Зміст
Вступ
1. Іксодові кліщі
2 Роди кліщів Ixodes
2.1 Рід Ixsodes
2.2 Рід Dermacentor14
2.3 Рід Haemaphysalis
2.4 Рід Rhipicephalus
2.5 Рід Hyalomma
2.6 Рід Boophilus
2.7 Рід Атblуотта
3 Епідеміологічне значення
Висновок
Література
Вступ
Екологія, що виникла в надрах біології в епоху перемоги дарвінізму і розвитку історичного підходу до аналізу життєвих явищ, швидко завоювала широке визнання і стала впливати на інші біологічні науки. Вивчаючи закономірності взаємин організмів з навколишнім середовищем, екологія початку впливати на систематику тварин і рослин, різні розділи морфології (цитологію, гістологію, ембріологію й ін.), фізіологію і генетику. Виникають екологічна фізіологія, екологічна палеонтологія й інші нові напрямки. У надрах зоології розвивається екологія тварин (з підрозділами: екологія комах, екологія риб, екологія птахів і т.д.), у ботаніку — екологія рослин (екологія водоростей, екологія мохів, екологія грибів і т.д.).
В міру розвитку екології розширилися представлення про рівні організації живих систем, досліджуваних екологією: організм, популяція, вид, біоценоз, екосистема. Екологія вивчає рівні організації живого від організмів до екосистем. Біосфера не може розглядатися як рівень організації живої системи, це лише сфера життя, глобальний біотоп, у якому існують усі екосистеми, біоценози, види, популяції, організми. Але біосфера вивчається екологією також, як нею вивчаються біотопи біоценозів, місцеперебування організмів, різні фактори середовища і середовища життя. У задачу популяційної екології входить вивчення чисельності окремих видів, спалахів їхнього розмноження, закономірностей розміщення видів по території, змін популяцій за часом. Екологія видів (аутекологія) переростає в екологію співтовариств різних видів — біоценологію, а остання — у біогеоценологію, оскільки біоценози можуть існувати лише в єдності з абіотичної середовищем. Сучасна екологія включає екологію особей, екологію популяцій і екологію співтовариств. Основною проблемою екології є проблема чисельності видів. Екологія вивчає взаємодію організмів із середовищем, що обумовлює розвиток, розмноження і виживання особей, структуру і динаміку утворених ними популяцій видів, структуру і динаміку співтовариств різних видів і історично сформовані на їхній основі специфічні пристосування видів; внутрішньовидові відносини і специфічна структура виду; співтовариства популяцій різних видів, різні на різних ділянках земної поверхні, їхні взаємні пристосування, що забезпечують біогенний круговорот речовин.
При вивченні зв'язків організмів з фізико-хімічними умовами середовища, що забезпечують обмін речовин, екологія спирається на дані фізіології. При цьому еколога цікавить не життя виду, а процеси, що відбуваються в організмі, пристосувальні особливості видів, їхні життєві форми. Закономірності розміщення видів по території неможливо вивчити без залучення даних біогеографов, геоботаніків, ландшафтоведів, ґрунтознавців, кліматологів. Таким чином, екологія являє собою науку, зв'язану з поруч біологічних і небіологічних наук, що зберігає в той же час свої специфічні задачі й об'єкти дослідження. Значні екологічні особливості характерні для кровососних комах і кліщів. Особливості їхня метаморфоза, дивергенція в розвитку, що супроводжується різкою зміною умов життя і гетеротропністю, змушують вивчати екологію окремих фаз розвитку. Личинка, лялечка, доросла комаха мають різну морфологію, різні функції і різні вимоги і зв'язки з зовнішнім середовищем. Важливою задачею екології кровососних комах і кліщів є з'ясування основних взаємозв'язків з навколишньою природою і людиною.
Предметом еволюційної екології є мікроеволюційні процеси, що протікають у процесі становлення екологічного гомеостазу в осередкових біогеоценозах. Задачі цього розділу екології полягають у виявленні екологічних механізмів еволюційного процесу, шляхів і форм еволюційного становлення індивідуальних, і видових адаптації в біоценотичних системах (Шилов, 1981).
Задачами екології комах і кліщів (як і інших тварин) є пізнання формування їхньої морфологічної і фізіологічної особливостей і способу життя в залежності від умов середовища, вивчення впливу середовища на чисельність особей даного виду, на характер розподілу їхній по території і на формування співтовариств організмів, що населяють ту чи іншу територію.
Екологія вивчає ті сторони відносин організмів із середовищем, від яких залежить успішність їхнього розвитку, виживання і розмноження. Вона вивчає не тільки безпосередні взаємини із середовищем, але й історично сформовані на їхній основі специфічні для кожного виду пристосування особей, що забезпечують різнобічні зв'язки із середовищем, їхній розвиток і дозрівання; внутрішньовидові відносини і специфічну для виду структуру його популяції, що визначає його спосіб життя (одиночний, стадний, колоніальний), спосіб використання життєвих ресурсів, розмноження і розселення виду, у результаті чого встановлюються рівень чисельності і характер її динаміки; різні в різних ділянках земної поверхні співтовариства популяцій видів, що зв'язані багатьма взаємними пристосуваннями, що забезпечують біогенний круговорот речовин.
Знання способу життя кровососних комах і кліщів у різних умовах середовища створює можливість профілактичних заходів, що перешкоджають їх масовому розмноженню. Знання місць виплоду комарів, бліх, кліщів і інших членистоногих — переносників збудників хвороб людини і тварин дозволяє цілеспрямовано проводити заходу щодо ліквідації цих трансмісивних хвороб.
Паразитологія як екологічна наука характеризується різними рівнями вивчення і самих паразитичних організмів, і паразитно-господарських відносин. Предметом і задачами паразитоценології служать вивчення процесів появ паразитизму в їхньому взаємозв'язку і взаємодії з метою розкриття механізмів, що визначають взаємини біоценотичних угруповань паразитів між собою і з хазяїнами, а також їхня залежність від умов навколишнього середовища. Паразитоценологія виникла на стику приватних наук і будується на основі системного підходу до явищ паразитизму в біосфері. В даний час виділяють п'ять екологічних рівнів паразитологичних досліджень, а саме: аутекологічні дослідження, демекологічні дослідження, вивчення паразитів на рівні виду, синекологічні (біоценотичні) дослідження і экосистемные дослідження. Аутэкологические дослідження являють собою вивчення паразитів на рівні організму. Воно включає виявлення впливів на паразита окремих факторів середовища і його реакцій на них.
Демекологичні дослідження включають вивчення паразитів на рівні популяції. Ці дослідження стосуються виявлення всіх паразитів даного виду, зв'язаних з одним хазяїном: у найпростішому випадку в одного екземпляра — хазяїна, у більш складному — у декількох екземплярів спільно живуть хазяїнів, тобто вивчення «популяції в популяції». Паразити популяції вивчаються з погляду їхньої чисельності і взаємини між собою (внутрипопуляційних відносин), що сприяють чи посиленню ослабленню штаму паразита.
Вивчення паразитів на рівні виду (ейдопаразитичний рівень) спрямовано на виявлення екології визначеного виду паразита в цілому, а саме: його життєвого циклу, зміни хазяїнів, особливостей вимог виду до умов життя на різних стадіях індивідуального розвитку, наявності популяції, внутрішньовидових (і міжпопуляційних) відносин.
Синекологічні (біоценотичні) дослідження включають вивчення біоценозу паразитів (паразитоценозу), тобто всього складу паразитів різних видів в одного хазяїна: у найпростішому випадку — в одного екземпляра хазяїна, у більш складному — у декількох екземплярів спільно живуть чи хазяїнів у виду взагалі. В останньому випадку вивчається паразитоценоз виду хазяїна. Синекологічні дослідження охоплюють вивчення складу видів паразитів і міжвидових відносин паразитів на різних стадіях індивідуального розвитку.
Екосистемні дослідження охоплюють вивчення взаємин паразитоценоза з хазяїном. Вивчаються різноманітні відносини в різних системах «паразит — хазяїн»: їхні взаємини й адаптація друг до друга, коеволюційні взаємини, міжвидові відносини усього паразитоценоза з видовою популяцією хазяїна. Екосистемні дослідження відносяться до категорії функціональної паразитології. В даний час велика увага приділяється питанням охорони навколишнього середовища і раціонального використання природних ресурсів, зв'язаних з діяльністю радянських людей у межах біосфери. У зв'язку з цим проблеми вивчення і раціонального освоєння біосфери здобувають важливе значення. З проблем біосфери й екології в даний час існує велика література. Однак з питань екології окремої групи тваринного світу — кровососних комах і кліщів — робіт мало, і вони найчастіше розрізнені. Основною екологічною особливістю даної групи членистоногих є існування на визначених етапах свого життєвого циклу стадії паразитизму. Це накладає своєрідний відбиток на екологію самих членистоногих і та шкода, що вони можуть принести хазяїну, на якому паразитують. Ця особливість диктує необхідність вивчення взаємин між членистоногим паразитом і теплокровним хазяїном, розмаїтість яких обумовлюється впливом комплексу біотичних і абіотичних компонентів біосфери.
Членистоногі являють собою найбільш різноманітні класи тваринного світу. Лише комахи складають понад 30 загонів і 1000 сімейств, десятки тисяч пологів і більш мільйона видів. Вони мають надзвичайно широке поширення в самих різних ландшафтах земної кулі. Значне місце серед них займають комахи і кліщі, багато хто з який, будучи паразитами тварин і людини, обумовлюють передачу збудників хвороб, а іноді і зберігають у своєму тілі цих збудників протягом всього і життя навіть передають потомству.
Саме вони служать основним об'єктом вивчення в системі медико-біологічних наук. Особливостям екології кровососних членистоногих, що приносять шкоду людині, присвячена дана книга.
1 Іксодові кліщі
Іксодові кліщі (сем. Ixodidae) являють собою високоспеціалізованних кровососних членистоногих, що нараховують близько 1000 видів, що відносяться до двох підродин: Ixodinae (рід Ixodes)
і Amblyomminae (пологи (Haemaphysails, Boophilus, Rhipicephalus, Dermacentor, Hyalomma, Amblyomma
і ін.). У СРСР зустрічається близько 100 видів. Поряд із кровососущими двокрилими кліщі сімейства Ixodidae є специфічними переносниками великої групи збудників трансмісивних хвороб людини і тварин, включаючи віруси, рикетсії, бактерії й ін., що обумовлено своєрідністю морфологічних, фізіологічних і біохімічних особливостей цієї групи членистоногих, значно відрізняються від кровососних комах. Вони передають збудників кліщового весняно-літнього енцефаліту, кліщових риккетсиозов, геморрагических лихоманок, туляремії й інших захворювань.
Переважна більшість захворювань, збудники яких передаються иксодовыми кліщами, відноситься до групи природно-осередкових трансмісивних хвороб, що мають поширення як у нас у країні, так і за рубежем. Иксодовые кліщі є облигатными тимчасовими паразитами хребетних тварин і відрізняються складним циклом розвитку. Вони проходять стадії яйця, личинки, німфи, імаго. Як правило, кліщі харчуються однократно в кожній активній фазі. Загальна тривалість і фенологические особливості життєвих циклів иксодовых кліщів відрізняються великою розмаїтістю й у значній мірі залежать від особливостей ареалу, територіальної приуроченості усередині нього, типів місцеперебувань, а також від особливостей зв'язків цих паразитів з хазяїнами. Деякі види іксодових кліщів є пасовищними паразитами, що підстерігають, інші - характеризуються гнездово-норовим типом паразитизму. Окремі види кліщів знаходяться на тілі теплокровного хазяїна велику частину життєвого циклу. Наприклад, у Boophilus
залишають тіло хазяїна лише насмоктавшись самки і німфи останнього віку, і єдиною активною фазою розвитку поза хазяїном є голодні личинки.
З харчуванням кліщів кров'ю на лічиквї, нимфальної і імагінальної фазах зв'язане закономірне чергування періодів «вільного» і паразитичного існування, зміна хазяїнів і значне ускладнення життєвих циклів. По числу змінюваних хазяїнів і місцеві линек життєві цикли кліщів підрозділяються на три групи: трьох-, дву- і однохозяйні (мал. 1.1).
Треххозяйний тип розвитку характерен усім Ixodinae і значної частини Ambluomminae.
Кліщі знаходяться на хазяїні тільки під час харчування. При гнездово-норовом паразитизмі можливості зустрічі з хазяїнами обмежені простором чи нори гнізда, і кліщі здебільшого харчуються на усіх фазах розвитку на особях одного виду. Прикладами можуть служити Ixodes crenulatus, що
паразитує на бабаках у Тянь-Шані, чи Rhipicephalus schulzei,
зв'язаний з різними видами степових гризунів і дрібних хижаків.
У пасовищних трьоххазяйних видів личинки і німфи звичайно харчуються на дрібних ссавцях, чи птахах, рідше, на рептиліях, а статевозрілі особи — на більш великих тваринах. Класичним прикладом подібного циклу служить розвиток Ixodes ricinus
і Iх. persulcatus, що
харчуються в статевонезрілих фазах на дрібних гризунах, комахоїдних і птахах, тоді як дорослі кліщі особливо часто зустрічаються на худобі, диких копитних, зайцях і їжаках. Подібне чергування хазяїнів у мешканця пустельних стацій Hyalomma asiaticum,
a також у присвячених переважно до степових і лугових стацій видів роду Dermacentor,
де хазяїнами личинок і німф служать гризуни, а статевозрілих форм — худоба і дикі копитні.
При двухазяйним циклі личинка, закінчивши кровосмоктання, залишається прикріпленої до хазяїна, линяє тут же на німфу, і кліщ відпадає по закінченні нимфального харчування. Двухазяйний розвиток найчастіше спостерігається в паразитів копитних тварин (Rhipicephalus bursa, Hyalomma detritum, Ну. anatolicum),
причому як статевонезрілі фази, так і дорослі кліщі паразитують на одному чи близьких видах. Лише в двухазяйного кліща Ну. plumbeum
личинки і німфи харчуються на птахах і гризунах, а імаго — на копитних.
