайданов и др. впервые обнаружили, что отбор по количественному признаку половой активности самцов дрозофилы приводил к направленному изменению рисунка локализации copia-подобных МГЭ. Сходные явления были обнаружены также в других системах МГЭ дрозофилы.
Васильева и др. (1987) селекционировали лабораторную популяцию D. melanogaster с прерванной радиальной жилкой крыла (олигогенная мутация radius incompletus — ri). Контрольная популяция riC имела проксимальный и дистальный фрагменты (табл. 3).
Таблица 3. Средняя длина проксимального (Х1) и дистального (Х2) фрагментов радиальной жилки крыла у самок различных линий дрозофил (генеалогию линии см. на рис. 1)
| линия |
год начала культивирования линии |
поколение |
Х1 |
Х2 |
| riC |
1974 |
220 |
1.91÷0.05 |
0.39÷0.07 |
| riSN |
1974 |
220 |
0.50÷0.02 |
отсутствует |
| riTII3 |
1979 |
140 |
1.05÷0.05 |
отсутствует |
| riTII49 |
1979 |
140 |
2.45÷0.09 |
1.47÷0.08 |
| riSP |
1982 |
40 |
3.80÷0.02 |
2.20÷0.03 |
В результате 70 поколений минус-отбора жилка была почти полностью редуцирована, и далее фенотип устойчиво воспроизводился без дальнейшего отбора в течение более 300 поколений (линия riSN). В результате 40 поколений плюс-отбора жилка была полностью восстановлена, фенотип далее тоже воспроизводится без дальнейшего отбора многие десятки поколений (линия riSP). В табл. 3 приведены количественные итоги селекции. Сравнительный анализ рисунков локализации МГЭ mdg-1, mdg-2 и copia показал (Васильева и др., 1987; Забанов и др., 1990), что все они имеют в контрольной линии совершенно различные рисунки локализации. В результате отбора по признаку в любом из двух направлений происходили характерные изменения рисунков, хорошо воспроизводимые в повторных экспериментах. Результаты изменения признаков и рисунков локализации МГЭ достаточно устойчиво воспроизводились в отселекционированных линиях. На рис. 1 изображено генеалогическое дерево этих и других исследованных линий.
Куколки дрозофил контрольной популяции (riC) были подвергнуты температурному воздействию: ступенчатому изменению температуры культивирования — 29° -» 18° С. Фенотипы обработанных особей оказались чувствительными к этому фактически стрессовому воздействию. В наибольшей степени это проявилось при обработке в течение двух коротких периодов куколочной стадии. Опуская детали, укажем, что главным эффектом было изменение экспрессии признака в последующих поколениях, которое устойчиво наследуется свыше 200 поколений. Температурная обработка в 1-й чувствительный период (113 ±5 ч.) приводила к существенной редукции количественного признака в последующих поколениях, обработка во 2-ой чувствительный период (149 ± 5 ч.) — к почти полному восстановлению жилки. Соответствующие "температурные" линии были названы riТIIЗ и riТI49 (см. табл. 3 и рис. 1).
Генетический анализ "селекционных" и "температурных" линий показал, что полигенные факторы, влияющие на количественный признак, распределены по всем хромосомам дрозофилы (Васильева и др., 1987; Забанов и др., 1990).
Гибридизация in situ тех же copia-подобных элементов с полигенными хромосомами личинок этих линий выявила вполне определенные, существенные и воспроизводимые изменения рисунков локализации по сравнению с контрольной линией (riC). Таким образом, разовое температурное стрессовое воздействие вызывает множественные транспозиции copia-подобных МГЭ и изменения полигенов, сопровождаемые также изменениями фенотипа. Укажем, что по фенотипу полученные "температурные" линии попарно сходны с "селекционными": riSN с пТПЗ, riSP с riTI49 (см. табл. 3). В ряде изогенных линий дрозофилы множественные перемещения МГЭ обнаружены непосредственно в следующем поколении (Колесникова и др., 1991).
Рисунки локализации МГЭ по сегментам цитологической карты полигенных хромосом представляют собой линейные последовательности бинарных символов. Их можно сравнивать между собой теми же способами, что и последовательности мономеров. В каждой из пяти линий были взяты по 7—9 личинок. Для каждой пары последовательностей строились расстояния по Хеммингу, матрица таких расстояний была использована для построения дерева сходства. На рис. 2 изображено такое бескорневое дерево, построенное методом UPGMA для рисунков mdg-2.
Основной результат такого анализа состоит в том, что линии с наиболее сходными рисунками локализации МГЭ оказались также наиболее близкими и по количественным проявлениям признаков: riSN и пТПЗ, riSP и riTI49. Еще раз подчеркнем, что сходство по признаку и по рисунку локализации МГЭ проявили линии, полученные принципиально разными путями: "селекционные" — путем длительного жесткого отбора по фенотипу, "температурные" — путем разовой температурной обработки в чувствительные периоды развития. Этот факт свидетельствует о том, что рисунок локализации МГЭ и экспрессия признака связаны между собой причинно (генотип — фенотип).
