СОДЕРЖАНИЕ
ВВЕДЕНИЕ. 2
РАЗДЕЛ 1. СВИДЕТЕЛЬСТВА ЭВОЛЮЦИИ НАСЕКОМЫХ.. 3
1.1 Окаменелости. 3
1.2 Филогенетические отношения. 5
1.3 Географическое распространение. 5
1.4 Данные палеогеографии. 6
РАЗДЕЛ 2. ВЫМЕРШИЕ ОТРЯДЫ НАСЕКОМЫХ.. 7
2.1 Подкласс Apterygota-первичнобескрылые. 7
2.2 Подкласс Pterygota - крылатые насекомые. 7
2.2.1 Инфракласс Paleoptera – древнекрылые. 7
2.2.2 Инфракласс Neoptera – Новокрылые. 11
РАЗДЕЛ 3. СВЯЗИ ИСКОПАЕМЫХ НАСЕКОМЫХ С СОВРЕМЕННЫМИ.. 16
РАЗДЕЛ 4. НАСЕКОМЫЕ И ИСТОРИЯ ЖИЗНИ НА ЗЕМЛЕ. 27
4.1 Древние крылатые насекомые. 27
4.2 Увеличение разнообразия наземных сообществ. 29
4.3 Обновление фауны насекомых. 29
4.4 Современная эволюция насекомых и цветковых растений. 30
4.5 Древние реликтовые фауны.. 38
ВЫВОДЫ.. 40
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ.. 42
Насекомые прошли долгий путь эволюции, прежде чем стали такими, какими мы их видим сегодня. Примитивные предки насекомых существовали уже 350 млн. лет назад, в девонском периоде, и, вероятно, еще много раньше. Эволюционная история насекомых была тесно связана с историей других животных и растений, постепенно осваивавших сушу и открывших для насекомых возможности приспособления к наземным местообитаниям. В новых экологических условиях у насекомых мало-помалу выработались морфологические, физиологические и поведенческие адаптации, которые позволили им пожалуй, успешнее, чем любой другой группе наземных животных, занять множество небольших, весьма специализированных экологических ниш. Филогенез современных отрядов насекомых и родственные отношения между ними сложны и отражают ряд важных и интересных этапов эволюции.
Начиная с зарождения жизни на Земле, в течение многих миллионов лет на нашей планете возникали и вымирали разнообразнейшие группы животных и растений, многие из них известны по ископаемым остаткам, а многие, как предполагают, не оставили никаких следов.
Класс насекомых существует уже более 350 млн. лет, и хотя ряд групп
этого класса вымер, насекомые в целом и в настоящее время
находятся в состоянии биологического прогресса, т. е. продолжают увеличивать свою численность и многообразие.
Цель работы состоит в том, чтобы проанализировать происхождение и эволюционирование насекомых.
Основными задачами работы являются следующие:
1) проанализировать свидетельства эволюции насекомых;
2) охарактеризировать вымершие отряды насекомых;
3) проследить связи ископаемых насекомых с современными;
4) дать характеристику насекомых и истории жизни на Земле.
Существует несколько источников данных, помогающих нам расшифровать эволюционную историю насекомых.
Они дают нам наиболее прямые сведения о прошлой эволюции. Ископаемые находки, хотя и далеко не полные, доставляют очень ценную информацию о том, что происходило в определенные геологические периоды, и тем самым помогают раскрыть ряд важных этапов в филогенезе современных групп насекомых.
Удивительно, что ископаемые насекомые, несмотря на их относительно малые размеры и хрупкость, обнаружены в очень многих местах. Чаще всего они встречаются там, где есть залежи угля или мелкозернистые осадочные породы, образовавшиеся в спокойных анаэробных (без доступа кислорода) условиях. Вокруг погребенного под осадками насекомого могут формироваться конкреции (рис. 1), предохраняющие его от разрушения. Ископаемых насекомых находят в наземных и озерных отложениях сланцев, в слоях вулканического пепла, торфа и других материалов. В большинстве случаев встречаются не целые насекомые, а их отдельные части. Наиболее часты находки крыльев, которые служат основой для идентификации большинства ископаемых форм. Остатки насекомых уже давно находили в пенсильванских и нижнепермских отложениях центральных районов США; хорошо известны также ископаемые фауны насекомых из четвертичных озерных отложений района Скалистых гор.
В ископаемых углях, особенно относящихся к меловому периоду (120 млн. лет назад) или более ранних, часто встречаются желваки янтаря (образованные из затвердевшей смолы хвойных растений), в которых могут содержаться хорошо сохранившиеся целые насекомые (рис. 2). Существует много местонахождений янтаря; наиболее известен раннетретичный балтийский янтарь из северных районов Европы. Другие местонахождения были найдены в штате Чьяпас на юге Мексики, в Манитобе и западной Канаде, на Аляске, в Бирме и Сибири. Находки насекомых в янтаре известны начиная с раннего мела.
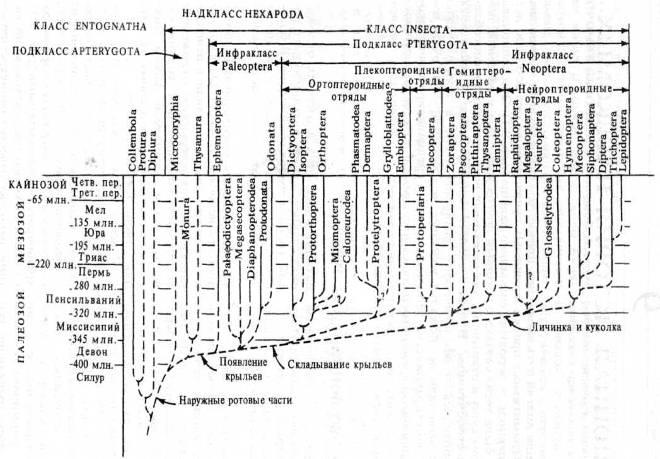
Рис. 1. Филогенетические отношения между отрядами насекомых и находки их ископаемых форм. Сплошными линиями представлены различные ветви в период после их первого появления в палеонтологической летописи; прерывистыми линиями-предполагаемые продолжения этих ветвей в прошлом (по данным сравнительной морфологии и другим косвенными данным). В таблицу не включен ряд предварительно описанных, но недостаточно изученных вымерших отрядов. Млн.-миллионов лет назад.
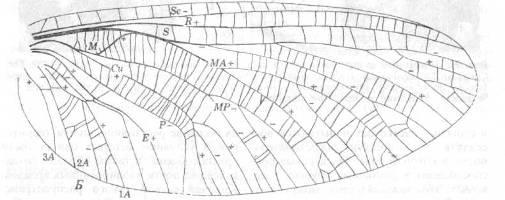
Рис. 2. А.
Фотография железорудной почки с отпечатком крыла древней поденки Lithoneura
mirifica
(MazonCreek в шт. Иллинойс). Б.
Рисунок жилкования крыла. (По Carpenter.)
Вторым, не столь прямым источником данных служит анализ морфологических сходств и различий современных насекомых, позволяющий установить филогенетические отношения. При таком анализе ищут примитивные черты в строении насекомых и пытаются воссоздать ту логическую последовательность, в которой появлялись новые приспособления в различных группах насекомых. Этот важный подход может сильно облегчить интерпретацию ископаемых находок.