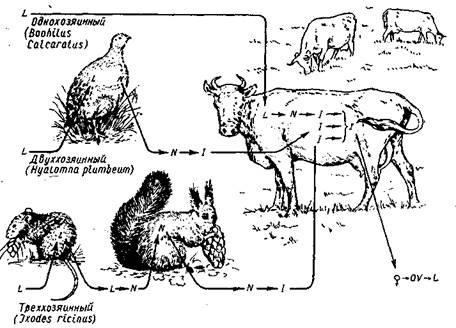
Рис. 1.1 Схема типів паразитизму в іксодових кліщів: I — дорослі кльоші; OV —
яйця; L —
личинки; N —
німфи
Найбільш пристосовані до паразитування на бродячих тваринах кліщі з однохазяйнним циклом розвитку, коли на тілі хазяїна відбуваються обидві линьки і відпадає самка, що наситилася. Подібний тип розвитку характерен усім видам роду Boophylus
і деяким видам з роду Hyalomma (Ну. scupense).
Однак в останнього виду велике число німф по насиченні відпадає з хазяїна і нормально линяє на імаго, так що в Ну. scupense
частини особей може бути характерен і двухазяйний тип розвитку. Слід зазначити, що типи розвитку в деяких видів іксодових кліщів змінюються під впливом навколишніх умов.
Запліднення самок відбувається частіше на хазяїні і, як виключення, у Зовнішнім середовищі. Самка, що наситилася кров'ю, залишає свого хазяїна і, деякий проміжок часу (4—60 днів), відкладає запліднені яйця. До цього часу самки заповзають під залишки рослинності, у тріщини ґрунту, у рослинну підстилку і т.п. Тривалість яйцекладки може затягуватися до 1—2 мес і більш. Тривалість дозрівання яєць і відкладання їх залежать від температури і вологості навколишнього середовища. Найбільш сприятливої є відносна вологість близько 100%. Одна самка відкладає від 200 до 17 тис. яєць (у тропічних видів — до 30тис.), що визначається поруч причин, у тому числі вгодованістю самки. Недостатньо наситилися самки можуть не відкладати яєць зовсім. Самці після спарювання і самки після відкладання яєць живуть нетривалий час (від декількох днів до місяця) і гинуть.
Розвиток яєць продовжується від 2 до 10 нед і більш, що визначається як видовими особливостями, так і температурою і вологістю. Особливо чуттєві яйця до коливань вологості. Так, наприклад, у кліщів lx. persulcatus
розвиток личинок і вихід їх з яєць припиняються при зниженні вологості повітря до 90%, а при вологості 65% вони гинуть. Занурення у воду не припиняє розвиток яєць (хоча воно сповільнюється), і після витягу з води вони розвиваються звичайно. Терміни розвитку личинок і німф також залежать від температури і вологості повітря і коливаються від 1 до 4 нед і більш. Для переходу в наступну фазу розвитку личинкам і німфам необхідно кровосмктання. Личинки і німфи, як і яйця, добре переносять затоплення і до 1 мес можуть знаходитися під водою.
У природі час розвитку кліщів різних видів неоднаково, тому що в цикл розвитку їх включаються сезонні діапаузи, що виявляються на різних фазах розвитку, що визначається екологічними умовами, у яких знаходяться кліщі. У залежності від тривалості розвитку і кількості діапауз розвиток іксодових кліщів може здійснюватися по однорічному, двурічному і трирічному циклам, а в деяких видів — і по чотирирічному.
При однорічному циклі розвитку кліщі на усіх фазах розвитку паразитують у теплий період одного року, діапауза буває в дорослих кліщів (чи німф). Такий цикл розвитку спостерігається звичайно в південних видів кліщів. Дворічний цикл характеризується тим, що в перший рік паразитують личинки і німфи, у другий — імаго, діапауза буває в німф і імаго (кліщ D. andersoni).
При трирічному циклі щороку має місце паразитизм кліщів тільки в одній рухливій фазі розвитку, після чого в них спостерігається діапауза (Ix. ricinus, їх. persulcatus).
На півночі в Ix. persulcatus
розвиток може затягуватися до 4 років, і, навпаки, на півдні в деяких видів сезонні явища в циклі розвитку можуть і небути, і генерація за генерацією випливає поза зв'язком із сезонами року (Boophylus calcaratus).
Тривалість життя імаго до 1—2 років, це ж і термін можливого голодування, але імаго деяких видів живуть і довше, наприклад D. pictus
— до 2—4 років, D. marginatus
— до 3 років. Личинки і німфи живуть і голодують від декількох місяців до 2 років.
Як уже згадувалися, іксодові кліщі є паразитами тимчасовими, і, як правило, імаго знаходяться на тілі своїх хребетних хазяїнів тільки в період кровосмоктання. Ссуть кров самки і самці, а також личинки і німфи (самці деяких видів, наприклад Iх. trianguliceps,
не харчуються зовсім). Іксодіди частіше підстерігають свій видобуток не в притулках (хоча є і норові види — паразити гризунів), а у відкритій природі, у траві і чагарниках. Вони займають звичайно характерну активну позу на кінцях листів, стеблинок, витягнувши і розставивши першу пару ніг з чуттєвим органом Галлера на лапках, готові моментально причепитися до минаючого мимо і тварині, що зачепила рослину, чи людині.
Кліщі можуть і самі виповзати з ґрунту, якщо поблизу їх з'являється об'єкт для кровосмоктання. Наприклад, голодні активні кліщі Iх. persulcatus
приблизно з відстані 1 м рухаються убік людини і наповзають на нього, виявлення людини відбувається, мабуть, за допомогою нюху. Більшість видів іксодових кліщів, малорухомих паразитів, що підстерігають, має широке коло хазяїнів різних видів (ссавців, птахів, рептилій), тобто їм властива поліфагія, і тільки деякі норові форми харчуються на хазяїні одного виду (наприклад, кліщ Ix. apronophorus
на водяних полівках). Багато видів кліщів дуже охоче нападають на людей.
Знання типу харчування і кола хазяїнів того чи іншого виду кліщів має велике епідеміологічне значення, тому що у випадку передачі збудників якого-небудь захворювання кліщами визначається коло можливих джерел і резервуарів збудників цього захворювання серед тварин, визначаються шляхи можливостей циркуляції збудника в чи природі серед домашніх тварин. Чекання хазяїна-прогодовувача кліщами в природних умовах може бути іноді дуже тривалим і зв'язане з поруч випадків. Тому в процесі еволюції в іксодових кліщів виробився ряд особливостей, що забезпечують їхнє існування і розвиток. Вони, як уже говорилося, здатне харчуватися кров'ю різних тварин і в кожній фазі розвитку харчуються тільки один раз, але ссуть кров порівняно довго, причому одночасно поглинається велика кількість крові; кліщі здатні довгостроково голодувати.
Тривалість харчування самок 6—12 сут, німф — 3—8 і личинок — 2—5 сут. Кліщі фіксуються на шкірі тварини за допомогою гипостома, що вводиться в ранку, прорізану хелицерами. Процес фіксації безболісний, тому що при проколі шкіри в ранку зі слиною попадають анестезуючі речовини.
Покриви тіла кліщів сильно розтяжні, до кінця кровосмоктання лінійні розміри їх збільшуються в 10 разів і більш, обсяг — майже в 300 разів, форма тіла стає яйцеподібної, колір свинцево-сірим. Під час кровосмоктання кліщ не тільки розтягується, але і розвивається, деякі клітки його тіла швидко розмножуються, тканини ростуть. Ця обставина має також визначене епідеміологічне значення, оскільки в заражених збудниками захворювань кліщах (наприклад, збудниками туляремії) у процесі кровосмоктання відбувається розмноження і збудника, і чим довше буде продовжуватися харчування, тим усі в більш зростаючих кількостях збудники будуть надходити в кров хазяїна. Тому в першої годинник після фіксації іксодові е кліщі як переносники менш небезпечні (збудників мало), і своєчасне раннє видалення зі шкіри навіть заражених кліщів нерідко може запобігти захворювання (якщо і вводиться, те мала доза збудника).
Звідси важливою мірою профілактики захворювань, переданих іксодовими кліщами, є періодичні само- і взаімоогляди в місцях поширення іксодід для раннього їхнього виявлення і видалення з тіла. Як відомо, аргасові кліщі заражають спірохетозом у першу хвилину кровосмоктання. Однократне харчування кліщів" на кожній фазі розвитку є чинником, що порівняно обмежує поширення збудників інфекційних захворювань. Заражається збудником кліщ на одній фазі, а здатної передати збудників стає тільки наступна фаза, за умови наявності трансфазовий чи трансоваріальної передачі збудника. Наприклад, вірусом кліщового енцефаліту заражається німфа кліща, харчуючи на бурундуках, а людині збудник може бути переданий тільки імаго.
Активні і харчуються дорослі форми частіше у весняно-літній період, у деяких видів спостерігається активність навесні і, у трохи меншому ступені, восени. Харчуються кліщі вдень і вночі. Личинки і німфи активні і харчуються частіше влітку.
Самостійні активні міграції в іксодових кліщів практично відсутні. Розсіювання їх у природі зв'язано з харчуванням і здійснюється тваринами-хазяїнами, яких кліщі залишають у різний час у різних місцях.
Іксодіди зустрічаються на різних територіях від північної границі лісової зони до екватора, причому при просуванні до півдня розмаїтість видів стає усе великим. Окремим видам властива приуроченість до визначених ландшафтів і місць життя. Слід зазначити, що не виключається можливість замету деякими тваринами, особливо птахами, у ті чи інші ландшафти не властивих їм видів кліщів. Іксодові кліщі заселяють різні біотопи в природі: рослинну підстилку, траву, чагарники, тріщини ґрунту, поверхневі шари її, ущелини під каменями; норові форми є частіше постійними мешканцями нір чи гризунів інших тварин. Активні голодні кліщі знаходяться на траві, чагарниках у позі чекання.
За деякими виключеннями, іксодові кліщі в населених пунктах не живуть і можуть бути тільки випадково занесені туди домашніми тваринами. Кліщ Rhipicephalus sanguineus
в усі рухливі фази розвитку паразитує на собаках, є мешканцем населених пунктів, живе в тріщинах стін надвірних будівель, у тріщинах ґрунту, у смітті на території садиби, але цей кліщ зустрічається в природі, наприклад у притулках дикобразів.
Найважливішою особливістю іксодід є нерівномірність, мозаїчність розподілу їхній по площі того чи іншого ландшафту, що визначається сукупністю ряду причин, з яких найбільше значення має фактор харчування, тобто наявність хазяїв-прогодовувачів на тій чи іншій території. Крім цього мають значення характер рослинності, особливості підстилки ґрунту, ступінь її зволоженості, вплив діяльності людини (оранка і т.п.). Але за інших рівних умов основним фактором, що визначає поширеність іксодових кліщів, є наявність тварин-прогодовувачів. Для імаго іксодід відкритих просторів у ролі таких прогодовувачів виступають дикі і доашні копитні і, у меншому ступені, хижаки і птахи. Тому в природі іксодові кліщі знаходяться не усюди, а в основному в місцях, зв'язаних з місцем життя тварин — прогодовувачі імаго (прогодовувачі личинок і німф поширені більш широко і практично завжди є там, де є прогодовувачі імаго). Такими стаціями в необжитих, мало відвідуваних людиною територіях є місця годівлі, відпочинку, водопою диких копитних — лосів, оленів, козуль, кіз, джейранів і т.п., а також тропи (у тайзі, лісі), по яких ці тварини пересуваються. Уздовж троп іксодових кліщів іноді збирається в 10—14 разів більше, ніж на відстані 5—10 м від них у глибині тайги. На освоєних людиною територіях це будуть пасовища домашньої тварин (великої і дрібної рогатої худоби, верблюдів, коней і т.п.).
По висоті розподіл нападаючих кліщів наступне: при обліку чисельності кліщів екраном у тайзі до 58% особей зустрічаються на висоті 25 див, на висоті від 25 до 75 див — близько 30% кліщів й 12%-на висоті 75—100 див. Таким чином, основна маса кліщів lx. persulcatus
у тайзі нападає з висоти близько 0,25—0,5 м.
Знання місць життя тих чи інших видів кліщів — переносників збудників заразних хвороб має велике епідеміологічне значення, тому що по характері ландшафту можна припустити наявність тих чи інших видів іксодових кліщів, а це визначає характер профілактичних і протиепідемічних заходів, місце їхнього проведення і т.п., а головне, дозволяє провести їхній попередньо.
На території СРСР у зоні тундри іксодових кліщів практично ні, що визначається холодним кліматом з коротким летом, зоною вічної мерзлоти, убогим рослинним покривом. Тільки на північно-заході Заполярья (у Карлов), де під впливом Гольфстріму клімат тепліше, зустрічається паразитуючий на птахах у їхніх гніздах кліщ Ixodes putus.
У лісовій зоні іксодових кліщі зустрічаються від берегів Балтики до Тихого океану. На заході північна границя поширення іксодід проходить по 65° с. ш., на Далекому Сході — знижується до 53° с. ш., що залежить від більш низької середньої температури на сході зони. Тайгові широколистяні ліси заселені переважно видами роду Ixodes.
Типовий для тайги кліщ Ix. persulcatus
розповсюджений від Камчатки, Курильськ островів, Сахаліну до західних границь СРСР. Найбільш численний у необжитій тайзі на вирубках, просіках, узліссях, по узбіччях звіриних стежок, у місцях відпочинку, годівлі, водопою великих ссавців. Стійкі вогнища масового розмноження кліщів виникають на місцях, де тайга навколо населених пунктів вирубана і територія використовується під пасовища. Звичайно на цій території виростають дрібнолисті породи і чагарник. На заході зони кліщі Ix. persulcatus
живуть тільки в північній її частині: райони Прибалтики, Ленінградська і Калінінградська області. Тут у європейських змішаних лісах домінує кліщ lx. ricinus,
що розповсюджений на сході до Волги, у лісових і чагарникових формаціях.