Убедимся, что с точки зрения популяционных механизмов — генетического дрейфа, накопления транспозиций и др.— эта связь не случайна. Во-первых, она воспроизводима при повторном выводе "температурных" линий в 1979,1982,1985,1986 гг. Во-вторых, роль генетического дрейфа в возникновении различий между линиями можно считать малой. При культивировании всех линий выполнялся комплекс антидрейфовых мероприятий, препятствующих случайной фиксации или потере вариантов локализации МГЭ (...) т.е. случайные флуктуации частот нивелировались, а фиксация становилась маловероятной. В-третьих, мы имели для сравнения два независимо построенных дерева — генеалогическое дерево линий (см. рис. 1), отражающее реальный процесс их выведения, и дерево сходства линий (рис. 2), отражающее степень их близости по рисункам локализации МГЭ. Сравнения этих деревьев показывает, что они совершенно различны. Это значит, что в процессе выведения и микроэволюции этих линий были нарушены условия, необходимые для того, чтобы дерево сходства дивергировавших последовательностей отражало реальную топологию дерева эволюции. В-четвертых, имеется ряд факторов, указывающих на "взрывообразный" характер транспозиций МГЭ в "температурных" линиях. По данным Колосниковой и др. (1991), в изогенных линиях, выведенных из контрольной линии пС, в следующем поколении после теплового шока найдены множественные транспозиции mdg-2. Юнакович и др. , использовав блотинг по Саузерну и зонды на присутствие copia-подобных МГЭ, показали, что после теплового шока в следующем поколении происходили массовые перемещения МГЭ. С другой стороны, и в "селекционных" линиях отбор происходил весьма неравномерно и занял лишь часть времени микроэволюции, а затем был прекращен, т.е. и здесь нельзя говорить о постоянстве скорости фиксации транспозиций (Васильева и др., 1987). Именно эти особенности являются причиной несовпадения двух построенных деревьев (рис. 1, 2). Заметим, что этот случай является одним из наиболее наглядных примеров того, что дерево сходства последовательностей не всегда изоморфно отображает реальный процесс эволюции.
Наконец, в-пятых, отметим, что сходство линий по рисункам локализации МГЭ может быть либо "остаточным", поскольку все линии имеют общее происхождение (см. рис. 1), либо вновь приобретенным, если спектры изменений у линий сходны, неслучайны. Сравнение спектров отличий четырех дочерних линий от контрольной (riC) показывает, что наиболее сходные по фенотипу линии имеют наиболее сходные спектры изменений локализации mdg-2. Так, линии riSN и riT 113 имеют 27 общих изменений из 33—35 по сравнению с riC; линии riSP и riT 149 имеют 17 общих изменений из 23—30 замен. Следовательно, подавляющая часть спектров изменений в высокой степени не случайна по своему положению канализована.
Многие другие copia-подобньге МГЭ обладают подобными же свойствами. Рисунки локализации mdg-1, mdg-3, copia почти полностью различны, но деревья сходства рисунков похожи друг на друга. В то же время, рисунки МГЭ вовлекаются в отбор по многим различным количественным признакам. Поэтому можно полагать, что мы имеем дело с геномной системой влияния МГЭ на проявление различных генов, чувствительной к стрессовым внешним воздействиям и отвечающей на селекцию. [4]
2.3 Гипотеза: эволюционная роль системы МГЭ в геномах эукариот
Суммируя вышеизложенное, можно констатировать, что система МГЭ эукариотического генома обладает, по крайней мере, следующими общими функциями: 1) является источником инсерционной изменчивости генов; 2) влияет на проявление количественных и качественных признаков; 3) откликается изменением рисунков локализации многих МГЭ на отбор по признакам; 4) откликается на внешние стрессорные воздействия, в частности — температурные, вспышками транспозиционной изменчивости. Такими свойствами в разной степени обладают МГЭ различных объектов — дрожжей, дрозофилы, растений, млекопитающих.
Механизм инсерционной изменчивости достаточно ясен. Отметим только, что активация транспозиций разными путями (дисгенное скрещивание а Р—М системе (МсКау, 1988), температурное воздействие в системе copia-подобных МГЭ должна усиливать эти компоненты изменчивости. Укажем также на существование "транспозиционных взрывов" в отдельных генеративных клетках дрозофилы, возникающих после генетического воздействия или спонтанно.
Механизм влияния МГЭ на экспрессию генов эукариот требует пояснений. В целом МГЭ содержат широкое разнообразие регуляторных сайтов, среди которых наиболее интересны энхансеры и регуляторные сайты теплового шока. Известно, что энхансеры способны в десятки и сотни раз усиливать транскрипцию соседних генов на расстоянии до 5000 н.п. методом контекстного анализа показали, что в структуре многих МГЭ (в том числе — mdg-1, mdg-2, mdg-4, copia) имеются энхансеро-подобные сайты.
Регуляторные сайты теплового шока (РСТШ) присутствуют в 5'-областях перед началом транскрипции эукариотичкеских генов, проявляющих эффект теплового шока. Размер их ~14 н.п., расстояние до блока Хогнесса sS 200 н.п. Восстановлен консенсус РСТШ, характерный для таких далеких форм, как дрожжи, дрозофила, лягушка, человек: CNNGAANNTTCNNG (N—любой нуклеотид). Здесь 8 консервативных позиций. В некоторых случаях РСТШ имеют другую структуру. Чем ближе РСТШ к блоку Хогнесса, тем эффективнее активация транскрипции тепловым шоком (Bienc, 1985). РСТШ можно рассматривать как энхансеры с позитивной регуляцией.