Третий источник данных - это изучение современного географического распространения групп насекомых и анализ их филогенетических взаимоотношений в связи с их расселением в прошлые эпохи. Для понимания распространения насекомых наиболее важны три биогеографические концепции. Согласно одной из них, некоторые насекомые расселялись на большие расстояния, минуя территории, к условиям которых они не были приспособлены, и таким образом создавался ряд почти изолированных ареалов. Крайний случай такого распространения -фауна островов Тихого океана. Согласно второй концепции, географическое распространение многих современных насекомых - это результат разделения некогда обширных ареалов на более мелкие. Причиной дробления первоначальных континентов и передвижения их частей в мезозойское и кайнозойское время обычно считают перемещение литосферных плит и раздвигание дна океанов. Третья концепция объясняет распространение большинства современных насекомых влиянием экологических факторов, таких, как, например, изменение климата в четвертичный период, которое привело к перемещению и изоляции отдельных частей прежних ареалов.
Изучение географических изменений в сопоставлении с данными об изменявшемся распространении насекомых в прошлом - еще один важный источник сведений. Примером может служить изучение насекомых пенсильванского периода, проведенное Дёрденом (
Durden, 1974), который обнаружил выраженный широтный градиент разнообразия насекомых по обе стороны от палеоэкватора той эпохи. Подобные исследования могут существенно помочь в понимании происхождения различных групп насекомых.
2.1 Подкласс
Apterygota-первичнобескрылые
Отряд MONURA - однохвостки
Примитивные бескрылые насекомые, сохранившие тергальные рудименты в лабиально-мандибулярной серии головных придатков (рис. 3). Подобно Microcoryphia (или Archeognatha), однохвостки имели 10 полных брюшных сегментов с парными придатками, но у них отсутствовали или были лишь рудиментарными церки на 11-м сегменте брюшка. Отряд описан по роду Dasyleptus
,
представители которого известны из верхнего карбона (Франция, штаты Нью-Мексико и Иллинойс в США) и нижней перми (Чехословакия, штаты Канзас и Оклахома).

Рис. 3. Однохвостка Dasyleptus
brongniarti
из нижнепермских отложений Кузнецкого бассейна в СССР. (По Шарову, из Родендорфа, 1962.)
2.2 Подкласс
Pterygota- крылатые насекомые
2.2.1 Инфракласс
Paleoptera – древнекрылые
Отряд Palaeodictyoptera
Это большая и весьма разнообразная группа, широко представленная в пенсильвании и перми. К этому отряду относятся некоторые из самых крупных ископаемых насекомых. Это были массивные (рис. 2, 4) формы с размахом крыльев нередко до 20 см, а иногда и более 50 см. Голова маленькая с тонкими антеннами, состоящими из многочисленных мелких члеников. Глаза большие и выпуклые. Ротовые части крупные, колющего типа-хоботок, приспособленный для высасывания жидкостей, вероятно, из растений.
Ноги у этих насекомых были короткими и крепкими, напоминающими ноги Ephemeroptera. Переднегрудь несла пару лопастей, чаще сердцевидных, присоединенных к переднеспинке. Эти лопасти были сильно склеротизированы, у многих форм они были крупными и заходили сверху за основания передних крыльев. Лопасти имели систему жилок, гомологичную жилкованию крыльев.
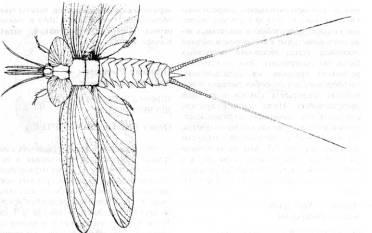
Рис. 4. Stenodictya
-
представитель отряда Paleodictyoptera из позднепенсильванских отложений Франции. (Из.Кика1оуа, 1969-1970, Psyche.)
Крылья палеодиктиоптер были очень разнообразны по размерам и форме (рис. 4). Базальные пластинки крыльев такого же типа, как у Protodonata и Odonata. Крылья обычно с цветными окантовками и круглыми пятнами, часто покрытые волосками, особенно вдоль жилок у основания крыла, по его краю и на крыловой мембране.
Строение относительно короткого брюшка этих насекомых такое же, как и у нимф Ephemeroptera. Церки, состоящие из большого числа сегментов, примерно вдвое длиннее брюшка. У самок-длинный крепкий яйцеклад, у самцов-короткие членистые класперы и парный эдеагус. Представители отряда обитали главным образом в низких влажных тропических и субтропических лесах и болотах и достигали высокой численности.
Отряд Megasecoptera
Отряд этот четко выделяется среди отрядов насекомых, относящихся к раннему Пенсильванию, особой системой жилкования и специфической серповидной формой крыльев (рис. 5). Размеры тела от средних до крупных (до 12 см), тело тонкое с длинными церками, ротовой аппарат сосательного типа имел вид вытянутого хоботка. От Megasecoptera, вероятно, произошел вымерший отряд Diaphanopterodea. Megasecoptera и Раlaeodictyoptera занимали сходные местообитания. Находки представителей отряда позже триаса неизвестны.
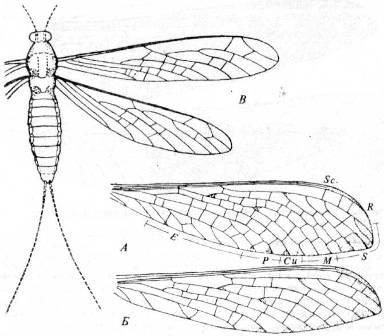
Рис. 5. Отряд Megasecoptera. Переднее (А)
и заднее (Б) крылья Aspidothorax
triangularis
из позднего карбона Европы. (По Carpenter.) В.
Pseudohymen
angustipenni
:
из нижнепермских отложений Урала. (Из Родендорфа и др.)
Отряд Diaphanopterodea
Этот отряд известен только из пенсильванских и пермских отложений. Ротовой аппарат этих насекомых представляет собой сосущий хоботок, состоящий из сильно измененных ротовых частей. Хотя по жилкованию крыльев Diaphanopterodea были в основных чертах сходны с Palaeodictyoptera и Megasecoptera, они могли складывать крылья над телом. Как они это делали, не вполне ясно, но во всяком случае не так, как Neoptera. У пермских представителей жилкование было сильно редуцировано.
Отряд Protodonata (или Meganisoptera) – гигантские стрекозы
Представители этого отряда, жившие с Пенсильвания по триас, были крупными, иногда гигантскими насекомыми с размахом крыльев от 12 до 75 см (рис. 6). Это самые крупные из известных насекомых. По общему облику они были очень похожи на стрекоз-с большими глазами, длинным тонким телом и хорошо развитыми мандибулами. Жилкование крыльев у них было примитивное - имелись все крупные жилки, кроме субнодальной, а жилки Р
и Е
были разделены у оснований. Стрекозы произошли от Protodonata в Пенсильвании. По всей вероятности, Protodonata были быстрыми и агрессивными хищниками.
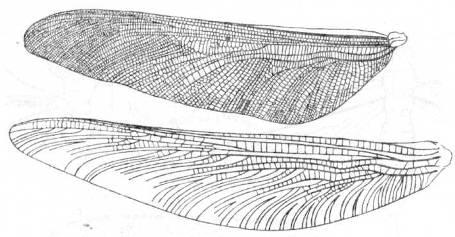
Рис. 6. Представители отряда Protodonata. (Из Родендорфа и др.; А - по
Handlirsch, 1906-1908; Б - по
Мартыновой.)
A
.
Meganeura
moyni
из верхнепенсильванских отложений Франции. Б.
Arctotypus
sinuatus
из верхнепермских отложений.
2.2.2 Инфракласс
Neoptera – Новокрылые
Ортоптероидные отряды
Из вымерших позднепалеозойских ортоптероидных насекомых хорошо известны четыре отряда: Protorthoptera, Miomoptera, Caloneurodea и Protelytroptera.
Отряд Protorthoptera
Эти насекомые жили в период от раннего пенсильвания до перми и относятся к наиболее обычным среди встречающихся в ископаемых находках. Отряд содержит много разнообразных групп; он самый ранний из известных ископаемых
Neoptera. У Protorthoptera были кожистые передние крылья и увеличенная анальная область на задних крыльях (рис. 7,А), а также хорошо развитый жующий ротовой аппарат.