У лісовій зоні на відкритих просторах широколистяних лісів розповсюджений кліщ D. pictus
(представник степової групи кліщів), вогнища масового розмноження його знаходяться на лісових лугах, узліссях, лужках і т.п., використовуваних під пасовища; зустрічаються від західних границь СРСР на схід до Західного Сибіру включно. Для широколистяних лісів Далекого Сходу характерні види роду Haemaphysalts (Н. concinna, Н. japonica).
Кліщі Н. concinna
часто зустрічаються й у широколистяних лісах Західної Європи, Алтаю, Кузбасу, Саянів, Закавказзя. Ділянки лугів, відкриті простори і чагарникові зарості далекосхідних широколистяних лісів заселені кліщами D. silvarum.
Для лісостепової зони характерне змішання фауни лісових і степових видів кліщів. У європейських острівних лісах найбільш поширені lx. ricinus, D. pictus
(тайговий тип Ix. persulcatus
відсутній), у степових ділянках — D. marginatus, у
Західно-Сибірському лісостепу — D. pictus
і D. marginatus.
В Сибіру (до Новосибірської області) і Забайкалье лісові і чагарникові ділянки, березові колки заселяють D. silvarum,
у степах Східного Сибіру і Забайкалья — типовий для монгольських степів D. nuttali.
Фауна кліщів степової зони вже більш багата видами. Типовими представниками є кліщі роду Dermacentor
— D. marginatus, D. nuttali.
В острівних лісах, залісених балках, чагарникових заростях зустрічаються лісові кліщі D. pictus
і lx. ricinus,
а також напівпустельні Rh. rossicus, Ну. scupense.
У південній частині зони на лугах, покладах живуть Hyalomma plum-beum,
восени кліщі збираються під стогами і копицями сіна, тут же, але в меншій кількості знаходяться Haemaphysails punctata.
У південно-західній частині зони на великій рогатій худобі паразитують у масі кліщі Boophylus calcaratus
(особливо в заплавах рік). На Чорноморському узбережжі в населених пунктах живе Rh. sanguineus.
З норових форм найбільш типові Ix. laguri laguri
у норах хом'яків, lx. crenulatus
у норах хижаків і гризунів. У степовій зоні в масовій кількості кліщі розмножуються тільки на пасовищах домашніх тварин; дикі копитні через їхню нечисленність грають тут малу роль.
Для напівпустелі характерні кліщі роду Rhipicephalus (Rh. rossicus, Rh. schulzei, Rh. turanicus);
сюди заходять і степові кліщі D. marginatus.
У напівпустелях кліщі поширені в основному в норах чи гризунів у заплавах рік. Деякі норові види (Rh. schulzei)
виповзають з нір і нападають на людей і великих тварин.
Пустельна зона найбільш багата видами іксодових кліщів (більш 20 видів), що, мабуть, зв'язано зі сприятливими кліматичними умовами, розмаїтістю біотопів, достатком тварин-хазяїнів. Для цієї зони характерні кліщі роду Hyalomma,
найбільш розповсюджений вид — Н. asiaticum asiaticum.
Личинки і німфи цих кліщів паразитують на піщанках і ховрашках, а дорослі — на дрібній рогатій худобі, верблюдах. У пустельному ландшафті кліщів небагато, здебільшого вони заселяють зрошувані землі, оазиси, заплави, пустельні передгір'я. Пустельні кліщі роду Hyalomma
нападають на тварин і людей у сутінковий чи нічний час; удень від прямих сонячних променів вони ховаються в норах, тріщинах землі й в інших притулках.
Таким чином, вивчення розподілу іксодових кліщів по території СРСР показує, що кількість видів при просуванні з півночі на південь збільшується. Багато видів кліщів властиві тільки визначеним рослинним формаціям, наприклад Ix. persulcatus
— тайговим лісам, Ix. ricinus
— листяним лісам, D. pictus
— лугам і т.п. Норові форми частіше зустрічаються в різних ландшафтах, але зате зв'язані з визначеним колом хазяїнів, наприклад кліщі Іх. apronophorus
— паразити водяних полівок — поширені в різних ландшафтно-кліматичних зонах по берегах дрібних водойм, lx. crenulatus
— у норах багатьох ссавців.
Варто помітити, що хоча різні види іксодід присвячені до визначених зон, але, як правило, у тих чи інших кількостях з різною частотою вони зустрічаються й у сусідніх зонах, відмінних по ландшафті. Нижче приводиться екологічна характеристика найважливіших іксодових кліщів, що мають епідеміологічне значення.
2
Роди кліщів Ixodes
2.1 Рід
Ixsodes
Ixodes persulcatus
живе в південній частині тайгової смуги — від Камчатки до Карелії, зрідка виявляється і південніше — у Московської, Брянської, Орловський областях, зустрічається на Алтаї (мал. 2.1). Характерні стації перебування — хвойні і листяні ліси; найбільш рясний у змішаних хвойно-листяних лісах. У тайзі кліщі збираються (до середини травня) уздовж стежок у місцях відпочинку і водопою диких копитних, в освоєній тайзі — на вирубках, використовуваних під пасовища, буреломах — місцях перебування великих хижаків, у чагарниках, кілках.
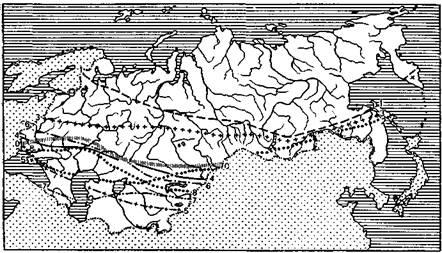
Рисунок 2.1 - Карта поширення кліщів Ixodoidea у
СРСР. Північна границя пологів:
/ — Izodes, 2
— Haemaphysalis, 3 — Dermacentor, 4
— Rhipicephalus, 5
— Boophilus,
6
— Hyalomma, 7
— Ornithodoros, 8 — Argas
Ix. persulcatus
— треххазяйнний кліщ. Дорослі його форми паразитують на великих домашніх тварин (в освоєних зонах тайги — на лосях, оленях і т.п.), личинки і німфи — на дрібних ссавцях (гризунах, комахоїдних хижаках), птахах, що ведуть наземний спосіб життя (наприклад, на рябчиках). На людину нападають як імаго, так іноді і личинки. Найбільша активність дорослих, німф і личинок спостерігається в першій половині теплого періоду року, імаго — із квітня, личинки і німфи — із травня.
Епідеміологічне значення Ix. persulcatus
полягає в тому, що він є переносником збудників кліщового енцефаліту і туляремії.
Ixodes ricinus
(мал. 2.2) розповсюджений у європейській частині СРСР (на схід приблизно до Волги), на Кавказу, у Західній Європі, Англії, Північній Африці. Найбільш характерні стації — дрібнолисті і хвойно-листяні ліси і відкриті простори з чагарниковими заростями, на півночі — сухі, а на півдні — вологі місця. Вогнища масового розмноження знаходяться в місцях випасу домашніх тварин і в місцях, де частіше тримаються великі дикі тварини (лосі), на вирубках, узліссях і т.п.

Рисунок 2.2 Ixodes ricinus
(самка)

Рисунок 2.3 - Схема сезонного ходу активності імаго Ixodes ricinus
у різних кліматичних умовах (по Л.В. Бабенко):
1
— район з теплою вологою зимою і сухому влітку; 2
— район із прохолодною зимою і рідкими морозними днями; 3
— район з континентальним кліматом середньої смуги Європи; 4
— се° вірні райони з коротким прохолодним летом
lx. ricinus
— треххазяинний кліщ. Для імаго хазяїнами служать великі ссавці, особливо домашні тварини, хижаки (лисиці, барсуки), олені, лосі, а також їжаки, зайці і птахи. Личинки і німфи паразитують переважно на дрібних наземних хребетних, у першу чергу на їжаках, а на півдні — також на зміях і ящірках. Нападають на людину дорослі і німфи, але іноді і личинки. На території Середнього Наддніпрянщин Ix. ricinus
відноситься до числа домінантних видів. Масове розмноження кліщів відзначається в листяних і змішаних лісах, у лісових болотах, однак у лісових біотопах навіть в одній і тій же місцевості lx, ricinus
розподілені нерівномірно. Найчастіше вони зосереджені на узбіччях і лісових дорогах, у заболочених місцях — на ділянках з помірною вологістю і добре дренірованній ґрунтом. Цей вид кліщів має широке коло хазяїнів (він виявлений на 45 видах ссавців). Основними хазяїнами статевозрілих кліщів служать домашні тварини, а також плямистий олень, лось, лисиця, козуля. Статевонезрілі фрази прогодовуються переважно на гризунах (рудий, звичайної і водяник полівках, лісовий і жолтогорлой мишах) і землерийках (звичайної і малий бурозубках).
lx. ricinus
зустрічається на савец в усі сезони року, однак зміни сезонної активності окремих фаз розвитку досить своєрідні. Активність дорослих кліщів починається в Середнього Наддніпрянщин з березня, масове паразитування на тварин збігається тут з початком випасу худоби (квітень), і число паразитуючих особей наростає до середини травня (до 250 кліщів на окремих тваринах). Велика чисельність і щільність Ix. ricinus
на даній території, як показали дослідження Н.Т. Никитченко (1985), порозуміваються достатньою вологістю, оптимальною температурою, сприятливим рослинним покривом, підстилкою і щільністю заселення хазяїнами кліщів. Інтенсивність поразки тварин трохи знижується в червні-серпні. У середині вересня спостерігається другий пік активності. Максимум кліщів на окремих тваринах у цей період досягає 65 особей.
Німфи на тварин з'являються в I декаді квітня. Максимум їхнього паразитування приходиться на червень. У липні число паразитуючих особей різко знижується. Потім відбувається підвищення у вересні з наступним зниженням до листопада. Крива сезонної активності німф даного виду кліщів має дві різко виражені вершини з максимумом у травні-червні і восени (вересень).
Личинки Ix. ricinus
протягом доби розповзаються від місця вилуплення, як показали дослідження Н.Н. Лебедєвої й А.В. Фильчагова (1985), на відстань 0,5—2,5 м. Однак велика їхня частина звичайно концентрується в радіусі до 1,5 м (60.—80%). Площа ділянки, зайнятого личинками з однієї кладки, складає в різні дні від 1 до 5,5 м2
, а сумарно за кілька тижнів — 7,5—8,5 мг. У пересуванні личинок не відзначається якої-небудь цілеспрямованості, вони розповзаються рівномірно в усі сторони. Голодні личинки в більшості випадків зустрічаються групами, у яких одночасно можуть знаходитися до 1000 і більш активних голодних особей (Левин, 1985). Активність личинок починається з квітня, максимум спостерігається в травні; у червні-липні вона знижується, а в серпні знову відзначається її підйом. Після другого піка активності починається повільний її спад;до кінця грудня. Період активності лічинкової стадії на тварин набагато триваліше (квітень-грудень), чим інших фаз розвитку.
Зимують Ix. ricinus
на усіх фазах розвитку. Статевозрілі голодні кліщі на зиму ховаються під лісову підстилку, суху траву, нори тварин.
У помірному поясі паразитування дорослих особей спостерігається в теплий період року з
двома максимумами: більш високим навесні (квітень— травень) і восени (серпень — вересень) (мал. 46). У середині літа відбуваються різке зниження активності дорослих і наростання активності ювенальних фаз, що паразитують на дрібних хребетним. У теплих районах (Північна Африка, Південна Європа, Закавказзя) сезон паразитування дорослих — осінньо-зимньо-весняний період; улітку знахідки кліщів на хазяях дуже рідкі.
Кліщ є переносником збудників кліщового енцефаліту, туляремії й інших захворювань, а також кровопаразитів — збудників хвороб домашніх тварин.
2.2 Рід Dermacentor
Dermacentor pictus
(мал. 2.4) розповсюджений у Середній Європі, Англії, на півдні смуги європейських широколистяних лісів і в лісостеп, у Західному Сибіру, у горах Криму, Кавказу, Закавказзя, Казахстану, Киргизії. Заселяє лісові лучки, пасовища, заливні луги, прикордонні степові ділянки, D. pictus
— трьоххазяйний кліщ. Дорослі великі кліщі паразитують на вівцях, коровах; молоді — на дрібних ссавцях, частіше на полівках; на їжаках і зайцях — усі форми. На людину нападає рідко. Активні дорослі кліщі навесні (у масі) і восени (5% весняної активності), молоді — улітку. У Закавказзя їхнє паразитування продовжується до зими (мал. 2.4).
Характерним для кліщів Dermacentor
є їхня різна кількість на різних пасовищах. Строкатість розподілу кліщів на пасовищах (а не на території взагалі) залежить від розміщення на них полівок — хазяїнів молодих форм кліщів. Весняна перевага статевозрілих кліщів на якій-небудь ділянці відповідає літньому поширенню популяції звичайної полівки за попередні роки. Основними біотопами служать купинясті луги, вирубки, узлісся, тобто ті місця, де полівки можуть укритися від хижаків. Крім цього кліщів звичайно більше там, де худоба знаходиться довше (місця «полуденка»). Менше кліщів спостерігається в місцях з витоптаною рослинністю (місця частого прогону худоби). Зазначену залежність кількості кліщів від кількості гризунів можна використовувати для прогнозу чисельності кліщів. Після піка чисельності гризунів у наступні роки, особливо в другий і третій (але не перший), пасовищах можна екати масової появи кліщів. Величезні запаси статевозрілих кліщів, що виникають у природі в рік масового розмноження мишеподібних гризунів, витрачається потім протягом наступних 3—4 років.

Рисунок 2.4 - Dermacentor pictus
(самець)
Епідеміологічне значення D. pictus
полягає в переносі збудників туляремії, омської геморрагічної лихоманки, кліщового сипного тифу.
Dermacentor silvarum
розповсюджений на Далекому Сході, у Примор'ї, у Забайкалье, на Алтаї до Західного Сибіру, у Північній Монголії. Типовими ландшафтами для цього кліща є лісостеп, ліси, але головним чином ділянки, піддані освоєнню людиною, покриті чагарниковою рослинністю вторинного типу. D. silvarum
— трьоххаозяйнний кліщ, дорослі паразитують на великих ссавцях — домашніх і диких, можлива зимівля на худобі; молоді форми — на дрібних ссавцях. Це перший весняний кліщ, максимум нападу — наприкінці травня, друга, менш висока хвиля активності — восени. Вид має велике епідеміологічне значення, тому що є переносником збудників кліщового енцефаліту і кліщового сипного тифу на Далекому Сході, а також кровопаразитів тварин (піроплазмоз, нуталліоз).