Показано, что активация транскрипции МГЭ copia у дрозофилы и DIRS1 у дрожжей при тепловом шоке связана с наличием РСТШ в их терминальных повторах). Капитонов и др. (1987) методом контекстного анализа обнаружили РСТШ-подобные сайты в семи из 13 исследованных ими МГЭ: mdg-1, mdg-4, hobo, P дрозофилы, BS1 и CIN1 кукурузы и в ретровирусе EV1 курицы. Все они находились на расстоянии ^ 150 н.п. от начала транскрипции и имели неслучайную гомологию либо с известными РСТШ, либо с их консенсусом. В mdg-2 и в МГЭ H.M.S. Beagle дрозофилы этот вариант РСТШ не обнаружен, но это не исключает возможного присутствия других РСТШ. Сходные результаты независимо получены также Макдональдом и сотрудниками. Интересно, что РСТШ найден также в последовательности секвенированного ретровируса HTLV—III (вирус СПИД). В некоторых МГЭ найдены РТСШ-подобные сайты на расстоянии > 200 н.п. от блока Хогнеса. Отметим, однако, что наличие РТСШ-подобного сайта еще не гарантирует его функционирование.
Прямое доказательство индукции транспозиций copia-подобного МГЭ mdg-2 при помощи теплового шока получено Колесниковой и др. (1991). В изогенной линии, самцы которой были подвергнуты тепловому шоку, транспозиции были найдены в следующем поколении, причем вероятности транспозиций возросли более чем на два порядка величин.
Таким образом, появление дополнительного энхансера вблизи функционального гена в результате инсерции несущего его МГЭ способно резко активизировать этот ген. Инсерция МГЭ, содержащего РСТШ, способна подчинить соседние гены системе теплового шока и сделать их чувствительными к стрессорным факторам окружения. В связи с этим можно сформулировать гипотезу о роли МГЭ в ответе на отбор по признакам и в температурных эффектах.
На основе имеющихся фактов (см. выше) необходимо постулировать, что рисунки локализации МГЭ являются существенной компонентой генетического механизма детерминации количественных признаков (Vasilyeva et al., 1985; Ratner, Vasilyeva, 1989). В общем случае эти системы содержат: 1) олигогены (гены главного эффекта), необходимые для формирования признака; 2) полигены, каждый из которых не необходим для формирования признака, но в совокупности они могут существенно изменить его экспрессию; 3) МГЭ, которые модифицируют, усиливают действие олигогенов и полигенов, вблизи которых они локализованы. Рисунок локализации МГЭ в каждом случае относительно стабилен; позиции, доступные для инсерции, вероятно, мечены на длительный срок "молекулярной памятью", существование которой в частном случае показано Мизрохи и др. (1985); возможно, что список доступных позиций зависит от компактизации хромосом и других явлений ядерно-хромосомного уровня. Рисунок локализации МГЭ распределяется по этим специфическим позициям (см. параметры n и m в п. 2). Скорее всего, рисунок МГЭ более или менее случайно наложен на топографию локализации олигогенов и полигенов и активирует их.
В рамках концепции лимитирующих факторов необходимо также предположить, что влияние как полигенов, так и МГЭ проявляется фенотипически только в тех случаях если они действуют на лимитирующие олигогены. Тогда их эффект оценивается отбором: вариабельность рисунков будет создавать вариабельность лимитирующих признаков, отбор по признакам будет приводить к отбору полигенов и рисунков модифицирующих их МГЭ. Определенный вклад в изменчивость признаков будут вносить также новые редкие транспозиции МГЭ. Ясно, однако, что селективное изменение рисунков не будет равномерным, поскольку это не нейтральный процесс фиксации перемещений МГЭ.
Для объяснения температурных эффектов предполагается, что стрессовое температурное воздействие (тепловой шок, ступенчатое изменение температуры и др.) индуцирует вспышку множественных у отдельных особей и массовых в популяции транспозиций МГЭ, которые выявляются уже в следующем поколении (Колесникова и др., 1991) и приводят к наследуемому изменению количественных и качественных признаков. Неслучайный характер температурно-индуцированных транспозиций можно связать с существованием каких-то достаточно жестких ограничений на локализацию этих транспозиций в геноме. Имеется в виду, что в рамках группы выделенных сайтов, в принципе доступных для инсерций МГЭ, т.е. обладающих гомологией к ним, имеются дополнительные ограничения на транспозиции, связанные со свойствами хромосом или клеточного ядра и изменяющиеся в ходе онтогенеза. Эти ограничения должны быть различны на разных этапах онтогенеза (в чувствительные периоды). Сходство фенотипов "селекционных" и "температурных" линий сопровождается сходством спектров перемещения МГЭ. Это возможно только в том случае, если спонтанные неиндуцированные транспозиции в "селекционных" линиях подчиняются таким же ограничениям, что и индуцированные в "температурных" линиях.
Последствия температурных воздействий могут быть весьма разнообразны. Непосредственная активация системы генов, подчиненных РСТШ, по-видимому, должна привести к усилению транскрипции и синтеза ферментов транспозиции (транспозаз, ревертаз и др.) и к увеличению вероятности транспозиции. В свою очередь, это вызовет вспышку инсерционного мутагенеза. Наличие энхансеров в структуре МГЭ приводит к заметной активации олигогенов и полигенов, в окрестность которых совершаются транспозиции. Иначе говоря, возможно массовое и множественное изменение экспрессии различных генетических систем. Кроме того, повышение синтеза ревертазы может усилить процесс амплификации различных генов через прямую и обратную транскрипцию.