Представители одной из больших групп - Protoblattoidea - внешне похожи на тараканов, хотя отличаются от них рядом признаков, в том числе жилкованием крыльев (рис. 7,Б). Еще одна группа, жившая в раннем Пенсильвании, сохранила примитивный архедиктион с семью анальными жилками на обеих парах крыльев.
Отряд Miomoptera
Отряд включает мелких и очень мелких насекомых с относительно длинными крыльями (рис. 8). Они еще редки в раннем Пенсильвании, но обычны в отложениях пермского периода. По жилкованию крыльев сходны с Psocoptera; имели жующий ротовой аппарат (с мандибулами) и короткие перки.
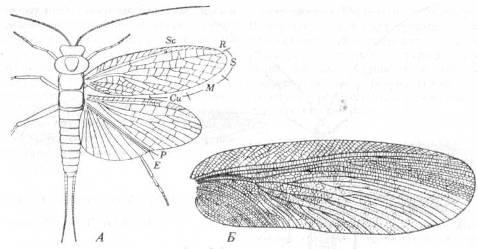
Рис. 7. Отряд Protorthoptera
.
(Из Родендорфа и др.; А - по
Carpenter; Б - по
Handlirsch, 1906-1908.)
A
.
Liomopterum
ornatum
(подотр. Paraplecoptera) из нижней перми Северной Америки. Б.
Крыло Stenoneura
fayoli
(подотр. Protoblattodea) из пенсильванских отложений западной Европы.

Рис. 8. Передние крылья представителей отряда Miomoptera. (По Tillyard; из Родендорфа и др.)
A
.
Delopterum
incertum
из верхнепермских отложений Урала. Б.
Permembria
delicatula
из нижнепермских отложений Северной Америки.
Отряд Caloneurodea
Эти насекомые жили в пенсильванско-пермское время и имели длинные, почти одинаковые передние и задние крылья (рис. 9), у которых базальные отрезки жилок М
и Си
слились, как у некоторых Protorthoptera. Ротовой аппарат был мандибулярного типа, церки-чрезвычайно короткие.
Отряд Protelytroptera(или Protocoleoptera)
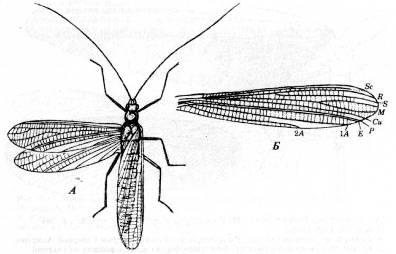
Рис. 9. Отряд Caloneurodea.
A
.
Paleuthygramma
tenuicorne
из нижнепермских отложений Урала. Б.
Переднее крыло Euthygramma
parallelum
из пермских отложений Архангельской области. (Из Родендорфа и др.)
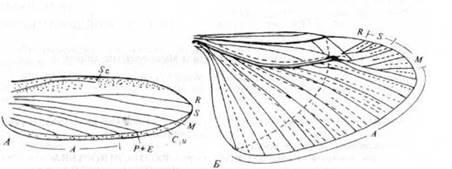
Рис. 10. Отряд Protelytroptera: крылья Protelytron
permianum
из нижнеперских отложений Северной Америки.
А.
Переднее крыло (из Carpenter). Б.
Заднее крыло (из Tillyard).
Еще один отряд мелких насекомых, известный из пермских отложений. У типичных представителей передние крылья превращены в хорошо развитые элитры; задние крылья широкие, с продольными и поперечными складками в расширенной анальной области (рис. 10). У этих насекомых были короткие толстые антенны, широкое туловище, сильные ноги и короткие церки. Судя по жилкованию крыльев, они, вероятно, были предками Dermaptera, а образование элитр у них считают независимым от развития элитр у Coleoptera.
Плекоптероидные отряды
Отряд Protoperlaria
Этот отряд, также известный только из пермских отложений, очень близок к Plecoptera (рис. 11). Передние крылья длиннее задних, у которых пять анальных жилок располагаются так, как у современных Plecoptera. У некоторых Protoperlaria была пара рудиментарных плоских лопастей, отходящих в горизонтальном направлении от переднегруди. Церки длинные; задние ноги толстые, длиннее передних и средних. По-видимому, от Protoperlaria произошли в доперм-ское время Plecoptera.
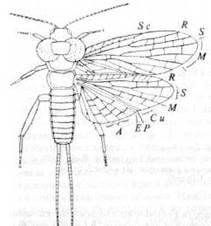
Риc. 11. Отряд Protoperlaria: Lemmatophora
typa
из нижнепермских отложений Канзаса, США. (Из Riek, 1970, с изменениями; по Carpenter.)
Нейроптероидные отряды
Отряд Glosselytrodea
Эта группа, единственная, вымершая среди эндоптериготных Neoptera, относится к пермско-юрскому времени. Жилкование крыльев (рис. 12) более специализированное, чем у Neuroptera или Mecoptera, а передние крылья тоже превращены в элитры. По многим признакам крылья, голова и грудь у Glosselytro-dea были сходны с таковыми Neuroptera. Возможно, что оба этих отряда имели близкого общего предка в позднем карбоне или ранней перми.

Рис. 12. Отряд Glosselytrodea.
А.
Переднее крыло Archoglossopterum
shoricum
из пермских слоев Кузнецкого бассейна. Б.
Переднее крыло Jurina
marginata
из пермских отложений Архангельской области. (Из Родендорфа и др.)
Paleoptera
Находки ископаемых насекомых, принадлежащих четырем вымершим и двум современным отрядам Paleoptera, свидетельствуют о том, что два выживших отряда - Ephemeroptera (поденки; рис. 13, А
и Б)
и Odonata (стрекозы) - это всего лишь остатки древней и очень разнообразной фауны крылатых насекомых Пенсильвания. Современные отряды представляют собой две разошедшиеся ветви Paleoptera. У Ephemeroptera способность к полету как на преимагинальной, так и имагинальной стадии развития, а также редукция ротового аппарата у этих стадий, произошедшая в послепалеозойское время, говорят об их давнем отделении от других Paleoptera.
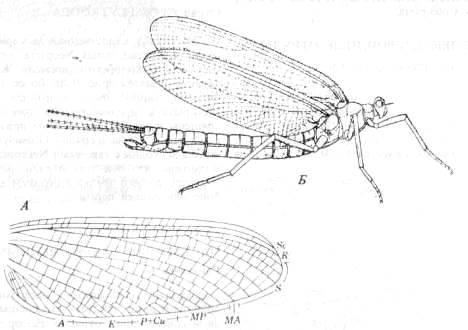
Рис. 13. Ранние представители отряда Ephemeroptera.
A
.
Triplosoha
pulchella
из верхнепенсильванских отложений Европы (из Riek, 1970, с изменениями; по Edmonds, Traver). Б.
Protereisma
регтшпит
из нижней перми Северной Америки (из Родендорфа и др., по Tillyard).
Odonata мало изменились со времени своего появления, и, возможно, их нимфы и взрослые особи всегда были хищниками. По строению ротового аппарата Protodonata сходны с активными, быстрыми стрекозами, но тело у них было значительно более массивное. Судя по облику ископаемых нимф Palaeodictyoptera и Megasecoptera, можно предположить, что они были наземными насекомыми, а их взрослые формы-растительноядными. Значение Paleoptera состоит в том, что они стали предками одной, а возможно, и двух групп насекомых, обладающих способностью складывать крылья над брюшком. Это обеспечивало крыльям защиту, когда они не использовались для полета, и значительно увеличивало подвижность особей.