Dermacentor marginatus
зустрічається на півдні європейської частини СРСР, Закавказзя, Казахстану, у горах Середньої Азії, на півдні Західного Сибіру, у Південній Європі, у рівнинних і гірських степах, у лісостепу, у гірських лісах (на пасовищах), балках, заплавних лугах, лісосмугах — у місцях, де живуть можливі хазяїни. Характерно, що кльоші D. marginatus
відсутні в населених пунктах. Дорослі особи годуються на великих домашніх і диких ссавцях (вовки, зайці, їжаки), можуть нападати на людину; молоді форми паразитують на дрібних ссавцях — гризунах, комахоїдних. Трьоххазяйнний вид, сезон паразиторування — весна (з лютого по травень на півдні), почасти осінь, для молодих форм — літо. Епідеміологічне значення виду в тім, що кльоші є переносниками і хоронителями збудників туляремії, кліщового сипного тифу, кровепаразитарних захворювань тварин.
Dermacentor nuttali
розповсюджений у степах Забайкалья, Східної «Сибіру, у Монголії. Це трьоххазяйнний кліщ, дорослі паразитують на великих, молоді фази — на дрібних ссавцях (крім цього на зайцях, кішках, собаках); часто зимують на хазяях, нападають на людину. Дорослі кліщі активні навесні і трохи слабкіше восени, максимум — з кінця квітня до середини травня (мал. 48). Є переносниками збудників кліщового сипного тифу.
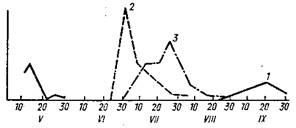
Рисунок 2.5 - Схема сезонного ходу активності різних фаз розвитку Dermacentor nuttali
(по В. А. Меринову): 1
— імаго. 2
— личинки, 3
— німфи. По вертикалі — нн-декіьі достатку; по горизонталі — дати
2.3 Рід Haemaphysalis
Haemaphysalis concinna
(мал. 2.6) особливо численний на Далекому Сході (Хабаровський і Приморський краї), на півдні європейської частини РСФСР, у Криму, Закавказзі, Західній Європі, Ірану, Південно-Східній Азії. Північніше 51° с. ш. цей вид не живе. Характерні стації: у листяних вологих і змішаних лісах, на відкритих ділянках, у вологих кочкарниках (Примор'я), у лісах Закавказзя. Вогнища масового розмноження кліщів — у місцях перебування диких копитних, оленярських парках.
Нает. concinna
— трьоххазяйнний кліщ, дорослі паразитують на великих ссавцях (копитних і хижих), активні з весни до осені, найбільш активні в червні; личинки і німфи — на дрібних ссавцях, птахах протягом усього теплого періоду, нападає на людину, але характерні погана присмоктування і менша агресивність, чим у lx. persulcatus;
можуть нападати і німфи. Кліщі даного виду є переносниками збудників кліщового сипного тифу і кліщового енцефаліту.
Haemaphysalis punctata
розповсюджений на півдні європейської частини СРСР, у Казахстану, Середній Азії, Європі, Північній Африці, Ірану. Характерні ландшафти: гірські ліси, степи і пустелі, долини рік. Розповсюджений широко, але вогнища масового розмноження не зустрічаються. Тртоххазяйнний кліщ, дорослі паразитують на великих ссавцях (великому і крейдою рогатій худобі, верблюдах, свинях), личинки і німфи на великих тваринах харчуються рідко, частіше на дрібних ссавцях, особливо птахах (курях, фазанах, галках, горобцях і т.п.), рідше — на рептиліях, нападає на людину. Сезон паразитування дорослих кліщів — весна й осінь, на півдні — цілий рік, личинок і німф — частіше влітку. Кліщі переносять збудників кліщового сипного тифу в Середній Азії.
2.4 Рід Rhipicephalus
Rhipicephalus sanguineus
(мал. 2.7) розповсюджений на всіх материках земної кулі, типові форми — у Середземномор'я; у СРСР — на Чорноморському узбережжі, у Криму, Закавказзі, Туркменії. Хазяїнами в основному служать собаки, рідше — інші домашні тварини, їжаки, хижаки, гризуни (сірі пацюки). На собаках кліщі харчуються на всіх рухливих стадіях розвитку по трьоххазяйнному типі. Часто кліщі заселяють приміщення, де є собаки (тріщини стін будівель, під штукатуркою і т.п.). У дикій природі живуть у лігвищах і норах шакала і дикобраза.
Рисунок 2.6 - Haemaphysalis concinna
(самець)
Рисунок 2.7 - Xhipicephalus sanguineus
(самець)
При 30° весь життєвий цикл продовжується близько 2 мес, але може затягуватися і до 3 років. Сезон паразитування продовжується з весни протягом усього літа (дорослі), личинки і німфи паразитують на собаках у масі влітку до жовтня (мал. 2.8). Нападає на людину. Епідеміологічне значення складається в передачі збудників марсельської лихоманки, лихоманки Ку, спірохет іспано-північноамериканського кліщового поворотного тифу, можливо, вірусу сказу (знайдені спонтанно заражені вірусом сказу кліщі Rh. sanguineus
— у Китаї, Поволжі), у США — збудників лихоманки Скелястих гір.
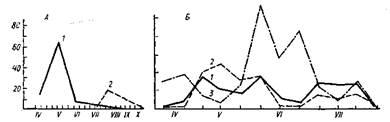
Рисунок 2.8 - Сезонний хід активності різних фаз життєвого циклу Rhipicephalus
(no
Е. А. Клюшкиной і К. И. Кондрашкиной).
А
— Rlupicephalus sanguineus
(Крим); Б
— Rh. schulzei
(Казахстан); 1
— імаго, 2 —
личинки, 3
— німфи. По вертикалі — індекси достатку, по горизонталі — місяці
Пустельні і напівпустельні — Rh. schulzei
(паразит ховрашків) і Rh. pumilio
(паразит піщанок, зайців і ін.) — зрідка можуть нападати і на великих тварин (верблюдів, велика і дрібна рогата худоба й ін.) і людини; можливо, грають деяку роль у поширенні чуми, тому що з зібраних у природі цих видів кліщів виділені збудники чуми.
Rhipicephalus pumilio
— трьоххазяйнний паразитичний кліщ, розповсюджений у рівнинних і передгірних напівпустелях Прикаспія, Казахстану, Середньої Азії, а поза СРСР — на заході Монголії і Китаю. Він часто зустрічається в тугайних заростях. Паразитування кліща на усіх фазах розвитку відзначається протягом усього теплого періоду, причому в личинок і німф спостерігається два підйоми чисельності (на початку і кінці теплого сезону), а дорослі кліщі активні звичайно в першій половині літа. Зимують усі фази. Таким чином, даний кліщ характеризується багаторічним (2—3 роки) циклом розвитку (Белозеров, 1985).
Харчування личинок і німф продовжується 3—8 днів. Самки починають відкладання яєць при 25° через 4—6 днів після насичення. Ембріональний розвиток при цій температурі продовжується 25—30 днів. Личинки активізуються через 5—8 днів після вилуплення незалежно від фотоперіодичних умов. Тривалість перетворення ситих личинок у німф при 25° складає 11 —15 днів, а при 30° — 7—10 днів. Голодні німфи виявляють високу рухливість через 5—8 днів після линяння в умовах довгого дня (20 ч світла в добу), а в умовах короткого дня (12 ч світла) зберігають пасивність не менш 4—6 мес. Світловий режим чітко позначається і на агресивності голодних личинок і німф: довгий день підсилює агресію, короткий — сприяє пасивності.
2.5 Рід Hyalomma
Hyalomma plumbeum
(мал. 2.9) розповсюджений на Україні, у Криму, на Кавказу, у Нижнім Поволжі, Середній Азії, Південній Європі, Африці, Ірану. Дорослі кліщі зустрічаються частіше на різнотравних ділянках (старі ріллі, цілина), у розрідженому листяному лісі, у западинах, балках, лісосмугах, у місцях існування їхніх хазяїнів. Відзначено, що кліщі восени збираються в копицях і стогах сіна, де і залишаються на зимівлю. Хазяїнами для дорослих особі служать головним чином сільськогосподарські тварини, але харчуються і на зайцях (русаках), дрохвах. Хазяїнами личинок і німф є в основному зайці, а також дикі (дрохва, курлатка й ін.) і домашні птахи. Тип харчування — двуххазяйний, сезон активності — весь теплий період року до листопада, зі зниженням чисельності нападу кліщів з липня (мал. 53). Передає збудників кримської геморрагичною лихоманки і лихоманки Ку.
Hyalomma anatolicum anatolicutn
розповсюджений на півдні СРСР (у Казахстану, Середній Азії, Закавказзі), в Ірану, Африці. Живе на цілинних ділянках, покладах, заплавних лугах. Паразитує на.великих ссавців, в основному домашніх тварин, джейранах. Треххозяинный вид. Сезон паразитування березень — листопад, найбільш численний у березні — квітні. Передає збудників геморрагичної лихоманки типу кримської в Середній Азії, лихоманки Ку.

Рисунок 2.9 - Hyalomma plumbeum (самка)
Hyalomma asiaticum asiaticum розповсюджений у Середній Азії, Казахстану, Закавказзі. Пустельна і напівпустельна форми зустрічаються на закріплених пісках, такырах, у норах гризунів (піщанок), у населених пунктах — у тріщинах дувалов, стін будівель. Паразитує на великих домашніх тваринах, головним чином верблюдах, великих диких ссавців, личинки — на гризунах, їжаках. Активний з березня по жовтень, найбільша чисельність у травні-червні. Передає збудників лихоманки Ку.

Рисунок 2.10 - Схема сезонного ходу заклещевленности худоби статевозрілими кліщами і граків — личинками і німфами Hyalomma plumbeum plumbeum
(пo Л. И. Зиміної із соавт.): 1
— імаго, 2
— личинки і німфи. По вертикалі — індекс достатку; по горизонталі — дати
Рисунок 2.11 - Boophilus calcaratus
(самець)
2.6 Рід Boophilus
У СРСР тільки один вид — Boophilus calcaratus (мал. 54), розповсюджений на півдні європейської частини, у Закавказзі і Середній Азії. Живе в різних ландшафтах — від вологих субтропіків до сухих степів і пустель, в останніх присвячений до заплав рік, оазисам. У масі знаходиться на пасовищах, особливо лісового, передгірного і гірського поясів. Паразит великої рогатої худоби. Активний переважно в теплий час року (до грудня), дає 2—3 генерації в рік. Розвивається по однохазяйнному типі. Личинки нездатні до тривалого голодування, що дозволяє звільняти від них пасовища шляхом тимчасового припинення випасу худоби на черговій ділянці. Має тільки ветеринарне значення як переносник піроплазмозу, фасциолеза, анаплазмозу, спірохетозу великої рогатої худоби.
2.7 Рід Атblуотта
У СРСР представники роду не виявлені. На Американському континенті деякі види мають велике епідеміологічне значення. Атblуотта cajenensis є переносником риккетсиоза — бразильської плямистої лихоманки (тифу Сан-Пауло). Спонтанно заражених кліщів знімали із собак і людей. Атblуотта americanum передають збудників лихоманки Скелястих гір, лихоманки Ку.
3
Епідеміологічне значення
Іксодові кліщі є важливими біологічними переносниками збудників багатьох хвороб людини і тварин, включаючи віруси, ріккетсії, бактерії, піроплазми, тейлерии й ін. У гемолімфі і яйцях кліщів пологів. Hyalomma і Boophilus часто зустрічається жгутиконосець Crithidia hyalommae, з їхнього організму виділено понад 15 видів грибів пологів Aspergillus, РепісіШит і ін.
У сьогодення - час у різних країнах установлена роль іксодових кліщів у передачі понад 29 арбовирусів. Епідеміологічне значення деяких з них приведено в табл. 11.
Кліщовий енцефаліт — гостре інфекційне захворювання з переважною поразкою нервової системи. Викликається вірусом групи Варбовирусів, що відноситься до дрібним Рнк-вірусам. Вірус довгостроково зберігається в організмі іксодових кліщів, у яких спостерігається його трансфазова передача. Інкубаційний період продовжується від 1 до 2 нед. Захворювання починається раптово з підвищення температури до високих цифр, а потім з'являються симптоми поразки центральної нервової системи: порушення чутливості, координованих рухів. Форма перебігу хвороби може бути різної — від безсимптомної і стертої до дуже важкими, ускладненими стійкими паралічами і парезами, а іноді вона може приводити до смерті. Захворювання звичайно з'являються в квітні — нюне, потім кількість їх різка знижується, що залежить від періоду активності кліщів.
Кліщовий енцефаліт є осередковою-осередковим-природно-осередковим трансміссивним захворюванням, розповсюдженим від узбережжя Тихого до Атлантичного океану. Установлено спонтанне вірусоносіївство чи виявлені антитіла до вірусу кліщового енцефаліту в більшості видів хребетних тварин, зв'язаних з лісовими біоценозами в межах ареалу цієї інфекції. Вірус виділений і від членистоногих різних груп. Однак не всі теплокровні і членистоногі відіграють однакову роль у циркуляції вірусу кліщового енцефаліту. Як показали дослідження Э. И. Коренберга (1977, 1985), абсолютна і відносна кількість видів тварин, що прогодовують личинок, німф і імаго кліщів Ixodes persulcatus і їх. ricinus, неоднаково в різних частинах ареалу кліщового енцефаліту і визначаються насамперед рівнем чисельності кліщів.
Таблиця 3.1 - Роль іксодових кліщів у передачі збудників деяких вірусних природно-очагових хвороб
| Захворювання |
Основні переносники |
Трансфазовая передача |
Трансовариальная передача |
Дикі хребетні — резервуари збудника |
Поширення |
| Кліщовий літній-весняно-літній енцефаліт |
fxadcs persulcatus.