Следует подчеркнуть, что система теплового шока индуцируется не только повышением температуры, но и воздействиями других весьма разнообразных внешних факторов: вирусным заражением клеток, обработкой ядами, детергентами, другими химическими факторами, нарушением энергетического обмена клеток и т.д. Все эти воздействия являются стрессовыми, неблагоприятными, а реакция системы теплового шока — генерализованной. Кроме того, уровень транскрипции и транспозиций некоторых МГЭ индуцируются УФ- и гамма-облучением, а в Р—М и I—R-системах дрозофилы — дисгенным скрещиванием.
Геномы эукариот, содержащие до 10% МГЭ различных семейств, теперь можно рассматривать как систему разнообразных паттернов МГЭ, которая может быстро перестраиваться под влиянием стрессовых внешних и геномных воздействий. В разных случаях транспозиции могут быть как случайными, так и жестко канализованными по своим локализациям, но они порождают новый вариант изменчивости для полигенных систем. В этом смысле МГЭ можно рассматривать как своеобразные рецепторы стрессирующих сигналов из внешней или генетической среды, "запускающие" системные вспышки наследуемой изменчивости в критические периоды эволюции популяций. Это влечет за собой множественные генетические последствия: быстрое изменение видовой нормы лимитирующих признаков, возможно, изменение лимитирования, изменение спектра дальнейших мутаций и перестроек, становление нового генетического гомеостаза и др. Участие в этих событиях ретропозонов (mdg и др.) означает, что их собственный генетический материал при репликации проходит РНК-стадию, которая имеет вероятность мутирования 10-3—10-4, что на несколько порядков выше, чем в ДНК-содержащих геномах эукариот. Таким образом, гены и функциональные сайты самих ретропозонов подвержены особо сильному мутагенезу. Этоа касается также генов, захваченных ретропозонами из генома хозяина. Критические, стрессовые условия существования часто сопряжены с прохождением популяций через стадию "бутылочного горлышка", которое может быть связано либо с массовым вымиранием; либо с освоением новых экологических ниш по "принципу основателя". В этих условиях новые формы, индуцированные через вспышки транспозиций, могут стать основателями новых популяций с резко измененным фенотипом по лимитирующим количественным или качественным признакам. Здесь возможны как адаптивные, так и случайные варианты быстрого преобразования. В результате фактически эти события могут стать одной из главных компонент изменчивости и эволюции генома дрозофил и других объектов. Не исключено, что изменение системы рисунков локализации МГЭ является одним из механизмов видообразования. Во всяком случае, гибридный дисгенез, индуцирующий транспозиции Р-фактора, является изолирующим механизмом между скрещиваемыми линиями дрозофил.
2.4 Макроэволюция Aiu- подобных повторов
Семейство Alu-повторов млекопитающих является одним из самых больших. У человека оно содержит (5-9) • 105 копий, у шимпанзе в 3 раза больше. Alu-повторы имеют размер ~300 н.п. и содержат два гомологичных мономера -130 н.п., причем второй из них имеет также инсерцию фрагмента 31 н.п. Повторы BI грызунов гомологичны мономерам Alu-повторов (Колчанов и др. 1988) и имеют -150 н.п., а также вставку 32 н.п. Предполагается, что Alu и BI транспозируются в геноме через механизм обратной транскрипции, т.е. являются ретропозонами, хотя строго это не доказано. В настоящее время секвенировано свыше 100 Alu-повторов приматов и десятки BI-повторов грызунов. Другую родственную группу повторов, имеющую распространение от архебактерий до человека, представляют собой гены 7SL РНК. Эта фракция РНК входит в состав рибонуклеопротеидных частиц, участвующих в транспорте секретируемых белков эндоплазматической сети. Колчанов и др. (1988) выполнили совместный генетический анализ мономеров Alu-повторов человека, BI-повторов крысы и 7SL РНК лягушки, крысы и человека (рис. 3). Были использованы известные палеонтологические датировки времени существования общих предков приматов и грызунов (~80•106лeт назад), млекопитающих и земноводных (~350•106 лет назад). Другие датировки были определены по дереву.
Основные результаты анализа состоят в следующем. Общим предшественником современных генов 7SL РНК, Alu- и BI-повторов были предки генов 7SL РНК. Однако повторы Alu и BI произошли непосредственно не от генов 7SL РНК приматов и грызунов, соответственно, а от некоторого общего гена-предка, который дуплицировался и дивергировал от генов 7SL РНК еще задолго до выделения филетических линий отрядов приматов и грызунов. Время этой дивергенции оценено ~ 260-106 лет назад, т.е. после разделения земноводных и млекопитающих. При этом предковый ген Alu и BI утратил существенную функциональную часть, которая наиболее консервативна у 7SL РНК. Небольшие инсерции (31 н.п. у Alu и 32 н.п. в BI) были; вероятно, приобретены после разделения грызунов и приматов.Оценка скоростей фиксации замен показала, что для генов 7SL РНК v = 2,2•10-10 замен на позицию в год, а для Alu и BI на порядок больше: v = 2,3•10-9 замен на позицию в год. Это согласуется с представлением об уменьшении функциональной нагрузки Alu и BI-повторов по сравнению с 7SL РНК за счет делеции консервативного функционального фрагмента. Величина скорости фиксации для Alu и BI близка к максимальной скорости нейтральной эволюции Vmax=5*10-9 для ДНК-геномов, что говорит о том, что существенная часть фиксированных замен могла быть нейтральной. Однако многие секвенированные Alu-повторы содержат различные функциональные сайты —промоторы РНК-полимеразы 111, энхансеры, сайты "затравки" обратной транскрипции и др., которые необходимы для транспозиции. Поэтому вряд ли эволюция Alu и BI полностью нейтральна. Alu-повторы человека распределены в геноме неравномерно, образуя кластеры. Шахмурадов и др. исследовали закономерности сходства повторов внутри и между кластерами. Оказалось, что имеется неслучайная гомология повторов и участков их встраивания, т.е. имеется своеобразное "притяжение" копий Alu к определенным зонам генома. Однако кластеры Alu-повторов неоднородны. Копии, входящие в один кластер, не всегда более сходны друг с другом, чем с копиями из других кластеров. Следовательно, рассеяние копий в геноме идет не по локальному принципу ,а скорее через общий пул ретропозонов от различных кластеров. Таким образом, филогенетический анализ последовательностей Alu-подобных МГЭ позволил высказать ряд важных соображений об их происхождении, дивергенции и характере их эволюции.