Neoptera
Наиболее важной группой, происходящей от ранних палеоптер, является группа Neoptera. Крылья этих насекомых благодаря сложной системе крыловых сочленованных склеритов могут складываться на спине вдоль тела. Совершенно иначе устроены крыловые сочленения у двух современных отрядов Paleoptera.
Ортоптероидные отряды
Среди ортоптероидных отрядов наиболее примитивными считаются Dictyoptera (тараканы) и Isoptera (термиты), и жилкование крыльев у этих отрядов в общих чертах сходно. Первый из них был хорошо представлен в Пенсильвании настоящими тараканами (подотряд Blattaria), тогда как самые ранние находки термитов относятся к мелу. В крыльях Protorthoptera из пенсильванских и пермских отложений обнаружены вариации в числе анальных жилок, различия между передними и задними крыльями, разные способы складывания крыльев. Наиболее примитивные Protorthoptera из раннего Пенсильвания являются самыми древними из известных крылатых насекомых, жилкование их крыльев представляло собой типичный архедиктион. Protelytroptera, по-видимому, имели общего предка с Dermaptera (уховертками); последние известны со средней юры.
Orthoptera (саранчовые) и, возможно, также Phasmatodea (палочники), Miomoptera и Caloneurodea представляют собой ветви, рано отделившиеся от одной или нескольких подгрупп внутри Protorthoptera. Phasmatodea известны из триасовых отложений Австралии. Представители Miomoptera и Caloneurodea, первые находки которых относятся к пенсильванскому периоду, были уже настолько специализированы, что их филогенетические связи трудно с точностью установить. Строение тела и крыльев сближает их с ортоптероидными отрядами. Orthoptera впервые появились в отложениях пенсильванского периода, и с тех пор жилкование крыльев у них изменилось лишь незначительно.
Еще один ортоптероидный отряд, представленный в палеонтологической летописи, это Embioptera (эмбии). Один род эмбий был найден в пермских отложениях восточной Европы (рис. 14).
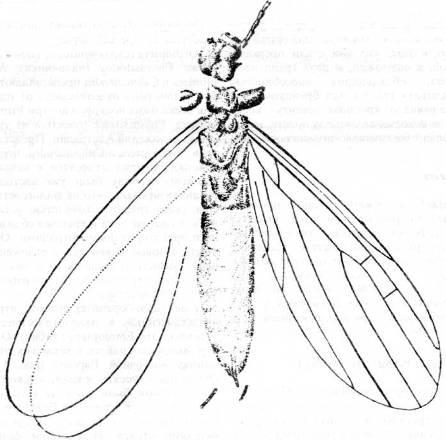
Рис. 14. Отряд Embioptera: Sheimia
sojanensis
из пермских слоев Архангельской области (По Родендорфу и др.)
Остальные относятся к кайнозойской эре и в основном были найдены в балтийском янтаре; эти формы в большинстве своем сходны с современными представителями отряда. Ископаемые формы Grylloblattodea - весьма специализированного отряда - неизвестны.
Плекоптероидные отряды
Есть весьма достоверные находки Protoperlaria и Plecoptera (веснянки) из отложений пермского периода. Эти два отряда насекомых, нимфы которых жили в воде, представляют собой небольшую четкую ветвь, отделившуюся от Protorthoptera. Веснянки известны из пермских, юрских, меловых и кайнозойских отложений.
Гемиптероидные отряды
Все ископаемые гемиптероиды принадлежат к современным отрядам, среди которых только у Zoroptera и Phthiraptera (вши) неизвестны ископаемые формы. Hemiptera (клопы) и Psocoptera (сеноеды) хорошо представлены в пермских отложениях, а появление Thysanoptera (трипсов) можно с достоверностью отнести к юрскому периоду. Все пермские Hemiptera (рис. 15) принадлежали к Homoptera (равнокрылым) и имели хоботок сосущего типа, характерный для этого подотряда. Вымершие семейства пермских Hemiptera весьма разнообразны, иногда с длинными яйцекладами, что указывает на их родство с некоторыми древними прямокрылыми. Они отлично приспособились к жидкой пище и, видимо, постепенно вытеснили палеодиктиоптер из экологических ниш с пищей такого рода.
У раннепермских Psocoptera, относящихся к подотряду Permopsocida (рис. 16, А),обе пары крыльев были почти одинаковыми, с простым жилкованием. К концу пермского периода по меньшей мере у двух семейств отряда Psocoptera уже были редуцированные задние крылья и видоизменено жилкование передних крыльев (рис. 16, Б);
эти формы были удивительно похожи на современных сеноедов. Ископаемых Thysanoptera немного, наиболее известны находки из юрских отложений Средней Азии. Известны также находки насекомых, похожих на трипсов, из верхнепермских отложений в Европейской части.
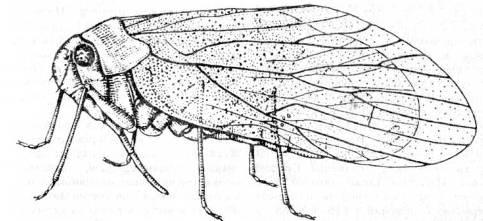
Рис. 15. Отряд Hemiptera: Permocicada
integra
из пермских отложений Архангельской области. (По Родендорфу и др.)
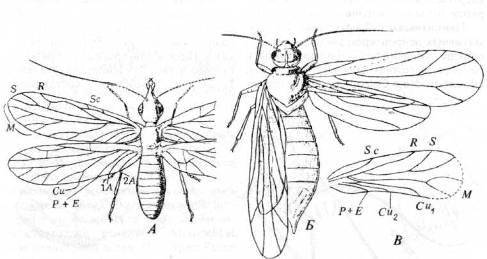
Рис. 16. Представители отряда Psocoptera.
A
.
Dichentomum
tinctum
(подотр. Permopsocida) из нижнепермских отложений Канзаса, США (по Carpenter). Б.
Zoropsocus
tomiensis
(подотр. Parapsocida) из пермских отложений Архангельской области. В.
Lophioneurodes
sarbalensis
(подотр. Parapsocida) из нижней перми, Кузнецкий бассейн (Б
и В
по Родендорфу и др.).
Нейроптероидные отряды
Эти отряды очень хорошо представлены ископаемыми формами. Обе главные ветви нейроптероидных отрядов впервые появились в пенсильванском периоде и были многочисленны в перми.
Примитивные нейроптероиды.
Из примитивных нейроптероидов отряды Megaloptera (большекрылые), Neuroptera (сетчатокрылые) (рис. 16), Glosselytrodea и Coleoptera (жуки) появились в пермском периоде. Появление Raphidioptera (верблюдки) (рис. 17) можно с достоверностью отнести к юрскому периоду. Обилие и видовое разнообразие представителей этих отрядов были значительно больше в пермском, триасовом и юрском периодах, чем сейчас.
Большинство семейств сформировалось еще до окончания юрского периода. Жуки-единственный отряд из примитивных нейроптероидов, освоивший множество наземных местообитаний, и в настоящее время они составляют почти 40% всех известных видов насекомых. У пермских ископаемых Coleoptera, относившихся к подотряду Archostemmata, уже появились признаки жилкования, характерные для элитр. Archostemmata доминировали среди Coleoptera до юрского периода, когда появилось большое число современных подотрядов и семейств.

Рис. 17. Отряд Neuroptera: Mesypochrysa
latipennis
из юрских отложений Казахстана (Из Родендорфа и др.)
Высшие нейроптероиды.