їх. ridnus.
Dermacentor pictus,
D. silvarum,
H. concinna
|
+ |
+ |
багато видів ссавців ІІ
птахів
|
лісова зона європейської і
азіатської частин СРСР
|
| Кримська і середньоазіатська геморрагичні лихоманки |
Hyalnmma plumbeum, Hy anatolicurn.
Ну. dctritum,
Rh. turanicum
|
+ |
— |
гризуни, птахи |
степові райони юта європейської частини СРСР і Середньої
Азії
|
| Омська геморрагічна чихорадка |
Dermacentor pictus,
D. marginatus
|
+ |
+ |
гризуни |
лісостеп Західного Сибіру |
| Киасанурска лісна хвороба |
Haemaphysalis spinigeia,
H. tur/uris,
H, papuana
Ixodes,
Amblyomma
|
+ |
— |
мавпи і дрібні лісові ссавці |
лісові райони Індії |
Шотландський енцефаліт, чи
вертушка овець
|
Ixodca ricinus |
+ |
— |
вівці і кози |
Англія |
| Кемеровська лихоманка |
Ixodes persutcalu!s |
даних немає |
— |
--- |
південь Центрального Сибіру |
| Лихоманка Західного Нілу |
види роду llyalomma |
+ |
— |
птаха |
Африка, Близьке Схід, Середземномор'я |
| Колорадська кліщова хороброрадка |
Dermacentor andciwni |
+ |
— |
гризуни |
США |
| Лихоманка Повассан |
Dermacentcr andersoni |
+ |
— |
дрібні ссавці |
Канада, США |
| Малайська лихоманка Лангат |
Ixodca granulatits |
+ |
— |
гризуни |
джунглі Малайської підлоги-
острова
|
| Хвороба овець Найробі |
Rhipicephalus appcndiculalus |
+ |
— |
циркуляція між вівцями і козами |
Східна Африка |
Важливу роль у диссемінації вірусу грають мишовидні гризуни і землерийки. Дослідження показують, що хазяїнами предимагінальних фаз іксодових кліщів у європейський-східно-європейських южнотаєжних і широколиственно-хвойних лісах можуть бути всі дрібні і середній величини звірки, зв'язані з лісовими біотопами; індекси достатку личинок у них приблизно в 5—10 разів вище, ніж індекси достатку німф. Найвищі среднемноголетние індекси достатку личинок (1,5—3,2) властиві полівкам і більшості землерийок. Основними годувальниками личінкової фази і важливими годувальниками німф у лісах сходу Російської рівнини є руді полівки (Clethrio-nomys glareolus) і звичайні бурозубки (Sorex araneus). Ці годувальники характерні для гнітючої частини ареалу кліщового енцефаліту з незначними варіантами, обумовленими місцевими особливостями фауни і чисельності звірків. Це підтверджено численними зоолого-паразитологічними спостереженнями, проведеними в різних частинах області поширення кліщового енцефаліту.
Крім дрібних ссавців годувальниками предимагінальних фаз кліщів Ix. ricinus і їх. persulcatus і особливо німф є більш великі тварини різних систематичних груп (їжаки, білки, бурундуки, кроти). На цих звірках одночасно може паразитувати від 20 до 70 кліщів. Таким чином, дрібні ссавці є головними хазяїнами лічинкової фази основних переносників вірусу кліщового енцефаліту (Ix. ricinus і їх. persulcatus) і цим забезпечують можливість існування кліщів на визначеному етапі їхнього життєвого циклу і всієї епізоотичної ланцюга природних вогнищ цієї інфекції.
Незважаючи на річні і сезонні розходження в чисельності дрібних ссавців нерівномірність їхнього розміщення, щорічно і майже повсюдно звірків буває, як правило, цілком достатньо, щоб прокормити така кількість личинок і німф, що забезпечує подальше існування генерації головних переносників вірусу кліщового енцефаліту.
Сірологічні дослідження показують, що тварини цієї групи часто заражаються кліщовим енцефалітом. Однак вони сприяють диссемінації вірусу лише в тих випадках, коли на них паразитує велике число як заражених, так і незаражених кліщів. У природних вогнищах такі ситуації мають місце лише в порівняно невеликої кількості видів тварин (землеройки-бурозубки, лісові полівки). У цілому дрібні ссавці лише в незначних межах змінюють масштаб інфікування покоління кліщів, обумовлений ступенем вірусофорності ситих імаго і їхньою здатністю до трансовариальної передачі збудника.
Великі ссавці (олені й інші копитні) є годувальниками не тільки імаго, але і великої кількості німф Ix. persulcatus і Ix. ricinus. Кліщі паразитують також на тваринних середніх розмірів: їжаках, зайцях і ін. Таким чином, ці дикі тварини, будучи годувальниками імаго і німф іксодових кліщів, мають важливе эпізоотичне значення. Вони обумовлюють існування короткого ланцюга передачі збудника кліщами від чи імаго імаго і німф і незаражених німф при їх одночасному кровосмоктанні на диких ссавцях. Це істотно доповнює загальновідомий довгий шлях передачі вірусу — трансоваріальний і трансфазовий.
Установлено, що у великих диких і домашніх тваринних і дрібних ссавців антитіла, що з'являються в крові внаслідок зараження кліщовим енцефалітом, довгостроково не зберігаються (Коренберг і ін., 1977). Їхні титри постійно знижуються, і при відсутності повторного зараження антитіла порівняно швидко зникають. Багато тварин стають сіронегативними вже до кінця сезону активності кліщів, що випливає за появою в їхнє крові антитіл. Швидке зникнення гуморальних антитіл у диких і домашніх тварин є однієї з характерних рис епізоотології кліщового енцефаліту. Можливість багаторазової вірусемії в довгостроково живучих ссавців дає підставу думати, що саме годувальники дорослих кліщів мають важливе значення для підтримки циркуляції вірусу кліщового енцефаліту у вогнищі, тому що вони визначають рівень зараженості ситих імаго іксодових кліщів і їхнього потомства. Сірологічно було встановлено, що птаха ряду видів можуть регулярно заражатися кліщовим енцефалітом. Однак загальна щільність населення птахів, як правило, значно менше, ніж щільність населення ссавців, тому абсолютне число птахів, що можуть протягом епізоотичного сезону заразитися кліщовим енцефалітом, порівняно невелике. Тому варто вважати, що птахи є лише додатковими годувальниками личинок і німф, і не вони визначають структуру природних вогнищ у епізоотичну ситуацію. Птахи слабко сприйнятливі до збудника кліщового енцефаліту, є лише додатковими хазяїнами вірусу і можуть брати помітну участь у процесі його циркуляції тільки у вогнищах, де на них нападає багато іксодових кліщів.
Таким чином, основними критеріями, що визначають значення виду теплокровних тварин у циркуляції вірусу кліщового енцефаліту, є: чисельність виду, характер динаміки чисельності даного виду тварини й особливо рівень чисельності паразитуючих на них кліщів. У першу чергу варто назвати оленів, домашніх копитних, мишевидних гризунів (особливо полівок: руду, червону, економку), землерийок. З птахів найбільше епізоотичне значення розраховують на, що харчуються на землі і тому доступні для личинок і німф кліщів (зозуля, вівсянка, зяблик, повзик, різні види дроздів).
Вірусемія у тварин не має великої залежності від ступеня чутливості до вірусу і продовжується в середньому близько 2 нед. У період сплячки тварин тривалість вірусемії може збільшитися до 3 мес., що обумовлює тривале збереження збудника в природі.
Основний шлях передачі вірусу кліщового енцефаліту — трансмісивний, однак можливе зараження людини аліментарним шляхом при вживанні незнезараженого молока кіз і корів. Вірус кліщового енцефаліту виділений від членистоногих багатьох груп: іксодових і гамазових кліщів, бліх, комарів, але їхня роль у епізоотології й епідеміології даного захворювання далеко не однакова.
Основними переносниками і резервуарами вірусу є іксодові кліщі Ixodes persulcatus і Ix. ricinus; менше значення мають Dermacentor silvarum, D. pictus, D. marginatus, Haemaphysalis concinna, Haem. japonica, Ix. trianguliceps. Ці кліщі виявлені як носії вірусу кліщового енцефаліту в різних районах СРСР, а також у Західній і Східній Європі. Збудник виділяється і від самок і від самців.
Організм кліща є сприятливим середовищем для розмноження вірусу. У Ix. persulcatus, заражених експериментально, вірус, інтенсивно розмножуючи, досягає максимальної концентрації до 40-му дню після інфікування. За цей період кількість вірусу зростає в 1000 разів у порівнянні зі змістом його в кліщах у перші дні після зараження. До 6-му дня після годівлі вірус проникає в усі органи кліща. Найбільша кількість вірусу накопичується в кишечнику, половому апараті і слинних залозах. Останнє сприяє дисемінації вірусу серед тварин — годувальників кліщів, в організм яких вірус проникає зі слинних залоз у період кровосмоктання. Значна концентрація збудника в яєчнику створює передумови для трансоваріальної передачі кліщами вірусу потомству. Установлено здатність вірусу перезимовувати в тілі кліща. Доведена трансфазова передача вірусу в іксодових кліщів.
Кількість заражених вірусом кліщів у вогнищах може коливатися від одиничних особ до 2—5% і навіть до 40%. Вірусофорність кліщів не є постійною величиною, що залежить як від біотичних, так і від абіотичних факторів зовнішнього середовища. У природних умовах поряд з дисемінацією вірусу йде процес звільнення кліщів від вірусу. Необхідною умовою тривалого існування природних вогнищ кліщового енцефаліту є періодичне додаткове зараження кліщів від хребетних тварин, оскільки передача вірусу трансфазово і трансоваріально в іксодових кліщів нерівномірна, вірус передається не всьому потомству і не завжди утримуватися в наступних фазах розвитку, що залежить від ступеня вірусемії у тварин-донорів.
Показники зараженості кліщів не знаходяться в прямої залежності від чисельності тих чи інших видів у природі. Порівняльне вивчення зараженості кліщів Ix. persulcatus і Нает. concinna показало, що умови розмноження вірусу в організмі кліщів нерівнозначні. Вірус у Ix. persulcatus поширювався більш інтенсивно і знаходився в більшій концентрації, чим у кліщах Я. concinna. Інфікування Ix. persulcatus і їх. ricinus у західних районах СРСР, ендемічних по кліщовому енцефаліті, приблизно однакова. Звідси випливає, що різні види іксодових кліщів відіграють неоднакову роль в епідеміології кліщового енцефаліту.
На зараженість кліщів вірусом кліщового енцефаліту впливає віковий склад популяції кліщів. З'ясуванню факторів, що впливають на зараженість іксодових кліщів вірусом кліщового енцефаліту, приділяється велика увага в зв'язку з важливістю встановлення й оцінки причинних зв'язків для епідеміологічного прогнозу. Згідно літературний даної, набір факторів, що впливають на рівень зараженості кліщів, досить великий — температура, кількість опадів, число хмарних днів, віковий чи склад рівень імунітету населення дрібних ссавців і ін. А.А. Катин (1984), ґрунтуючись на експериментальних даних про тривалість збереження вірусу в діапаузуючий і недіапаузитуючий кліщах, а також на результатах польових досліджень, припустив, що зараженість кліщів вище в популяціях, що розвиваються переважно по 3-літньому циклі (без діапауз), чим у популяціях, що розвиваються по 4- і 5-літньому циклам (з однієї чи двома діапаузами).
Дослідження, проведені співробітниками Інституту медичної паразитології і тропічної медицини ім. Е.И. Марциновского Мінздраву СРСР показали, що значний перепад висот Джойского хребта і різна експозиція схилів обумовлюють істотні ландшафтні розходження в умовах існування тайгових кліщів і їх годувальників. В міру збільшення висоти місцевості підвищується вологість, але зменшується кількість тепла. У зв'язку з цим розвиток
кліщів на різних ділянках йшло по різних схемах. У нижній частині пояса подтайги на висотах близько 500 м над ур. м. популяція імаго тайгового кліща на 1/4 складається з особ, що розвиваються по 3-літньому циклі (без діапауз), половина — по 4-літньому (з однією діапаузою) і 1/4 — по 5-літньому (із двома діапаузами). Зі збільшенням висоти місцевості зменшується температура повітря і скорочується вегетаційний період. Відповідно в популяції кліщів зростає частка діапаузуючих особ. Так, у підпоясі черневої тайги по 3-літньому циклі розвивалося лише близько 3% особ, по 4-літньому — близько 1/3 і 5-літньому — 2/3. У підпоясі гірничо-тайгових лісів (1000 м) 1/5 популяції кліщів складалася з особ, що розвиваються по 4-літньому циклі, і 4/5 — по 5-літньому (Наумов, 1975; Гутова, 1985).
Середня за 13 років спостережень зараженість кліщів на цих висотах склала відповідно 0,2, 0,7 і 0%, сумарне за сезон достаток дрібних ссавців — 1,4, 10,7 і 14,8 на 100 ловушко-суток. Ці матеріали — ще одне свідчення на користь думки про поліфакторну залежність рівня зараженості кліщів.
У східних районах країни (Урал, Сибір, Далекий Схід) зараження людини відбувається в основному через кліщів Ix. persulcatus, а в західних — через Ix. ricinus. Однак в останні роки в Прибалтиці і Ленінградській області кліщі Ix. persulcatus відіграють неменшу роль переносників вірусу кліщового енцефаліту, чим їх. ricinus. Роль інших видів іксодових кліщів не повинна применшуватися, тому що в деяких районах їхня зараженість може бути значної. Так, наприклад, у Красноярськом краї кліщ Ix. persulcatus у зв'язку з ландшафтною особливістю території майже не зустрічається, що переважає тут видом є Нает. concinna, що і служить основним резервуаром і переносником вірусу в даному вогнищі. У Кустанайській області Казахстану роль основного резервуара і переносника вірусу грає кліщ Dermacentor marginatus. Отже, в окремих ландшафтно-географічних зонах домінуючими переносниками можуть бути Я. concinna, D. marginatus і деякі інші. Однак основними переносниками вірусу кліщового енцефаліту є кліщі Ix. persulcatus і Ix. ricinus.