3. Роль горизонтального переноса генетического материала в эволюции генома
Наряду с транспозонами, не способными очевидно, существовать вне генома и образовывать свободные молекулы ДНК, описаны элементы, обнаруживаемые как в составе генома, так и вне его. Существование таких подвижных элементов дает возможность обсуждать роль горизонтального переноса генетического материала в эволюции генома.
Если описанные выше изменения структуры генома передаются из поколения в поколение организмов одного и того же вида, т.е. по вертикали, то горизонтальный перенос генетической информации может происходить и между организмами разных видов, одновременно существующими на Земле.
Горизонтальный перенос генов является главным источником инноваций, инструментом быстрого приобретения и возникновения новых генов, способных радикально изменить свойства клеток, расширить их адаптационный потенциал. Изменчивость организмов в результате горизонтальной передачи генов реализуется через различные каналы генетической коммуникации - процессы коньюгации, трансдукции, трансформации, процессы переноса генов в составе векторов - плазмид, вирусов, мобильных элементов. Активный перенос генов может происходить в симбиотических, паразитарных или ассоциативных системах, где осуществляется физический контакт клеток. В сущности, современная генетическая инженерия, использующая разного типа векторы, базируется на принципах горизонтального переноса генов, хотя еще недавно не было четкого понимания того, что такого рода генная инженерия широко распространена в природе и играет важную роль в эволюции. И только работы в области геномики в последние 10 лет доказали, что горизонтальный перенос генов был и остается (особенно в мире прокариот) одним из главных механизмов видообразования. Конечно, в ходе вертикальной эволюции повышалась степень автономизации организмов, возникали и совершенствовались барьеры, препятствующие горизонтальным генным переносам и "размыванию"
геномов. Это касалось и ограничения контактов между организмами, механизмов проникновения и транспортировки молекул ДНК, действия систем рестрикции, разрушающих "чужую" ДНК, репаративных механизмов, обеспечивающих стабильность собственных геномов. Поэтому частота горизонтальных переносов была наиболее высокой на ранних этапах становления биосферы и снижалась по мере эволюции высших эукариот с усложнением организации генетического аппарата и развитием систем репродуктивной изоляции.
Горизонтальный перенос генов можно выявить по ряду показателей. Во-первых, по нуклеотидному составу ДНК (ГЦ-содержание), который является видоспецифичным признаком. Отличие в нуклеотидном составе отдельного сегмента от остальной части генома является указанием на присутствие "чужих" генов; иногда это целые кластеры, содержащие профаги, мобильные элементы, "островки" патогенности и т.д. Во-вторых, по частоте встречаемости в гене определенных кодонов. В генах каждого вида преимущественно используется ограниченный набор кодонов. Например, в гене gap у кишечной палочки содержится 20 лейциновых кодонов, но из 6 синонимичных триплетов кодон CTG встречается 19 раз, а кодон TTA только один раз. В гомологичном гене Bacillus subtilis кодон CTG вообще не используется, а преимущественно встречается TTA кодон. Таким образом, "чужие" гены легко обнаружить по отличию в частоте встречаемости кодонов. При этом, однако, надо учитывать, что в ходе длительной эволюции происходит амелиорация нуклеотидного состава, т.е. процесс унификации использования кодонов за счет мутаций и рекомбинации. "Чужие" гены становятся неотличимыми от своих собственных. Третий важный критерий - существенное отличие в положении анализируемого гена на филогенетическом дереве от большинства других генов. О "чужеродном" происхождении гена может говорить и высокая степень его сходства с гомологичным геном из отдаленного таксона при отсутствии подобного гена у филогенетически близких "родственников". Например, в случае, когда типично эукариотический ген вдруг обнаруживается у какого-то одного вида бактерий, а у других бактерий этого гена нет. На основе применения рассмотренных выше критериев можно достаточно легко выявить "вкрапления" в геном чужих сегментов ДНК, приобретенных в результате горизонтального переноса. Возможно три варианта переносов: 1) Приобретение нового гена, для которого нет гомолога в собственном
геноме и в геномах филогенетически родственных организмов. В этом случае возникает принципиально новое качество; 2) Приобретение паралогичного (структурно похожего) гена с генетически отдаленным родством. В результате такого переноса увеличивается функциональное разнообразие белков в клетке; 3) Приобретение нового гена ксенолога, функционально замещающего свой собственный ген, который при этом, как правило, элиминируется. Новый и старый гены структурно различаются между собой, но обеспечивают аналогичные физиологические функции.