Сюда относятся Hymenoptera (осы и пчелы), образующие независимую ветвь; Mecoptera (скорпионовые мухи), Siphonaptera (блохи) и Diptera (двукрылые), образующие вторую ветвь; Trichoptera (ручейники) и Lepidoptera(бабочки), составляющие третью ветвь. Формы, близкие к Trichoptera, и представители Mecoptera известны из пермских отложений, что указывает на еще более раннее отделение этих ветвей, может быть в пенсильвании. Hymenopteга впервые появились в триасовых слоях. В пермском периоде Mecoptera были уже достаточно дифференцированным отрядом и включали не менее трех подотрядов. У одного из них, Protomecopteга, жилкование крыльев было сходно с жилкованием у ископаемых насекомых, похожих на Megaloptera и живших примерно в то же время, и у их потомков-представителей семейства Meropeidae, ныне реликтовой ветви. Подотряд Еmnеcoptera представлен в пермских отложениях восемью или более семействами (рис. 18), большинство из которых вымерли к концу мезозоя или, претерпев сильные изменения, образовали современные семейства. Две группы Eumecoptera, составлявшие две четкие самостоятельные ветви на протяжении всего мезозойского периода, дали начало двум современным ветвям: 1) семейству Bittacidae и 2) семействам Panorpidae и Рапог-podidae. Мезозойский подотряд Paratrichoptera существовал в триасе, но в кайнозое уже не встречался. Пермские и мезозойские Mecoptera были намного более разнообразны, чем современные Mecoptera, и от них произошли два специализированных отряда - Siphonaptera и Dipteга.
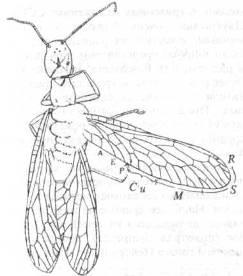
Рис. 18. Отряд Raphidioptera: Mesoraphidia
pterostigmalis
из юрских отложений Казахстана. (Из Родендорфа и др., по Handlirsch 1906-1908.)
Ископаемых остатков Siphonaptera (блохи) мало, сюда относятся находки из нижнемеловых отложений Австралии и раннечетвертичнаго янтаря (рис. 19) из Прибалтики. Отсутствие крыльев и выработка адаптации к паразитическому образу жизни и кровососанию не дают возможности четко установить происхождение блох в пределах группы Mecoptera.
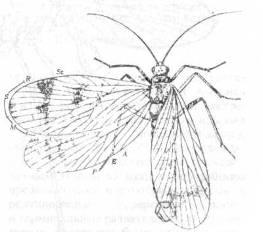
Рис. 19. Agetochorista
tillyardi
-
представитель пермских Mecoptera (подотр. Раrаmеcoptera) с Урала. (Из Родендорфа и др., по Handlirsch, 1906-1908.)
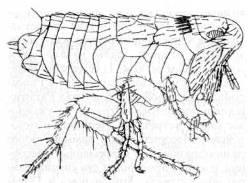
Рис. 20. Блоха Palaeopsylla
clebsiana
из позднеэоценового балтийского янтаря. (Из Родендорфа и др.)
В отряде двукрылых больше ископаемых форм (рис. 20). Крылья, сходные с крыльями Diptera, находили в пермских отложениях Австралии и, возможно, в триасовых отложениях. Двукрылые, очень близкие к современным, известны из ранней юры, где были широко представлены Nematocera и ряд семейств Brachycera. В эоцене и в более ранних слоях встречались представители большинства семейств двукрылых. Представители семейств высших мух из группы Schizophora появляются в средне- и верхнекайнозойских отложениях.
Hymenoptera (пилильщики, муравьи, пчелы, осы) очень часто встречаются среди ископаемых насекомых мезозоя и кайнозоя. Наиболее древние из них, примитивные пилильщики из семейства Xyelidae (подотряд Symphyta), известны из нижнего триаса Центральной Азии, верхнего триаса Австралии и юрских отложений в Азии (рис. 21, А).
Представители подотряда Apocrita впервые обнаружены в юре, а паразитические Ichneumonoidea (рис. 21, Б)
и Chalcidoidea хорошо представлены в мелу и более поздних слоях (рис. 21, В).
Sphecoidea известны из нижнемеловых отложений Австралии. Formicidae (муравьи) и Vespidae (осы) были многочисленны в начале кайнозоя, aApoidea (пчелы) в олигоцене. По-видимому, последовательность появления различных ископаемых форм Hymenoptera отражает некоторые этапы эволюции этого отряда.
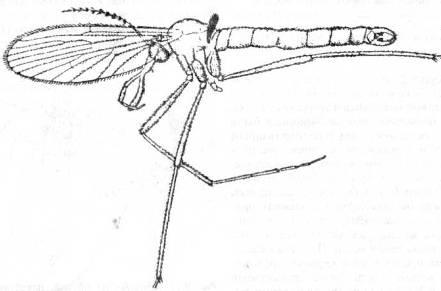
Рис. 20. Двукрылое Macrochile
spectrum
из позднеэоценового балтийского янтаря.
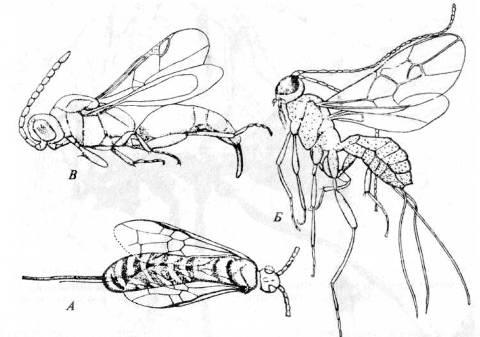
Рис. 21. Отряд Hymenoptera. A. Anaxyela gracilis
(подотр. Symphyta из юрских отложений в Казахстане). Б.
Microtypus
longicornis
(подотр. Apocrita, надсем. Ichneumonida) из эоценового балтийского янтаря; В.
Cryptoserphus
pinorus
(сем. Proctotrupidae) из эоценового балтийского янтаря. (Из Родендорфа и др.)
Trichoptera (ручейники) и Lepidoptera (бабочки) представляют собой еще одну сильно продвинувшуюся в своей эволюции группу нейроптероидных насекомых. Trichopteraили их непосредственные предшественники известны из ранне-пермских отложений Северной Америки и из позднепермских отложений Австралии. По характеру жилкования крыльев они были сходны с древними Mecoptera, что указывает на их общее происхождение. Пермские ручейники были промежуточным этапом на пути к мезозойским представителям этого отряда, у них были редуцированы поперечные жилки и терминальные разветвления. В верхнем триасе Австралии были найдены представители семейства Mesopsychidae, а в верхнем триасе и нижней юре северозападной Европы - представители семейства Necrotaulidae. В этих семействах досих пор сохранились самостоятельные основания у жилок М
и Сu1
. У более поздних представителей отряда основания этих двух жилок слиты. Большая часть других ископаемых находок ручейников относится к меловому периоду и олигоцену. В ископаемых находках Trichoptera довольно часто сохраняются домики личинок, конструкция которых сложилась, по-видимому, уже к триасовому периоду. Lepidoptera отряд, хорошо всем известный из-за огромного разнообразия крупных, ярко окрашенных форм, эволюционно очень молод и относительно скудно представлен в ископаемых находках. Известно несколько примитивных семейств из меловых отложений, однако большинство ископаемых бабочек - из эоцена и олигоцена северной Европы (балтийский янтарь), а несколько из олигоценовых отложений Северной Америки. Подотряд Jugatae был уже хорошо представлен к началу кайнозоя. По жилкованию крыльев его представители сходны с некоторыми ручейниками и, вероятно, произошли от ручейников в среднемезозойское время. У более высокоразвитых Frenatae,известных из балтийского янтаря, передние и задние крылья уже не были одинаковыми. Самый прогрессивный подотряд Rhopalocera вообще не имеет ископаемых форм.
Миссисипский и пенсильванский периоды. В верхнедевонских и миссисипских отложениях ископаемых насекомых до сих пор найдено не было. Однако быстрая эволюция амфибий с разделением
их на четыре отряда в течение миссисипского периода могла быть связана с использованием в качестве пищи пресноводных членистоногих, в особенности личинок насекомых.