На людину нападають майже винятково дорослі кліщі, що і визначає основну роль їхній в інфікуванні. Зараження людини відбувається при кровосмоктання інфікованого кліща. Більшість захворівши вказують в анамнезі на присмоктування іксодових кліщів. Частота присмоктування кліщів у захворілих у різних вогнищах складає близько 80%. Імовірність контакту з кліщами у вогнищах кліщового енцефаліту дуже висока. Звичайно кліщі підстерігають годувальників уздовж троп тварин, де вони і виявляються в найбільшій кількості. Кліщі можуть переповзати також на осіб, що доглядають за худобою, що приносить багато кліщів після пасіння, на узліссях лісу. Кліщ проникає під одяг людини і найчастіше присмоктується в області плеча, груди, шиї і голови. Кровосмоктання кліща звичайно невідчутно, тому що його слина містить знеболюючу речовину. Кровосмоктання продовжується 4—6 днів,
Зараження людини кліщовим енцефалітом можливо також аліментарним шляхом — при вживанні сирого молока заражених кіз і корів. Кози хворіють на кліщовий енцефаліт. У заражених тварин спостерігаються втрата апетиту, млявість, менінгеальні симптоми, паралічі задніх кінцівок. У крові кіз вірус знаходиться протягом 3 днів, а в молоці — 8 днів після захворювання. Корови до вірусу не сприйнятливі, але в них також спостерігається вірусемія.
Повсюдно у всіх вогнищах кліщового енцефаліту спостерігається весняно-літня сезонність захворюваності, що обумовлено активністю іксодових кліщів у цей час року. При тривалості епідемічного періоду в 4—5 мес перші одиничні захворювання відзначаються найчастіше в другій половині квітня — першій половині травня. Максимум захворюваності, як правило, реєструється в червні, потім вона знижується і з другої половини липня відзначається знову у виді одиничних, випадків. Перезимовані вірусофорні кліщі є джерелом вірусу енцефаліту, і їхній перший напад на людей знаходиться в прямого зв'язку з настанням весняного потеплення і таненням снігового покриву.
Визначення початку епідемічного сезону дуже важливо для своєчасної організації протиепідемічних заходів (вакцинація, противокліщові заходу). Кінець епідемічного сезону приходиться на липень, але одиничні випадки можуть спостерігатися у вересні, частіше вони відносяться до першої половини серпня.
Сприйнятливість людей до кліщового енцефаліту загальна. Найбільше часто захворювання відзначаються серед облич середнього віку. Професійний склад захворілих визначається особливостями розташування природних вогнищ, характером організаційно-господарської діяльності населення і сформованим укладом життя. Найбільший ризик захворюваності в облич, що безпосередньо працюють у лісі. Серед старожилів поступово з віком формується імунний прошарок, що виникає в результаті латентної чи клінічно вираженої інфекції. Кліщовий енцефаліт після видужання людини залишає тривалий і міцний імунітет.
Природні вогнища кліщового енцефаліту вперше були виявлені в тайгових районах Далекого Сходу. В даний час вогнища відомі в ряді районів лісової зони СРСР — у РСФСР, Української, Білоруської, Казахської, Киргизької, Литовської, Латвійської, і Естонської РСР. У закордонних країнах вогнища кліщового енцефаліту зареєстровані в Чехословакії, Польщі, ГДР, ФРН, Фінляндії, Швеції, Ірландії, Австрії, Угорщини, Югославії, Китаї.
Геморрагічні лихоманки являють собою групу маючих деяка подібність клінічного плину інфекційних захворювань вірусної етиології що супроводжуються домінуючим у картині хвороби геморрагічним синдромом, що розвивається в результаті поразки капілярів і дрібних судин — геморрагічна сип, кровоточивость (носові, легеневі, маткові, шлунково-кишкові кровотечі), поразка бруньок (кров у сечі). Усі вони мають природну очаговість і доведений чи ймовірний трансмісивний механізм передачі вірусу іксодовими і гамазовими чи кліщами іншими ектопаразитами гризунів і інших теплокровних тварин.
Кримська геморагічная лихоманка. Джерелом вірусу є дикі (зайці, ушасті їжаки, птахи) і домашні тварини (велика рогата худоба, верблюди), а також іксодові кліщі роду Hyalomma (Ну. plumbeum, Ну. anatolicum, можливо, Ну. detri-tum). Основний шлях передачі збудника — трансмісивний, через укус іксодових кліщів, що зберігають вірус довічно і передають його трансфазово і трансоваріально. Крім трансмісивного шляху передачі вірусу геморагіченої лихоманки в Середній Азії доведена можливість зараження від хворих при близькому зіткненні з ними (персонал лікувальних установ при відході за хворими), через виділення хворого, особливо кров, що містить великі концентрації вірусу.
У період вірусемії можлива передача вірусу кліщами від людини. Найбільш небезпечними місцями для зараження людини є покриті травою степові ділянки, старі землі під пором, скирти сіна, де в літні місяці виявляється велика кількість кліщів. Пік захворюваності падає на липень — серпень. Сприйнятливі до кримського геморагічної лихоманці обличчя усіх віків або його підлоги. Звичайно занедужують обличчя активного віку в силу більш частого контакту з переносником вірусу. Імунний прошарок серед жителів ендемічних районів практично отсутствует. Майже все населення в потенційних вогнищах не має природного імунітету і піддано захворюванню. Постінфекціонний імунітет носить узкоспецифічний характер, тривалість його не перевищує 1—2 років.
Природні й антропургічні вогнища кримської геморагіческой лихоманки зареєстровані в степових районах Кримського і Керченського півостровів, у Ростовській і Астраханській областях, Ставропольському краї, Туркменській, Узбецькій, Таджицькій, Киргизькій, Казахській, Азербайджанській і Вірменській республіках, у Болгарії, Румунії, Югославії, Пакистану, у ряді країн Африки (Конго, Заїр, Уганда, Нігерія).
Омська геморагічна лихоманка. Хвороба характеризується коротким інкубаційним періодом (2—4 дні), лихоманкою тривалістю від 5 до 15 днів, слабістю, поразкою бруньок, геморагіями на шкірі і слизуватих. Через 3—4 нед симптоми зникають, температура падає, хворі видужують. Летальність досягає 1-5%.
Основний природний резервуар вірусу в природі — іксодові кліщі D. pictus і D. marginatus. Установлена їх спонтанна інфікування, здатність передавати вірус по ходу метаморфоза, а також трансоваріально своєму потомству. Личинки і німфи іксодових кліщів паразитують на гризунах і птахах, а дорослі особи нападають на велику і дрібну рогату худобу і людей. Резервуарами збудника в природі також служать ондатри, водяні полівки, полівки-економки, узкочерепні полівки, бурозубки. Не виключена участь у циркуляції вірусу і деяких інших ссавців (ховрашки, зайці, їжаки, хом'яки), птахів (ворони, граки, випі), жаб і ящірок. У перерахованих тварин установлена спонтанна інфіктування і сприйнятливість до вірусу.
Поряд із трансмісивним і аліментарним шляхами тварини і людина можуть заражатися аерогенним шляхом при вдиханні вірусутримного матеріалу, що попадає в повітря при підсиханні сечі і випорожнень хворих тварин.
У природних водоймах улітку вірус омської геморагіченої лихоманки може залишатися життєздатним протягом 18—20 ч у концентраціях, достатніх для аліментарного інфікування дрібних ссавців. Вірус з великою сталістю виділяється із сечею і випорожненнями тварин, що приводить до інфікування навколишнього середовища. Можливе зараження людини аліментарним шляхом — через воду і харчові продукти, заражені виділеннями хворих гризунів. У цьому випадку захворювання можуть виникнути й узимку. Сприйнятливість до вірусу омської геморагічної лихоманки загальна і висока. Найчастіше хворіють люди, що знаходилися в чи поле в лісі (працівники сільського господарства, мисливці, що беруть участь у промислі на ондатру). Захворювання носять спорадчий чи груповий характер. У місцевих жителів у крові часто виявляються антитіла, тому частіше занедужують приїжджі і Природні вогнища омської геморагічної лихоманки поширені в північній лісостеповій зоні Західного Сибіру і Північного Казахстану, Барабинського степу, Омській, Новосибірській, Оренбурзькій, Тюменській і Курганській областях.
Кіасанурська геморрагічна лихоманка (кіасанурська лісова хвороба) викликається вірусом із групи кліщового енцефаліту. Захворювання в людей виникає після 8-денного інкубаційного періоду і характеризується раптовим підйомом температури до 39°, сильним головним болем, болями в попереку і кінцівках, загальмованістю мови. У такому стані хворі знаходяться протягом перших 2 нед хвороби. З 3—4-го дні хвороби розвиваються нудота, блювота, поноси, сип на м'якому небі, гіпермія кон’юнктив, склер, світлобоязнь, носові, шлунково-кишкові кровотечі, кровохаркання, кровотеча з ясен. Температура нормалізується через 7— 14 днів, кровотечі припиняються на кілька днів пізніше. Може бути друга хвиля захворювання. Період видужання затягується на 3—5 нед. Летальність складає 8—12%. Летальний результат спостерігається звичайно на 7—9-й день хвороби.
Джерелом збудника кіасанурської лісової хвороби є мавпи, що у природних вогнищах хворіють на цю лихоманку і гинуть. У Macaco radiata летальність досягає 85%. Основними переносниками, що заражають людей, є кліщі Haemaphysails spl-nlgera і Нает. turanicum. Вірус виділений також від личинок, німф і імаго кліщів Haemaphy sails turturis, Нает. рариапа, Нает. blsplnosa, Нает. mlnuta, Нает. kyasanurensls, у яких установлена трансоваркальна передача вірусу. Можлива участь у передачі вірусу кліщів пологів Ixodes і Атыуотта.
Багаторазові випадки виділення вірусу з пацюків Rattus blanfordi до із кліщів, що паразитують на гризунах, свідчить про велике значення в циркуляції вірусу також дрібних ссавців (білки, землерийки й ін.). Можливо, відому роль у циркуляції вірусу грають дикобрази і деякі види наземних птахів, що транспортують заражених кліщів. Антитіла до вірусу кіасанурсьої лісової хвороби знайдені в окремих видів кажанів, коней, ослів, верблюдів, овець. Крім трансмісивного шляху передачі не виключається можливість зараження людей через сире молоко корів; це припущення засноване на спостереженні групових захворювань у населених пунктах.
Захворюваність серед людей звичайно виникає в січні і наростає до квітня-травня. Сприйнятливість до вірусу в людей висока. Щорічно виявляються десятки випадків захворювання. Захворювання спостерігаються в облич різних віків, що відвідують лісові масиви і піддаються напад заражених кліщів. Спостерігаються «насінні» спалахи в населених пунктах, зв'язані з аліментарним зараженням. Після перехворіння залишається стійкий імунітет.
Захворювання уперше виявлене в Індії в штаті Майсур (до півдня від Бомбея). В даний час не виключається наявність вогнищ і в інших районах Індії. За межами Індії хвороба не виявлена.
Трансмісивні риккетсиози. Іксодові кліщі є переносниками великої групи хвороб, викликуваних риккетсіями. У ряді випадків кліщі є і резервуарами патогенних риккетсій, забезпечуючи можливість трансовариальної передачі збудника протягом двох-трьох поколінь. Серед риккетсій, зв'язаних із кліщами, слід зазначити 3 триби: Rickettsiae, Ehrlichieae і Wolbachieae. У першій трибі об'єднані всі патогенні для людини види пологів
Rlckettsla і Coxiella. Види роду Rlckettsla на підставі сірологічних особливостей і по поводженню в хребетних і членистоногих відносять до підродів Rlckettsla, Dermacentroxenus і Zinssera. Іноді, цим підродам придається родовий ранг. З іксодовими кліщами особливо тісно зв'язані представники підроду Dermacentroxenus (збудники риккетсиозів групи плямистих лихоманок) і Coxiella bur-neti (збудник лихоманки Ку). При штучному інфікуванні й інших представниках групи, за винятком Zinssera tsutsu-gamushi, успішно розвиваються в організмі іксодових і аргасових кліщів (Weyer, 1964).
У трибу Ehrlichieae входять збудники риккетсиозів нижчих мавп, копитних і хижих. У трибу Wolbachieae поєднуються види, що зустрічаються тільки в комахах і кліщах, причому багато хто з них дуже патогенні для своїх хазяїнів.
Кліщовий риккетсіоз Північної Азії. Збудник захворювання — Dermacentroxenus slblrlcus — малостійкий у зовнішнім середовищі, але протягом усієї зими виживає в кліщах-переносниках. Він може бути виділений із крові й органів диких ссавців, із кліщів, із крові хворих людей. Хвороба характеризується гостролихоманочним плином з наявністю первинного афекту на місці укусу кліща і макулопапулезної висипки. Інкубаційний період складає 3—6 днів. Хвороба починається з продромальних явищ у виді нездужання, головного болю, болю в м'язах, слабості. Сип з'являється на 4—5-й день хвороби і зникає на 10—12-й день, залишаючи після себе пігментацію. Температура швидко досягає високих цифр, тримається 8—10 днів і падає літично. Прогноз сприятливий.
Кліщовий риккетсіоз характеризується природної очаговістю й облігатно-трансмісивним механізмом передачі. Джерелом збудника інфекції є різні види гризунів: польова миша, будинкова миша, пацюк карако, полівка міхно, узкочерепна полівка, пеструха, степова, ховрашок довгохвостий, ховрашок червонощокий, хом'як звичайний, хом'як даурський, бурундук, заєць-русак і ін.
Основними переносниками в Східному Сибіру, Забайкаллі, Тувинської АРСР, МНР є кліщі Dermacentor nuttali; у Західному Сибіру, Алтайському краї — D. plctus, D. marginatus; у Казахстану — D. marginatus; на Далекому Сході (Примор'я, Хабаровський край) — D. sllvarum, Haemaphysalls conclnna в Алма-Атинській обл., у Киргизії — D. marginatus, Haem. punctata; у Вірменії — Rhlplcep-halus sangulneus, D. marginatus.