Какую выгоду может получить организм, приобретая чужой ген путем горизонтального переноса? 1) Новый путь биосинтеза или катаболизма, обеспечивающий организму преимущества в изменившихся условиях; например, появление способности утилизировать новый субстрат. 2) Повышение устойчивости к антибиотикам, токсинам, патогенам, подавляющим рост клеток данного вида; через горизонтальный перенос могут быть получены и гены, ответственные за средства "нападения", характерные, например, для патогенных микроорганизмов. 3) Замещение предсуществующих генов такими генами, продукты которых увеличивают эффективность функционирования клеточных систем: например, повышение термоустойчивости, резистентности к ингибиторам, оптимизация кинетических характеристик белка, интеграция в сложные комплексы и т.п. 4) Приобретенные гены могут оказаться и функционально нейтральными, дублирующими уже имеющиеся гены; такие дополнительные гены являются страховкой для организма в тех случаях, когда свой собственный ген будет поврежден мутацией или "замолчит" из-за нарушения в системах регуляции. Приобретение "чужих" генов может изменить направление эволюции вида, существенно повлиять на фенотип организма, на его способность к адаптации в экологическом сообществе. Новый ген может дать начало новой субпопуляции, которая способна вытеснить предсуществующий вид. Горизонтальный перенос генов способствует ускорению эволюционного процесса, по сравнению с градуальным накоплением мутаций или внутригеномными перестройками. Конечно, при этом не отрицается селективное значение мутационных утрат какой-то функции и важная эволюционная роль мутаций в генах, контролирующих стабильность генома (системы репликации, репарации, модификации ДНК и т.д.) и механизмы регуляции и координации генного действия. В таблице 1 приведены некоторые данные о горизонтальном переносе генов у прокариот, полученные при полногеномном анализе большого числа видов бактерий и архей. Эти сведения позволяют сделать ряд обобщений, касающихся закономерностей латерального генного переноса.
Табл. 1 Горизонтальный перенос генов у архей и бактерий
| Вид |
число генов в геноме |
перенесенные гены |
| количество |
% в геноме |
| АРХЕИArchaeoglobus fulgidus Methanococcus jannaschii Pyrococcus horikoshii Aeropyrum pernix |
2407 1715 2064 2694 |
179 77 154 370 |
8.4 5.0 7.6 14.0 |
| ПАТОГЕННЫЕ БАКТЕРИИ Mycoplasma pneumoniae Chlamydia trachomatis Rickettsia prowazekii Treponema pallidum Haemophilus influenzae Helicobacter pylori Mycobacterium tuberculosis |
677 894 834 1031 1709 1553 3918 |
39 36 28 77 96 89 187 |
5.9 4.3 3.6 8.3 6.2 6.4 5.0 |
| СВОБОДНОЖИВУЩИЕ БАКТЕРИИ Aquifex aeolicus Thermotoga maritima Escherichia coli Pseudomonas aeruginosa Bacillus subtilis Synechocystis sp. |
1552 1846 4289 4036 4110 3169 |
72 198 381 411 537 219* |
4.8 11.6 9.6 10.1 14.8 7.5 |
| Из обзора: Koonin E.V., Makarova K.S., Arvind L. 2001. Annual Rev. Microbiol. v.55: 709-42 |
1) Доля латерально полученных генов варьирует у разных видов и может достигать 10-15% от общего числа генов в геноме . 2) Наибольшее количество переносов характерно для свободноживущих бактерий с широкими экологическими ареалами (почвенные бациллы, псевдомонады и др.). 3) Наименьшее число переносов обнаружено у большинства патогенных бактерий, живущих в узких эконишах; эти переносы, однако, весьма важны, так как определяют такие признаки как вирулентность и токсичность. 4) Переносы специфичны, поскольку приобретенный ген обнаруживается, как правило, только в клетках определенного вида или даже штамма. 5) Реже всего в горизонтальные переносы вовлечены гены информационных систем (транскрипции, трансляции, репликации), составляющие базовый геном. Продукты этих генов входят в сложные белковые комплексы, где "чужие" белки не встраиваются или не функционируют. Исключением являются гены, кодирующие аминоацил-тРНК-синтетазы, что связано с особенностями работы изоакцепторных транспортных РНК. 6) Чаще всего в горизонтальном переносе участвуют гены операционных систем, обслуживающих метаболизм, транспортные пути, механизмы сигнальной трансдукции. У многих бактерий среди латерально привнесенных генов большую долю составляют функционально неизученные гены и гены, не имеющие сходных ортологов в реципиентном организме. 7) В составе приобретенных сегментов ДНК часто обнаруживаются профаги, плазмиды, кассеты резистентности, гены белков, участвующих в процессах сайт-специфической и "незаконной" рекомбинации, обеспечивающей интеграцию "чужих" генов. В большинстве случаев трудно определить, какие конкретно организмы могли быть донорами в горизонтальном переносе, поскольку сравнительная геномика дает информацию, в основном, о степени сходства гомологичных генов в разных геномах. Только в случае филогенетически близкородственных видов или штаммов (например, для двух штаммов с разной степенью вирулентности) можно предположительно судить о приобретении или утрате определенного участка генома. В переносах же между представителями разных царств невозможно определить природу первичного донора и реципиента даже в случаях предполагаемого переноса признака термофилии от архей в клетки бактерий. Косвенно о направлении такого переноса можно судить только тогда, когда у многих видов архей какие-то гены встречаются часто, но обнаруживаются в
геноме только одного или весьма ограниченного числа видов бактерий. В частности, на этом основании говорят о возможном переносе генов из термофильных архей в клетки предшественников современных термофильных видов бактерий Thermotoga maritima и Aquifex aeolicus. У этих бактерий около 15%