Мелководные континентальные шельфы были покрыты широкими речными дельтами, где росли густые леса, состоявшие из псилофитов, древовидных плаунов, хвощей и родственных форм, семенных папоротников и ранних форм хвойных (кордаитов и древних Conifer
ales
).
Эти леса были источником образования крупнейших залежей каменного угля, а также многих отложений, богатых ископаемыми остатками насекомых. После середины пенсильванского периода в бассейнах горных озер возникло много болотистых лесов.
Фауна насекомых в пенсильванский период состояла не менее чем из 12 отрядов; судя по отдельным фрагментарным находкам, были еще и другие отряды. Известно около 35 значительных мест нахождения ископаемых, и собранный там материал показывает, что Paleoptera, в особенности представители отрядов Paleodictyoptera, Megasecoptera и Diaphanoptera, достигали высокой численности и разнообразия. Весьма многообразны были и Neoptera, особенно многочислен был отряд Protorthopteга. Отряды Miomoptera, Caloneurodea и Orthoptera тоже отличались большим разнообразием форм. Другую большую группу пенсильванских насекомых, включавшую более двух десятков семейств, составляли тараканоподобные насекомые, в том числе подотряд Protoblattoidea (Protorthoptera). Из-за обилия находок этих форм пенсильванский период называют «веком тараканов». С точки зрения палеогеографии представляет интерес постепенное продвижение мест находок пенсильванских насекомых к северу и югу от первоначального их расположения в виде узкого экваториального пояса.
Пермский период.
Суперконтинент Пангея простирался от северных полярных областей через умеренные, субтропические и тропические зоны до южной полярной области. Пермские болота оставили после себя много угольных пластов и не менее 75 мест крупных находок ископаемых насекомых.
Почти все пенсильванские отряды насекомых перешли в пермские слои, и разнообразие их форм продолжало возрастать (рис. 9.36); кроме того, появилось не менее 10 новых отрядов, которые стали доминировать в находках. Были хорошо представлены Psocoptera (сеноеды), Hemiptera (клопы), Mecoptera (скорпионовые мухи) и Соleoptera (жуки). Впервые во время пермского периода появляются настоящие Odonata (стрекозы), Protelytroptera, Pro-toperlaria, Plecoptera (веснянки), Neuroptera (сетчатокрылые), Glosselytrodea, Trichoptera (ручейники) и, возможно, Меgaloptera (большекрылые). Многие из этих древних отрядов к концу периода вымерли, но 10 или большее число отрядов образовали исходные ветви, от которых произошли современные отряды.
В пермском периоде быстро развивались также две другие группы наземных организмов-рептилии и высшие растения. Рептилии, появившиеся лишь в конце пенсильванского периода, в перми быстро достигли расцвета. Среди них были травоядные, хищные и много насекомоядных форм. Увеличивалось и разнообразие растений. Во влажных стациях доминировали споровые и семенные папоротники, однако многие древние кони-ферофиты заселяли все более и более сухие местообитания.
Триасовый период. В
триасе происходили большие изменения в условиях жизни на суше, и это отражалось в эволюционной истории насекомых. Большие области суперконтинента Пангеи становились теплыми и сухими и покрывались красными аллювиальными равнинами. Приспосабливаясь к этим новым условиям, быстро изменялась растительность. Кониферофиты были представлены многими формами Cordaitales, Co-niferales и Taxales. В теплых умеренных зонах широко распространялись Gink-goales и, возможно, Gnetales, а в тропических и субтропических областях - Суса-deoidales и цветущие Cycadales. Все эти изменения в условиях жизни привели к важным эволюционным изменениям и у насекомых.
Мест, где в триасовых отложениях были найдены ископаемые насекомые, значительно меньше, чем мест с находками из более ранних или более поздних отложений. Известно около десятка таких мест, в основном в Австралии и Центральной Азии, но только в некоторых из них оказалось много хорошо сохранившихся насекомых. По меньшей мере девять отрядов, появившихся в пермский период, не пережили триас. В триасовых отложениях впервые появились примитивные Hymenoptera (пилильщики), Phasmatodea (палочники) и Diptera (двукрылые); однако высокая степень дифференциации этих групп, вероятно, указывает на более раннее возникновение. Личиночные ходы, столь типичные для современных жуков из семейств Ви-prestidae и Scolytidae (рис. 9.37), до сих пор сохранились в окаменевших деревьях из триас 'вых слоев.
Юрский период. В юре Пангея оставалась почти без изменений. Вдоль западного края Северной Америки от Мексики до Аляски -протянулась в основном в меридиональном направлении горная система. Наносы и ветры, которые дули с этих гор, а также трансгрессии и регрессии внутренних морей сильно влияли на климат Северной Америки.
В юрский период быстрыми темпами продолжалась эволюция высших растений. В отложениях, относящихся к концу этого периода, встречаются пыльцевые зерна, сходные с пыльцой двудольных покрытосеменных. Семенные папоротники были в это время редкими и не пережили юру. Кониферофиты, особенно Ginkgoales и Cycadales, были широко распространены.
Насекомые юрского периода известны из нескольких хорошо изученных источников, к которым относятся нижнеюрские отложения северо-западной Европы, несколько мест со среднеюрскими отложениями вблизи гор Каратау в Азии и несколько в Иркутской области в Азиатской части Европы. В этих юрских фаунах появилось много «современных» новых подотрядов и надсемейств в отрядах Neuroptera, Mecoptera и Diptera. Разнообразная флора предоставляла насекомым обильную пищу. У некоторых растений уже были хорошо заметные цветки, и вполне возможно, что насекомые посещали и опыляли их. Впервые в юре появились Dermaptera (уховертки) и настоящие Thysanoptera (трипсы), и в целом фауна насекомых этого периода выглядела вполне современной. То, что в юрском периоде уже были млекопитающие, летающие рептилии и птицы, позволяет предполагать, что и Phthiraptera (вши), и Siphonaptera (блохи) уже могли в то время паразитировать на соответствующих хозяевах.
Меловой период. Продолжительность мелового периода составляет 60-70 млн. лет. В течение этого времени бассейн Северо-Атлантического океана расширялся, постепенно отделяя южные районы Европы от Северной Америки и обе эти области от северной Африки и северной части Южной Америки. Бассейн Южно-Атлантического океанастал в это время расширяться, так что Африка и Южная Америка начали отделяться друг от друга примерно с середины мелового периода. На протяжении всего периода между Северной Америкой и Европой существовала материковая связь через северную Европу, шельф Баренцева моря и Гренландию. Высокие горные цепи Кордильер по-прежнему оказывали сильное влияние на климат прилежащих областей.
В этот период быстро эволюционировали покрытосеменные растения, особенно двудольные. К концу периода мы находим уже почти все современные семейства двудольных. Появляются осоки и злаки. Это обогащение фауны цветковых растений, многие из которых опылялись насекомыми, сопровождалось увеличением числа групп насекомых, использующих эти растения в качестве источника пищи; возник, в частности, ряд новых семейств бабочек, питающихся пыльцой и нектаром. Ископаемых насекомых из меловых отложений (рис. 22) известно не так много, но все же такие находки обнаружены в ряде мест Аляски, Канады, Сибири и Среднего Востока. Фауна насекомых была довольно разнообразна и включала веснянок, стрекоз, тараканов, мелких двукрылых, тлей, ручейников, паразитических перепончатокрылых, термитов и низшие семейства бабочек.

Рис. 22. Ископаемые насекомые из мелового янтаря (озеро Сидар, пров. Манитоба, Канада). (ИзMcAlpine, Martin, 1969, The Beaver; рисункиК. A. Hamilton, AgricultureCanada.)