У природних вогнищах кліщового риккетсіозу Північної Азії збудник перезимовує в організмі дорослих кліщів. Навесні кліщі харчуються на великих диких чи тваринах домашній худобі, звичайно не сприйнятливих до Dermacentroxenus slblrlcus. Зараження личинки і німфи відбувається на гризунах, серед яких у літній період високий відсоток сприйнятливого до інфекцій молодняку. Захворілі після нападу заражених кліщів тварини у свою чергу забезпечують інфікування нових ліній кліщів, підтримуючи в такий спосіб безперервність циркуляції збудника у вогнищі.
Харчування кліщів на імунних тваринах не приводить до зникнення в них риккетсій. Установлено багатомісячне збереження збудника в організмі кліщів, а також його трансфазова і трансова-реальна передача. Від статевозрілих D. marginatus третього покоління удалося виділити риккетсіий згодом 4 роки після зараження батьківського покоління. Перебування риккетсий у кліщах не порушує життєві функції останніх. У іксодових кліщах риккетсії знаходяться переважно в клітках зєднувальнотканних елементів і гемолімфи, у клітках середньої кишки й у слинних залозах. У порожнині кишечнику відзначене зникнення риккетсії незабаром після харчування, а потім повторна поява при руйнуванні заселених ними кліток.
Типовими місцями поширення вогнищ кліщового риккетсіозу є: степовий, лісостеповий і луговий ландшафти і чагарникові зарості, вологі ліси, кочкарники, напівпустелі. Сезонність захворювання — із квітня по жовтень з максимумом у травні, що зв'язано з активністю іксодових кліщів. Кліщовий риккетсиоз Північної Азії уражає в однаковому ступені чоловіків і жінок, незалежно від їхнього віку. Захворювання залишає після себе досить напружений імунітет. Ареал кліщового риккетсіозу охоплює величезний простір від Уралу до берегів Тихого океану, включаючи Далекий Схід, Забайкалье, Сибір, Алтайський край, Казахську і Киргизьку -РСР. За межами СРСР кліщовий риккетсіоз Північної Азії встановлений у східних районах Монгольської Народної Республіки.
Марсельська лихоманка. Збудник марсельської лихоманки — Dermacentoxenus conori — малостійкий у зовнішнім середовищі, характеризується здатністю до внутрішньоядерного розмноження. Інкубаційний період при марсельській лихоманці триває 5—7 днів. Захворювання характеризується гострим початком, ознобом, підвищенням температури до 39—40°, що тримається 10—14 днів. На 4—5-й день хвороби з'являється макулопапулізна сип, частіше на долонях і підошвах. На місці укусу кліща завжди мається первинний афект. Захворюваність звичайно носить спорадичний характер, рідше спостерігаються спалахи. Прогноз сприятливий, летальні ісходи рідкі. Джерелом інфекції, резервуаром і переносником збудника є іксодовий кліщ Rhipicephalis sanguineus (на Чорноморському узбережжі і Середземномор'ї). У країнах Південної і Східної Африки переносниками риккетсії є Rhipicephalus sanguineus, Rh. appendiculalus, Rh. simis, Haemaphysalis laechii, Amblyomma hebraeum і деякі інші види; в Індії і країнах Південно-Східної Азії переносниками служать маловивчені види пологів Haemaphysalis, Ixodes.
У кліщах Rh. sanguineus риккетсії виявлені в клітках кишечнику, слинних залоз, гіподерми і яєчника. Установлено тривале збереження риккетсії в переноснику, включаючи феномени трансфазовій і трансовариальної передачі. Зараження людини відбувається через укуси кліщів, а також у результаті влучення риккетсії на слизуваті око і носа з рук, інфікованих при роздавлюванні кліщів. Захворювання звичайно спостерігаються в літню пору (червень — вересень), що зв'язано з періодом найбільшої активності собачих кліщів в антропургичених вогнищах. У більшості випадків захворювання марсельською лихоманкою зустрічаються в облич у віці від 30 до 70 років, однак хвороба уражає і дітей. Перенесене захворювання залишає напружений імунітет. Випадки повторного захворювання марсельською лихоманкою в літературі не описані.
Захворювання поширене в теплій і субтропічній зонах приморських районів Європи, Африки і частково Азії, у США. Найбільш насичене ендемічне вогнище знаходиться в Середземномор'я. Лихоманка К у. Збудник лихоманки Ку — риккетсії З-xiella s. Rickettsia burneti. Риккетсії Бернета надзвичайно стійкі в зовнішнім середовищі і зберігаються у висушеному стані протягом декількох тижнів і навіть місяців. Лихоманка Ку — гостролихомночне захворювання, що відрізняється від інших риккетсіозів пневмонією. Інкубаційний період триває 10—15 днів. Хворі скаржаться на слабість, сильний головний біль. Захворювання продовжується 1—3 нед, але можуть бути і затяжні випадки з різноманітною клінічною картиною. Летальні наслідки рідкі.
Лихоманка Ку — зоонозне, природно-осередкове трансмісивне захворювання. Джерелами і резервуарами збудника є гризуни й інші тварини. Збудник хвороби виділений від оленів, джейранів, зайців, білок, піщанок, хом'яків, полівок (водяний, сибірської і червоний і ін.), бабаків, ховрашків, тушканчиків сірих пацюків і інших гризунів а також різних птахів: горобців, ластівок, шпаків, дятлів, вівсянок і багатьох інших. Ці тварини в тім чи іншому ступені відіграють роль джерел збудників.
Поряд із природними вогнищами лихоманки. Ку зустрічаються також і антропургичні вогнища, де джерелами інфекції є домашні тваринні і синантропні гризуни. Заражені тварини виділяють риккетсії Бернета із сечею і молоком, вони можуть заражати пасовищних кліщів, що харчуються їх кров'ю. Передача збудника в природних вогнищах від тварини до тварини здійснюється через; членистоногих переносників — іксодових, аргасових, гамазових і краснотелковых кліщів. По ряду біологічних і епідеміологічних особливостей (здатність до фільтрації, виняткова стійкість до абіотичним факторів зовнішнього середовища, поліадаптивність і різноманіття шляхів передачі) риккетсія Бернета помітно відрізняються від інших риккетсії.
У природних вогнищах лихоманки Ку серед різних груп кровососів найбільш тісні зв'язки з риккетсіями Бернета спостерігаються в іксодових, потім в аргасових кліщів. Для іксодід установлені довічне збереження збудника, його трансфазовая і трансова-риальная передача, виділення протягом усього життя разом з фекаліями, а також відсутність інактивації риккетсії в голодуючих особях чи під час зимівлі. Максимальні терміни переживання риккетсії в кліщах і їхнього виділення з фекаліями досягали для їх. гісі-nus 572 днів, а для Ну. asiaticum, включаючи особ дочірнього покоління, — 737 днів, що не є межею (Балашов, Дайтер, 1973).
В організмі кліщів риккетсії розмножуються, проникають у гемолімфу і поширюються в багато органів. Розмноження риккетсії в порожнині середньої кишки немає. Головним місцем існування і розмноження риккетсії Бернета служать травні клітки середньої кишки. При руйнуванні їхній збудник виводиться в просвіт кишечнику і разом з фекаліями виводиться назовні. Фекалії заражених кліщів Ну. asiaticum, D. andersoni і Rh. sanguineus високо инфекціозні. Так, у фекаліях кліща D. andersoni риккетсії виявлялися спустя 586 днів після інфікування. Збудник лихоманки Ку часто зустрічається усередині кліток мальпігієвих судин і в ооцитах. У клітки слинних залоз і в слину риккетсії Бернета проникають не завжди, тому питання про можливість передачі риккетсії іншокулятивним шляхом остаточно не вирішений. Трансовариальна передача риккетсії Бернета іксодовими кліщами непостійна.
У СРСР найбільш розповсюдженими і важливими переносниками збудника лихоманки Ку є іксодові кліщі, у яких риккетсії розмножуються і передаються при кровосмоктанні: Hyalomma asiaticum, Ну. anatolicum anatolicum, Ну. plumbeum plumbeum, Rhipicephalus turanicus, Rh. sanguineus, Ixodes ricinus. Ці види передають кліщів трансфазово і трансовариально. Люди заражаються різними шляхами. Висохлі випорожнення тварин довгостроково (тижня, іноді і місяці) містять життєздатних збудників. Зараження можливе при вдиханні пилу з доріг, де прогоняється худоба, з пасовищ, дворів, де маються тварини-носії; при використанні забрудненої соломи як підстилковий матеріал; при обмолотах забрудненого хліба; на бойнях; при відході за худобою; на виробництвах, що займаються обробкою і переробкою вовни, шкіри і т.п.; у лабораторіях, що працюють зі збудниками лихоманки Ку, і ін. Слід зазначити, що при цьому шляху передачі може мати місце одночасне захворювання великої кількості людей. Аліментарний шлях зараження можливий при вживанні в їжу сирого молока, молочних продуктів, рідше — через м'ясо заражених тварин. Слабокисле середовище сприяє загибелі риккетсій (швидка загибель у кисляку, ацидофіліні). Не виключене зараження людей при уживанні води, овочів, заражених виділеннями тварин. Зараження через ушкоджену шкіру і слизуваті відбувається при влученні на них збудників при зіткненні з хворими тваринами, при відході за ними, при забої й обробленні туш. Не виключається трансмісивний шлях передачі при влученні на людей іксодових кліщів на пасовищах.
Таким чином, роль іксодових кліщів в епідеміології і епізотології лихоманки Ку зводиться до збереження збудників у природі (резервуар) і поширенню їх серед дикі і домашні тварин (переносники), люди заражаються лихоманкою Ку через кліщів відносно рідко. До захворювання сприйнятливі усі вікові групи людей. Зараженню частіше піддані обличчя, що працюють на тваринницьких фермах і підприємствах, зайняті переробкою сировини, що містить риккетсій. Обличчя, яке перенесло лихоманку Ку, здобувають тривалий імунітет.
У нашій країні вогнища лихоманки Ку зустрічаються в різних ландшафтних зонах: пустельної і степовий (південь Середньої Азії, Північний Казахстан), лісостеповий (Алтай, Крим), лісовий (Ленінградська і Кіровска області), субтропічної (Азербайджан). Особливості кожного з типів вогнищ визначаються видовим складом теплокровних тварин і їх ектопаразитів. У закордонних країнах лихоманка Ку відома в США, Новій Зеландії, Західній Європі, Південної і Південно-Східної Азії, Африці і Латинській Америці.
Плямиста лихоманка Скелястих гір. Збудник захворювання — Dermacentroxenus rickettsi — малостійкий у зовнішнім середовищі, при кімнатній температурі зберігається протягом 6—8 днів. Клінічний перебіг хвороби вкрай різноманітно. Інкубаційний період коливається від 2—5 до 14 сут. Звичайне захворювання починається гостро, різким ознобом, швидким підйомом температури, що залишається високої 2—3 нед. На 2—6-і сут з'являється висипка. Загальний стан хворого важке: головний біль, блювота, безсоння, паралічі. Летальність коливається від 5—10 до 80%.
Сприйнятливими хребетними тваринами є гризуни, свинки, кролики, бурундуки, собаки, великі ссавці. Збудники виділені від декількох видів полівок, зайців, бурундуків, ховрашків (Microtus pensilvanicus, Leporis americanus і ін.). Риккетсій виділені і від різних видів іксодових кліщів: Dermacentor ап-dersoni, D. variabilis, Amblyomma cajenensis, Haemaphysalis leporis pallustris у США і Канаді; з Amblyomma cajenensis і A. strlatum у Бразилії; з Rhipicephalus sanguineus у Мексиці; з Haemaphysalis laechii у Східній Африці і від інших видів. У багатьох з них доведена трансфазова і трансовариальна передача збудника,, тому кліщі є не тільки переносниками, але і резервуаром риккетсій.
Основними переносниками збудника є Dermacentor variabilis і D. andersoni, спонтанна інфікованість яким коливається від 1 до 3%. У циркуляції збудника серед гризунів важлива роль належить кліщам Haemaphysalis leporis palustris, Dermacentor parumapestus і Amblyomma americanum. У D. variabilis і D. andersoni установлені трансовариальна передача і передача риккетсій самкам зі спермою інфікованих самців. Трансовариальна: передача спостерігається в 30—40% самок D. variabilis, інфікованих риккетсіями. Риккетсий виявляються в цитоплазмі ооцитів і епітеліальних кліток яєчників кліщів D. andersoni, в особ, що харчуються, кількість риккетсій помітно збільшується.
Поширення риккетсій усередині популяцій кліщів можливо також у процесі запліднення інфікованими самцями незаражених самок під час спарювання їх на великих хребетних, не сприйнятливих до D. rickettsi. Риккетсій при цьому переносяться зі спермою в яйця, що запліднюються, і надалі з останніх розвиваються інфіковані кліщі наступного покоління. У зв'язку зі здатністю одного самця до послідовного запліднення декількох самок цей шлях може бути досить ефективним, хоча відсоток инфікованості потомства залишається не дослідженим.
Риккетсій, що надійшли в організм кліща, попадають потім у клітки середньої кишки, де відбувається їхнє інтенсивне розмноження. З кишечнику частина риккетсій проникає в гемолімфу, а з її в слинні залози, яєчник, мальпігієві судини, м'язову і нервову тканини. Значна кількість риккетсій виділяється з кліщів разом з фекаліями, але цей спосіб передачі збудника не має істотного епідеміологічного значення. Поза організмом кліща здатність, що інфікує, риккетсій швидко губиться, так що головним шляхом передачі служить уведення збудника зі слиною безпосередньо в організм хазяїна. Зараження людей відбувається під час кровосмоктання кліщів, не виключається можливість зараження при влученні матеріалу, що містить збудників (рідина з роздавлених кліщів), на слизуваті чи шкіру.