генома составляют типично архейные гены, которых нет у других бактерий. Это пример адаптивного переноса генов, хотя неясно обеспечил ли он этим бактериям завоевание новой экологической ниши, или перенос произошел вследствие контактов клеток бактерий и архей в общей эконише. По критериям гомологии на геномном уровне горизонтальные переносы генов легко выявляются не только у прокариот, но и у эукариот, в ядерном геноме которых немало генов бактериального или архейного происхождения. Это результат перемещения в ядро генов из митохондрий или хлоропластов, возникших на базе прокариотических геномов, претерпевших редукционную
эволюцию. Обсуждается и другая причина мозаичного строения ядерных
геномов эукариот - за счет интеграции чужеродных фрагментов ДНК, поступающих с пищей, например, при поедании бактерий протистами. Геномный анализ показал, что горизонтальный перенос генов может происходить и в обратном направлении - от эукариот в клетки прокариот. Это возможно в условиях тесных физических контактов в симбиотических или паразитических системах, при наличии челночных векторных переносчиков. Например, предполагается, что донором эукариотического гена термоустойчивой аминоацил-ТРНК-синтетазы у археи Pyrococcus могли быть полихеты. В
геноме патогенных бактерий риккетсии и хламидии обнаружено около 20 эукариотических генов. Среди них несвойственные прокариотам гены белков, транспортирующих АТФ и АДФ. Эти транспортные системы позволяют патогенным бактериям "выкачивать" энергию из клетки хозяина, не затрачивая собственные ресурсы. Другие эукариотические гены, приобретенные патогенными бактериями путем горизонтального переноса, обеспечивают "нападение" на клетку хозяина - это ферменты протеолиза, ингибиторы иммунного ответа, рецепторные белки, облегчающие патогенам проникновение в клетку хозяина. Подобного рода горизонтальные переносы генов, по-видимому, происходили сравнительно недавно в процессе превращения непатогенных штаммов в патогенные. Характерно, что у риккетсии (возбудителя тифа) немало генов гомологичных генам животных, а в геноме хламидии (возбудителя трахомы) больше генов, типичных для растений . Это указывает, во-первых, на различия в путях горизонтального переноса у этих филогенетически далеких друг от друга бактерий, а во-вторых, позволяет предполагать, что предки хламидии были сперва паразитами растений, а затем уже переключились на животных. Не исключено, что переносы генов патогенности шли между риккетсиями и хламидиями, живущими в одной экологической нише в организме млекопитающих. Поскольку гены являются сложными структурами и содержат различные домены, ответственные за разные функции в белковом продукте, то, очевидно, что через горизонтальный перенос могут передаваться не только целые гены или блоки генов, но и фрагменты генов, содержащие отдельные домены. Гены как дискретные единицы могут элиминироваться, но их домены в результате рекомбинационных событий становятся сегментами других генов или участвуют в образовании новых генов. Таким образом, в геномном анализе важно определять не только степень гомологии целых генов, но и наличие доменов в необычных сочетаниях. Встраивание нового домена может менять локализацию белкового продукта в клетке, узнавание и передачу сигналов, и многие другие характеристики, влияющие на физиологическую функцию гена. Выявлено более 40 случаев появления "чужих" доменов, приобретенных сравнительно недавно и еще не подвергшихся амелиорации в составе генов, кодирующих различные белки. В частности, это домены, ответственные за белок-белковые и ДНК-белковые взаимодействия, т.е. участвующие в сборке сложных комплексов или в регуляции действия генов. Как правило, происходит рекомбинационное слияние домена из латерально привнесенного гена с резидентным геном, причем партнерами могут быть и филогенетически отдаленные организмы. В сущности, это то, что соответствует задачам генной и белковой инженерии по искусственному созданию белков с новыми свойствами. Поэтому так важно выяснить механизмы перекомбинации доменов, лежащие в основе возникновения новых генов (и новых видов организмов) в результате горизонтального переноса генов. Суммируя представления о роли горизонтального переноса генов в биологической эволюции можно сделать следующее заключение: Во-первых, геномика не дает информации о том, какие конкретные виды организмов были первичными донорами генов. Обнаруживаемые в
геномах чужеродные гены могли попасть туда через цепочку промежуточных хозяев. Согласно схеме Ф.Дулитла, автора термина "Латеральная геномика", на самых ранних этапах эволюции существовало общее генное "коммунальное хозяйство" в неустоявшихся предковых клетках, между которыми происходил активный горизонтальный обмен генами. В этот период шло накопление генетической информации с последующей автономизацией клеток, давших начало отдельным таксономическим линиям. Картина эволюционных связей в мире предковых прокариот представляла собой не столько ветвящееся дерево, сколько своего рода мицелий с переплетенной сетью горизонтальных переносов в самых разнообразных и неожиданных направлениях, диктуемых возможностями физических контактов клеток в общих или перекрывающихся экологических нишах. Поэтому и геномы прокариот и эукариот мозаичны. Данные геномики позволяют утверждать, что в ходе эволюции происходили массивные генные переносы как внутри царств, так и между ними.