А.Хальцида (сем. Mymaridae, отр. Hymenoptera). Б
.
Мелкийхищныйклоп (сем. Anthocoridae, отр. Hemiptera). В
.
Жук (сем. Scydmaenidae, отр. Coleoptera). Г
.
Личинкажука-хищника (сем. Staphylinidae).
Многие из них были либо близкими родственниками современных родов, либо представителями этих родов; другие принадлежали к более примитивным родам. Особенный интерес представляют те семейства юрских насекомых, которые были промежуточными звеньями между современными семействами. В качестве примера можно привести ископаемое семейство Jascopidae, известное по одной нимфе из канадского мелового янтаря; оно заполняет промежуток между семействами Cercopidae и Cicadellidae (из Homoptera), представляя собой раннюю стадию в эволюции Cicadellidae от церкопидоподобного предка.
4.5 Становление современных фаунистических областей
Кайнозойская эра.
Несмотря на небольшую продолжительность кайнозойской эры (60-70 млн. лет), ископаемые свидетельства об энтомофауне этого времени наиболее полны.
Изменения климата.
Современная физическая картина земной поверхности сформировалась в основном в кайнозойскую эру. Происшедшее в кайнозое похолодание и сильные колебания температуры, последовавшие в четвертичном периоде, оказали большое влияние и на современное распространение насекомых. Эти события приводили к частому и значительному расширению и сужению областей с определенными условиями среды, к смещению климатических поясов к экватору или от него; в мире насекомых они вызывали многократное образование изолированных и реликтовых ареалов. Хотя подобные экстремальные климатические изменения могли происходить и раньше, в пенсильванское время и в начале пермского периода, в мезозое столь сильных колебаний климата не было.
Географические изменения.
В кайнозойскую эру продолжалось также отдаление друг от друга и усиление изоляции большинства континентов. Северная Америка и северная часть Европы все еще раздвигались, и к середине эоцена сухопутная связь между ними была прервана. В позднеэоценовое и олигоценовое время по мере продвижения на север континентального блока, включающего Африку, Индию и Средний Восток, теплые прибрежные равнины по южным окраинам Центральной Европы, Центральной Азии и Китая подвергались деформации. Ко времени миоцена Гималайская горная система простиралась почти без разрывов от Европы до южной Азии, и к плиоцену эти горы достигли огромной высоты. С северной стороны этой горной цепи образовался ряд обширных впадин, и на месте многих из них возникли озера неморского происхождения (остатками этих озер являются Черное, Каспийское и Азовское моря). В наше время Гималаи представляют собой климатический и физический барьер между Палеарктикой и Восточной зоогеографической областью.
Австралия и Новая Гвинея тоже двигались на север, заняв положение, близкое к границе с Восточной зоогеографической областью. Резкие контрасты климатических и природных условий в сочетании с относительно недавним передвижением материков ограничивали взаимопроникновение животных и растений Восточной и Австралийской областей.
С юрского периода вплоть до начала плиоцена Северная и Южная Америки были разделены. От Африки и Антарктиды Южная Америка отделилась в конце мелового периода. Панамский перешеек образовался, по-видимому, в середине плиоцена, т. е. 4-6 млн. лет назад. Хотя обмен животными и растениями через этот узкий перешеек шел весьма интенсивно, оба американских континента сохранили ряд четких биотических особенностей. Наибольшие изменения произошли с млекопитающими; североамериканские плацентарные формы в значительной мере вытеснили сумчатых и плацентарных млекопитающих Южной Америки.
Северная Америка и северные районы
Азии на протяжении почти всей кайнозойской эры неоднократно имели связь в области Берингова пролива, и в результате фауны неарктической и палеарктической частей Голарктики стали очень сходны между собой. В Северной Америке значительное увеличение высоты Кордильер и Скалистых гор, происходившее в миоцене, плиоцене и четвертичном периоде, привело к разделению Провинции бассейнов и хребтов (в юго-западной части США) на мелкие, экологически обособленные участки. Провинция бассейнов и хребтов, Скалистые горы и Великие Равнины были подняты на 2-Л
км в виде широкого регионального свода, а последующая эрозия придала этому горному ландшафту современный облик. Это поднятие создало «дождевую тень», что привело к распространению луговых сообществ на Великие Равнины. Горы стали холоднее, и на них стало выпадать больше атмосферных осадков.
Среди ископаемых насекомых из верхнеэоценового и нижнеолигоценового балтийского янтаря есть представители нескольких отрядов и подотрядов, которые не встречались в более ранних отложениях: Mantodea, Embioptera и специализированные паразитические жуки Strepsiptera (веерокрылые). Давно известны находки олигоценовых насекомых из северной Европы (Германия и Франция), Северной Америки, Центральной Азии и с Кавказа; они послужили основой для ряда классических исследований. Миоценовые находки известны из северной Европы и с Северного Кавказа, а также из пустыни Мохаве в Калифорнии. Плиоценовых находок значительно меньше.
Большинство ископаемых насекомых, найденных в балтийском янтаре, принадлежат к современным родам. Некоторые семейства высших Hymenoptera и Lepidoptera в коллекциях насекомых из балтийского янтаря отсутствуют и, вероятно, появились в эоцене. Недалеко от Флориссанта (штат Колорадо) в тонких слоях вулканического пепла, осевшего в водоемах у подножия старых Скалистых гор, сохранилось много ископаемых насекомых из олигоцена. Эти насекомые жили в самых разнообразных экологических условиях, от тропических до умеренно-холодных, и их фауна не уступала по многообразию родов современной фауне. Некоторые из этих родов отсутствуют теперь в Северной Америке, но встречаются в других местах - например, мухи цеце (род Glossina
)
известны сейчас только в Экваториальной Африке. Среди олигоценовых находок из Флориссанта есть интересные златоглазки из семейства Osmylidae, а современные представители этого семейства живут только в тропических областях.
Ледниковые эпохи четвертичного периода (плейстоцен).
После плиоцена во всем мире более прохладные эпохи неоднократно чередовались с чуть более теплыми. Во многих районах, особенно в Антарктике и в высоких северных широтах Америки и Евразии, росли и распространялись льды, которые затем таяли. С севера льды продвигались на юг и покрывали обширные территории. Известно четыре главных оледенения, и после каждого из них наступал долгий период с климатом более теплым, чем в настоящее время. Последние два из этих оледенений, а может быть и предыдущие, включали несколько циклов с чередованием ледниковых и межледниковых периодов. Сейчас мы находимся в регрессивной фазе последнего ледникового периода, и лишь остатки северной ледяной шапки сохраняются ныне в виде изолированных ледников или островных масс, как на Гренландии.
Расселение, дизъюнктивные ареалы и видообразование.
Изменения климата во время смены ледниковых и межледниковых периодов оказали огромное влияние на живую природу Северной Америки, особенно в северных районах, отличавшихся умеренным климатом. В то время как обмен между фаунами Северной Америки и западной Европы отсутствовал, северо-западная часть Северной Америки и северо-восточная часть Азии были связаны по суше Беринговым мостом, который мог служить путем для расселения животных. В результате, когда условия на севере стали такими же, как в наше время, или даже более теплыми, многие виды насекомых, приспособленные к умеренно-холодному климату, расселились по всей Голарктике, распространяясь из Северной Америки в Азию или в обратном направлении. Во время следующего похолодания, при новом увеличении площади ледников ареалы этих видов расширялись к югу, так как южные окраины их ареалов становились холоднее и лучше соответствовали потребностям этих насекомых. Когда северные границы ареалов смещались к югу от Берингова моста, ареал каждого вида распадался на две части, между которыми не было генетического обмена. Если такое разобщение было достаточно долгим, популяции внутри каждой из этих частей могли стать самостоятельными видами. Когда ледники таяли и наступал очередной теплый межледниковый период, ареалы видов перемещались к северу. При этом вновь происходило смешивание азиатских и североамериканских видов, многие из которых были близкородственными, но уже генетически изолированными; в результате формировалась новая группа голарктических видов, а затем весь этот длительный процесс образования новых видов под влиянием климатических факторов повторялся снова.