Перебування риккетсій в організмі кліща впливає на властивості збудника. Періодичне харчування кліщів стимулює розмноження риккетсій, а тривале голодування може привести до полицею втраті збудника. Так, у голодуючих самках і самцях D. variabilis, інфікованих у лічинквої чи нимфальної фазах, швидке зменшення кількості риккетсій в організмі починається на 8-м мес голодування, а через 14—16 мес вони були відсутні у всіх досліджених особей. Перезимовка риккетсій в організмі кліщів, а в ряді випадків і трансфазова передача приводять до утрати вірулентності, що, однак, має оборотний характер. У процесі харчування кліщів на теплокровна тварин вихідна вірулентність штамів легко відновлюється.
Кліщі D. andersoni і D. variabilis відіграють важливу роль у підтримці в природі штамів з високим рівнем вірулентності, оскільки встановлене, що тривале пасуваня риккетсій безпосередньо від хребетних до хребетного приводить до зниження їхньої вірулентності, тоді як пасування через кліщів відновлює вірулентність.
Існують природні й антропугічні вогнища лихоманки Скелястих гір. У природних вогнищах основним переносником у США в західних штатах є Dermacentor andersoni — лісовий кліщ, активний у квітні — травні, у цей час відбувається більшість заражень у лісі, але цей вид може бути на пасовищах домашніх тварин (антропургичні вогнища). Кліщі D. varlabilis (у східних штатах США), Rhipicephalus sanguineus паразитують на собаках, зараження відбувається в населених пунктах (антропургічні вогнища).
До плямистої лихоманки Скелястих гір однаково сприйнятливі усі вікові групи людей. Захворювання залишає після себе тривалий і напружений імунітет. Плямиста лихоманка Скелястих гір поширена в США, Канаді, Мексиці, Бразилії, Колумбії, Африці й Індії. У СРСР захворювання немає.
Бактеріальні хвороби. Туляремія викликається бактерією Francisella tularensis, що має ряд географічних рас: F. t. holarc-tica, розповсюджена в Європі й Азії; F. t. mediaasiatica, виявлена в деяких районах Середньої Азії; F. t. nearctica, найбільш вірулентну, зухвалу туляремію в Північній Америці. Туляре-мийная паличка стійка в зовнішнім середовищі: понад 1 мес — у вологому ґрунті, до 3 мес — у воді, понад 1 мес — у висушеному зерні. Інкубаційний період при туляремії складає 3—7 днів. Це гостре пропасне захворювання з переважною поразкою лімфатичних вузлів, з дуже різноманітними клінічними проявами (бубонна, виразково-бубонна, очна, ангінозно-бубонна, абдомінальна, чи кишкова, і легенева форми). Тривалість плину захворювання варіює від 15—30 днів до 2—3 і більш місяців. Летальність близько 0,5%, в основному при легеневій і абдомінальній формах.
Природна зараженість збудниками туляремії виявлена більш ніж у 60 видів диких і домашніх тварин (гризунів, комахоїдних, хижих, птахів, рептилій і ін.). Основним джерелом інфекції є гризуни, особливо водяники полівки, у яких туляремія може протікати в хронічній формі, не викликаючи падежу, що сприяє збереженню збудника в природі. Важливе значення мають також зайці, бурундуки, ондатри й в антропургічних вогнищах — мишовидні гризуни. Радянськими й іноземними дослідниками виявлено понад 25 видів іксодових кліщів — носіїв збудників туляремії в природі: lx. rlcinus, Ix. persulca-tus, їх. laguri laguri, Ix. apronophorus, Haemaphysalis concinna, Haem. punctata, Dermacentor pictus, D. marginatus, D. nuttali, D. silvarum, D. andersoni, D. variabilis, D. daghestanicus, Ripicephalus rossicus, Rh. pumilio, Hyalomma plumbeum, Amblyomma americanum, Haemaphysalis punctata, Haem. otophila і ін.
Іксодові кліщі є хоронителями і переносниками туляремійного мікроба FT. tularensis. В експерименті здатність до збереження і передачі цього збудника встановлена для багатьох видів іксодід і 5 видів аргасових кліщів. Найбільш ефективними переносниками служать види роду Dermacentor. Можуть заражатися на усіх фазах розвитку при годівлі на тварин з високим рівнем бактерій. Навпроти, при харчуванні личинок D. marginatus на малочутливих до інфекції тварин кліщі поглинають незначну кількість мікробів, недостатнє для наступної трансфазовій передачі. При надходженні в організм достатньої кількості збудника він передається трансфазово по всьому циклі розвитку.
В організмі кліща відбувається розмноження туляремійного мікроба, інтенсивність якого визначається фізіологічним станом переносника. Так, у кліщів Dermacentor pictus, інфікованих у личинковій фазі, кількість збудника в організмі збільшується на стадії нимфального харчування в 1000 разів і харчування самок — у 1000—10000 разів. У той же час на стадії линяння личинок і нимф, кількість збудника в організмі зменшувалося в 100—1000 разів, а при линьке німф і імаго — у 10 разів. У самок цього виду, що наситилися, в організмі міститься до 10 млрд туляремійних мікробів, що в 1000—10000 разів перевершує їхню кількість у личинках, що наситилися. Трансовариальна передача туляремійних мікробів іксодовими кліщами немає. Виділення мікробів у личинок дочірнього покоління можна пояснити поверхневим зараженням яєць при їх відкладкі.
Туляремійний мікроб інтенсивно розмножується і довічно зберігається в клітках середньої кишки. При руйнуванні цих кліток Francisella tularensis надходять у порожнину кишечнику і разом з фекаліями виводяться з організму. Збудник проникає й у гемолімфу кліщів, де розмножується і заселяє слинні і коксальні залози, нервовий ганглій, мальпігієві судини й інші внутрішні органи. Це пояснює можливість передачі збудника туляремії інокулятивним і конитамінативним шляхами. В аргасових кліщів важливу роль може грати також виділення великих кількостей збудника разом з коксальною рідиною.
Люди заражаються туляремією різними шляхами: контактним, аерогенним, аліментарним і трансмісивним. При контактному шляху зараження людини відбувається під час вилову гризунів, зняття з них шкурок; збудник при цьому проникає через шкіру і слизуваті оболонки, викликаючи бубонну, виразково-бубонну, а надалі і септичну форму хвороби. Аерогенний шлях частіше спостерігається при вдиханні пилу, забрудненої випорожненнями хворих гризунів, при молотьбі, перекладці соломи, сіна, віянні, сушінні, навантаженні і розвантаженні зерна, при використанні соломи як підстилковий матеріал. Іноді можливий і повітряно-краплинний шлях зараження — при роботі в лабораторіях з культурами, розкритті трупів гризунів, що упали від туляремії, чи при вдиханні крапельок води, що утворяться під час миття овочів (буряка, картоплі), поверхня яких забруднена виділеннями гризунів.
При аліментарному шляху зараження відбувається у випадку вживання в їжу продуктів, що не піддаються термічній обробці, забруднених випорожненнями гризунів (хліб, сухарі, галети, цукор, сало і т.п.), а також забрудненої, незнезараженої води для питва, умивання, чищення зубів; при купанні у водоймах, по берегах яких спостерігається епізоотія туляремії серед водяних полівок, ондатр. Трансмісивний шлях передачі здійснюється иксодовими і гамазовими кліщами, комарами, ґедзями, мошками, мокрицями і кровосмкчущими мухами при кровосмоктанні чи утиранні їхніх випорожнень у чи шкіру слизуваті оболонки.
Частіше відзначається передача збудника зі слиною в процесі кровосмоктання, причому чим довше кровосмоктання, тим більша кількість мікробів в одиницю часу вводиться кліщем. Можливе зараження гризунів при поїданні ними заражених чи кліщів при злизуванні їхніх випорожнень з вовни. Найбільш ефективними переносниками збудників туляремії є кліщі роду Dermacentor. Число заражених кліщів у вогнищах звичайно близько 1%, іноді досягає 5— 6%. Іксодові кліщі є не тільки переносниками, але і резервуарами збудників, зберігаючи їх у міжепізоотичний період (зимою) у личинках у період їхньої діапаузи. Завдяки цьому непохитне підтримка туляремії в межах осередкових територій звичайно зв'язується з місцями існування іксодових кліщів. У межах природних вогнищ туляремії виділяються численні ландшафтні варіанти з перевагою тих чи інших видів хребетних і безхребетних хазяїнів збудників. Сприйнятливість людини до туляремії майже загальна. Перенесене захворювання залишає стійкий імунітет.
Географічне поширення туляремії обмежене лише північною півкулею. Вона відома в Америці — у США, Канаді, Меконгові, Венесуелі; у Європі — у Франції, Бельгії, Нідерландах, Швейцарії, Італії, Німеччині, Австрії, Чехословакії, Угорщини, Польщі Югославії, Румунії, Болгарії, Греції, Норвегії, Швеції, Фінляндії і СРСР; в Азії (крім СРСР) — у Туреччині, Японії, Китаї.
Інші бактеріальні інфекції. Установлено спонтанне носительство чумного мікроба багатьма видами іксодових кліщів, зібраних у природних вогнищах цієї інфекції. Іксодові кліщі неодноразово виявлялися в природі спонтанно зараженими збудниками чуми. Чумних мікробів знаходили в наступних видів іксодових кліщів: lxodes crenulatus (норовий паразит широкого кола ссавців, у тому числі бабаків), Hyalomma asiaticum asiaticum (хазяїни — верблюди, рідше велика рогата худоба, у молодих форм — ховрашки, піщанки), Haemaphysalis numidiana turanica (норовий паразит ховрашків, піщанок), Haem. schulzei (норовий паразит ховрашків), Rh. pumilio (паразит піщанок, зайців, їжаків) і ін. У Ix. crenulatus. Ну. asiaticum, Rh. schulzei доведена здатність збудника до трансфазовій передачі, але в обмеженому ступені. Граничні терміни збереження Jersinia pestis у кліщах Jx. crenulatus досягали 509 днів. У той же час більшість досвідів із зараженням гризунів шляхом годівлі на них інфікованих кліщів закінчувалося невдачею.
Установлено спонтанну зараженість багатьох видів іксодових кліщів збудником бруцельозу; виділялися культури бруцелл із кліщів, зібраних у природі і неблагополучних по бруцельозі тваринницьких господарствах, чи знятих із хворих тварин. Збудники виявлені в наступних видів: Ну. plumbeum, Ну. scupen.se, D. marginatus, D. pictus, D. daghestanicus, Haem. punctata. Виявлено спонтанну зараженість іксодових кльошів, зібраних у природі, збудниками ряду інших інфекційних хвороб. Збудники листеріозу — Listerella monocitogenes — виділені з кліщів їх. rid-nus, D. pictus, H. anatolicum, Rh. bursa, D. marginatus.
Висновок
Важливою особливістю екології комах і кліщів є необхідність вивчення особливостей життя як окремих видів, так і біоценозів у природній обстановці, вивчення комах як частини природи. До області екології відноситься вивчення явищ, у яких членистоноге виступає не як окрема особину, а як вид, вступаючи в ті чи інші взаємини з іншими видами комах, чи кліщів інших організмів, а також з фізичним середовищем. Таким чином, основним змістом екології є вивчення різноманіття взаємозв'язків організмів і середовища; закономірностей утворення і розвитку популяцій як форми існування виду, їх диференцировки і динаміки, їхньої чисельності; закономірностей формування і розвитку біоценозів як вираження взаємин організмів у конкретних умовах місцеперебування. Центральним об'єктом екології служать зв'язку, взаємини організмів із середовищем у природній обстановці і життя біоценозу в єдності з абіотичною середовищем (біогеоценоз, екологія культурних ландшафтів). Вивчення зв'язку організмів із середовищем складає теоретичну основу сучасної біології. Дані екології кровососних комах і кліщів необхідні для організації ефективних заходів щодо профілактики трансмісивних хвороб і боротьбі з паразитами людини і тварин. Екологія коштує в одному ряді з морфологією, фізіологією й іншими науками, що мають справа з усіма таксономічними підрозділами, що відбивають специфіку організму (зоологією, ботанікою, бактеріологією і більш вузькими — ентомологією, мікробіологією і т.д.).
Екологія, таким чином, є одним з фундаментальних розділів біології. Використовуючи методи практично всіх біологічних наук, екологія встановлює спосіб, якої кожен вид дозволяє основну задачу, — самозбереження і поширення (Беклемишев, І945). Екологія комах і кліщів тісно зв'язана з іншими науками. Наприклад, щоб зрозуміти шкода, заподіювана іксодовим кліщем, і запобігти його, потрібно уміти відрізняти його вид від інших. Цьому учать нас систематика і морфологія. Далі ми повинні знати спосіб життя імаго кліща і його предімагіальних фаз, місця їхній існування, характер харчування, умови, при яких вони нападають на людину, і т.д. Про все це говорить екологія кліщів. Без знання їхніх життєвих відправлень (подиху, травлення, розмноження і т.д.), про які говорить фізіологія, неможливо зрозуміти спосіб життя кліща. Приватні науки зв'язані один з одним і разом з тим зберігають свої особливості.
Література
1. Адамович В.Л. Количественные взаимодействия между хазяином и ектопаразитами в биогеноценозе. - М., 1990
2. Бала шов Ю.С. Кровососущие клещи – переносники болезней человека и животных. – С.-П.: Наука – 1989г.
3. Бала шов Ю.С. Організм иксодидных клещей как середа обитания тарнсмисивных иньекций. – С.-П.: Наука – 1989г.
4. Бей – Бабиенко Г.Я. Общая энтология - 3-е издание М.: Высшая школа 1991г.
5. Белозеров В.Н. Материалы к биологии клеща 2000г.
6. Буренкова Л.А. Екологія клеща М. 1990г.
7. Константинов О.К. Изучение фауны и экологии иксодовых клещей 1994г.
8. Кусов В.Н. Некоторые вопросы размножения клещей 1992г.
9. Павловський Е.Н. Організм как середа обитания 1998г.
10. Павловський Е.Н., Скринник А.Н К биологии клещей 1998г.
11. Померанцев Б.И. Иксодовые клещи 1990г.
12. Тарасов В.В. Членистоногие переносники возбудителей болезней человека 1992г.
13. Чиров П.А Паразитические членистоногие и позвоночные животные 2000г.
14. Шилов И.А. Біосфера, урони организаци жизни и проблемы экологии 1998г.
15. Яхонтов В.В. Екологія насекомых 1995г.
|