Многие организмы (в первую очередь прокариоты) участвовали в горизонтальных переносах как "проточные" емкости. Если бы в процессе эволюции энтеробактерия Escherichia coli только приобретала, но не теряла гены, то размер ее
генома должен был бы удвоится за 100 млн. лет с тех пор как произошла дивергенция с близкородственной бактерией Salmonella.
Этот расчет сделан на основе применения метода "молекулярных часов". Но размер
генома E.coli почти не изменился за это длительное время: геномный анализ выявил не только приобретение большого числа генов, но и отсутствие многих "старых" генов, обнаруживаемых у сальмонеллы. "Захваченные" гены могут сохраниться в геноме, но могут и подвергнуться элиминации или переносу в клетки другого организма. Так как в ходе эволюции происходит амелиорация чужих генов, то по изменению нуклеотидного состава и частоты встречаемости кодонов можно выявить те гены, которые пришли давно, а какие появились недавно. Баланс генных потоков, приводящих к приобретению и утрате генов, определяет не только адаптивную изменчивость организмов, но и поддержание оптимизированного размера генома . Скорости генных потоков у разных групп организмов различны и влияют на темпы
эволюции
геномов. Геномный анализ позволяет сопоставить время приобретения и утраты определенных генов с геологическими эпохами, экологическими кризисами и региональной динамикой биоты (по крайней мере, в отношении
геномов
модельных видов прокариот и протистов). Другое обобщение заключается в том, что не только сам геном, но и механизмы горизонтального переноса генов являются объектами эволюционного процесса. Эволюционируют каналы генетической коммуникации, механизмы рекомбинации, участвующие в "приживлении" или "отторжении" чужих генов. Методы геномики помогают исследовать закономерности эволюции механизмов рекомбинации, их роль в изменчивости организмов. Если в какие-то периоды глобальных геосферных и биосферных перемен меняется диапазон векторных систем (вирусов, плазмид, мобильных элементов и т.п.), повышается рекомбинационная активность и частота горизонтальных переносов генов, то увеличивается и вероятность приобретения новых генов, обеспечивающих селективные преимущества клеток. Это не менее важный фактор эволюции, чем изменение темпов спонтанного или индуцированного мутагенеза. Волны горизонтального переноса генов могут быть инициированы повышением плотности контактирующих популяций. Там, где нет активных контактов, не будет и эффективного переноса генов. Особенно результативны такие процессы в многокомпонентных сообществах, где взаимодействуют различные организмы, метаболически связанные между собой. [3]
Заключение
В историческом плане вопрос об эволюции генов является важнейшим, поскольку эволюция генов связана с истоками жизни вообще и ее совершенствованием в частности. Поскольку выявлена «начальная роль в происхождении жизни РНК, то предполагают, то начало эволюции генов датируется 3,5-4 млрд лет назад, когда. Сформировались первые молекулы РНК, которые каким-то образом детерминировали синтез белков, т. е. были первыми хранителями генетической информации. Однако когда выявилась необходимость в повышении эффективности синтеза белков, способность кодирования генетической информации перешла к ДНК, которая стала главным хранителем генетической информации. Что касается РНК, то она оказалась между ДНК и белком, став «переносчиком» информации. Конечно, эта гипотеза не имеет доказательств. Тем не менее многие считают, что появление ДНК связано с усложнением структуры клеток и, следовательно, необходимостью солирования большого количества информации по сравнению с ДНК. Другими словами, с началом участия ДНК в хранении генетической информации стал развиваться генетический код. [2]
Суммируя вышеизложенное, можно заключить,
1. Системы МГЭ эукариотических геномов являются источником инсерционной изменчивости, влияют на экспрессию количественных и качественных признаков, изменяют свои рисунки локализации МГЭ в ответ на отбор по признакам и на внешнее стрессорное (температурное и др.) воздействие. Температурные эффекты МГЭ, вероятно, связаны с действием системы теплового шока. МГЭ выполняют роль своего рода рецепторов внешних стрессирующих сигналов, инициирующих вспышки транспозиционной изменчивости в критические периоды эволюции популяций, что может приводить к быстрому преобразованию гомеостатической видовой нормы и, возможно, — к видообразованию.
2. В рамках сальтационной теории неравномерной по темпам эволюции анализ механизмов и закономерностей горизонтального переноса генов дает новый ключ к изучению корреляции геосферных изменений и биологической эволюции на планете.
Список использованной литературы
1. Биология. В 2 кн. Кн.1: Учеб. Для медиц. Спец. Вузов / В.Н. Ярыгин, В.И. Васильева, И. Н. Волков, В.В. Синельщикова; Под ред. В.Н. Ярыгина. – 4-е изд., испр. И доп. – М.: Высш.шк., 2001 – 432 с.: ил. (с. 149-153)
2. Пехов А. П. Биология с основами экологии: Учебник. 2-е изд., испр. И доп. –СПб.: Издательство «Лань», 2004. – 688с.: ил. - ( Учебники для вузов. Специальная литература).(с.315)
3. С.В.Шестаков «Роль горизонтального переноса генов в эволюции», Доклад, прочитанный на теоретическом семинаре геологов и биологов "Происхождение живых систем". 15-20 августа 2003 г., Горный Алтай, стационар "Денисова Пещера". Электронная публикация
4. "Современные проблемы теории Эволюции" (ред. Л.П.Татаринов). М.: Наука, 1993
|