Такой же механизм лежал в основе видообразования в горных районах в пределах одного континента. Например, холодолюбивый род веснянок Allocapnia
,
распространение которого ограничено холодными ручьями восточной части Северной Америки, дал начало большой группе видов (около 38); этому способствовали прежде всего волны рассеяния, связанные с флуктуациями климата, а затем изоляция географически обособившихся популяций родительских видов в нескольких горных районах-на севере и юге Аппалачских гор, на плато Камберленд и в горах Озарк-Уошито (штаты Миссури, Арканзас и Оклахома). Судя по имеющимся данным, все или почти все эти виды возникли, вероятно, в плейстоцене, т.е. в последние 3 млн. лет.
Ареал северного холодолюбивого вида ручейников Glossosoma
intermedium
в целом сейчас простирается от западной Европы через Азию на восток до Миннесоты и Миссури. На протяжении всего огромного ареала вид поразительно однороден, и это указывает на то, что он распространился по всей Голарктике во время последнего оледенения - возможно, всего лишь 12-30 тыс. лет назад. Однако ареал этого вида разделен на ряд изолированных популяций, что создает условия для образования новых видов в будущем.
В юго-западных районах США благодаря климатическим изменениям в четвертичном периоде возник еще один механизм видообразования. Ледниковые периоды были холодными и дождливыми, а межледниковые - жаркими и сухими. Целый ряд насекомых в этих районах, в том числе многие жуки, осы и пчелы, приспособлен к условиям пустыни. В дождливые эпохи их ареалы были ограничены небольшими изолированными слегка приподнятыми пустынными или полупустынными районами, расположенными между холодными, влажными горными хребтами. В это время такие изолированные, приспособленные к сухому климату популяции становились видами. В следующий межледниковый период сухие жаркие районы расширялись за счет прохладныхи влажных, в результате возникавших контактов происходило расселение и смешение видов.
Благодаря находкам в торфе и богатом органическим веществом иле появляется все больше данных об ископаемых насекомых четвертичного периода. У некоторых насекомых, например у жуков, части экзоскелета настолько прочны, что их можно вымывать из рыхлых отложений, не повреждая. Недавно были изучены находки Coleoptera из позднеплейстоценовых (висконсинских) отложений. Большинство этих ископаемых жуков принадлежит к современным видам жуков из семейств, приспособленных к умеренному климату и, судя по полученным данным, эволюционирующих сравнительно медленно. Перемещения, сужения и расширения климатических поясов в четвертичном периоде имели для насекомых многосторонние последствия, важнейшими из которых были изменения их ареалов и характера расселения ; при этом темпы эволюции не были во всех группах одинаковыми.
Среди современных насекомых, живущих в областях, входивших в древнюю Гондвану, огромное число форм принадлежит к семействам, известным только из Австралии, Новой Зеландии и Южной Америки. Некоторые другие встречаются, помимо указанных районов, также в Южной Африке. Большинство этих насекомых приспособлены к южной умеренной зоне, ведут оседлый образ жизни и не имеют средств для преодоления океанических пространств. К таким насекомым относятся, например, нелетающие растительноядные клопы из семейства Peloridiidae (Hemiptera), долгоносики из семейства Belidae, нелетающие сверчки из семейства Cylindrachetidae, три семейства и одно подсемейство веснянок (Plecoptera). В этот южный фаунистический комплекс входят также примитивные скорпионовые мухи из семейства Nannochoristidae - единственные дожившие до наших дней представители отряда Mecoptera, имеющие холодолюбивых водных личинок. Это семейство известно и из пермских отложений Австралии. Из двукрылых представителями этой фауны являются род Pelecorhynchus
,
родственный толстоножкам (сем. Bibionidae) и слепням (сем. Tabanidae), род Trichophthalma
(сем. Nemestrinidae), подсемейство Ceratomerinae мух-толкунчиков (сем. Empididae) и примитивное семейство комаров Blephariceridae. Есть и другие примеры, указывающие на общность происхождения этой фауны. Большинство таких видов - дожившие до наших дней представители древних примитивных ветвей, и это заставляет предполагать, что когда-то до раскола Гондваны у них были сплошные обширные ареалы. В каком-то смысле эти насекомые-реликтовые, эндемичные и дизъюнктивные в одно и то же время!
Существует несколько гипотез о происхождении насекомых и их взаимоотношениях с другими классами членистоногих. Основные затруднения заключаются в том, что палеонтологи еще не нашли отпечатков ископаемых форм, которые могли бы с достаточным основанием считаться предками насекомых. Тем не менее уже накопленные материалы по ископаемым насекомым позволяют относить время их возникновения к началу девонского геологического периода и в самых общих чертах представить себе существовавшую тогда экологическую обстановку. В девонский период, начавшийся 375 млн. лет назад и продолжавшийся 50 млн. лет, осуществлялся важнейший эволюционный этап развития жизни — выход растений из водоемов на сушу. К началу этого периода на суше растений не было, а жизнь концентрировалась в водоемах, где имелись водоросли и беспозвоночные.
В настоящее время полагают, что в дополнение к многочисленным современным отрядам насекомых прежде, судя по ископаемым остаткам, существовало еще 11 или даже больше отрядов. Большинство исчезнувших отрядов впервые появились в отложениях, относящихся к Пенсильванию или ранней перми, и в основном вымерли уже к концу пермского периода. Среди этих исчезнувших форм были и ранние предки некоторых современных отрядов насекомых. Большинство этих ископаемых предков морфологически были более примитивны и менее специализированы, чем их потомки; поэтому в позднем палеозое различия между группами насекомых еще не были такими четкими, как между соответствующими современными отрядами. С другой стороны, изучение поздних палеозойских насекомых ясно показывает, что к концу Пенсильвания основные этапы эволюции насекомых были уже завершены. Это относится к развитию крыльев, их складыванию, дифференциации способов питания и выработке жизненных циклов с полным превращением. Ниже даются краткие описания тех вымерших отрядов насекомых, которые уже достаточно изучены.
Морфологическая организация, которая характерна для насекомых, обеспечила им фундаментальный морфологический прогресс (ароморфоз). На основе этой организации они освоили воздушно-наземную среду обитания. В дальнейшем насекомые приспособились к самым разнообразным частным средам, в результате чего у них возникли различные конкретные приспособительные адаптации (идиоадаптации). На основе этих адаптации в прошлом и в настоящее время у насекомых происходит интенсивное видообразование.
СПИСОК
ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1) Андрианова Н.С. Экология насекомых: Курс лекций. – М., 1970. – 158 с.
2) Бей-Биенко Г. Я. Общая энтомология. Издание третье, дополненное. М.:Высшая школа, 1980. - 416 с.
3) Грант В. Эволюция организмов. - М., 1980.
4) Жизнь животных: В 6-ти томах. – М., 1985.
5) Майр Э. Популяции, виды и эволюция. М., 1974.
6) Мамаев Б.М. Энтомология. - М.: Мир, 1976. – 426 с.
7) Росс Г. и др. Энтомология: пер. с англ. – М.: Мир, 1985. – 576 с.
8) Савчук М.П. Зоологія безхребетних. – К., 1965. – 503 с.
9) Северцов А.Н. Эволюционная теория. – М., 1967.
10) Щербак Г.Й. Зоологія безхребетних. Кн. 1-3. – К.: Либідь, 1995.
11) Яблоков А.В., Юсупов А.Г. Эволюционное учение. – М.: Высш. шк., 1989. – 335 с.
|