Прежде всего, конечно, придется разобраться: а кто, собственно говоря, принимает эти сигналы, кто является тем адресатом, которому предназначена информация? Кто отправитель – вроде бы ясно, по крайней мере, когда речь идет о передаче информации из внешней среды: это органы чувств, на которые действуют физические или химические факторы. Но если мы хотим выражаться более аккуратно, то надо отметить, что всегда, когда мы говорим «мозг посылает сигнал мышцам глаза» или «раздражающий сетчатку сигнал передается в мозг», все равно речь идет на самом деле о передаче сигнала от одной клетки к другой клетке. Так что вся сложная работа нервной системы – регулирование работы внутренних органов, управление движениями, будь то простые и неосознаваемые движения или сложные целенаправленные движения руки живописца – все это, в сущности, основано на «разговоре клеток». Причем все эти «собеседники» вовсе не болтуны: каждая клетка выполняет свою работу, а иногда делает и несколько дел.
Разнообразие функций, выполняемых этим сложным «коллективом», обеспечивается двумя факторами: тем, как клетки соединены между собой, и тем, как устроены «стыки» между клетками.
Что такое синапс
Простейшая реакция нервной системы на внешний раздражитель – это рефлекс. Он осуществляется рефлекторной дугой, строение которой всем хорошо известно. На схеме рефлекторной дуги стрелками обозначен путь сигнала от одной клетки к другой. Этот путь выглядит так, как будто клетки соединены проволочками. На самом деле «проволочки» – это отростки клеток, которые входят в рефлекторную дугу. Давайте присмотримся к более детальному портрету нейрона - элемента рефлекторной дуги. Нервный импульс возникает в теле клетки и распространяется по ее аксону. Аксон заканчивается множеством тоненьких веточек, которые называются терминалями. С этих-то терминалей сигнал и переходит на другие клетки-адресаты: непосредственно на их тела или, чаще, на их «приемные отростки» – дендриты. Аксон может давать до 1 ООО терминалей, оканчивающихся на разных клетках. С другой стороны, типичный нейрон позвоночного получает от 1 ООО до 10 ООО терминалей от других клеток.
Итак, от клетки к клетке сигнал передается через контакт «терминаль – дендрит» или «терминаль – тело клетки». Этот контакт и называется синапсом. Термин «синапс» был введен в физиологию знаменитым английским ученым Ч. Шеррингтоном.
В месте контакта не спайка
, а разрыв
Если говорить о техническом устройстве, то в месте контакта двух участков электрической схемы ничего особенного не происходит: два проводочка соединяются друг с другом – проще всего их спаять.
Среди биологов, изучавших нервную систему, долгое время существовало представление, что мозг – это непрерывная сеть, так что все нервные клетки имеют общую протоплазму, т.е. соединены так, как сосуды кровеносной системы. Считалось, что, например, клетки кожи или мышц – это отдельные клетки, а мозг – единая сеть.
Но вот в 1875 г. итальянский ученый, профессор гистологии университета в Павии К. Гольджи придумал новый способ окраски клеток – серебрение. При серебрении из тысяч лежащих рядом клеток окрашивается одна-единственная, но зато полностью со всеми своими отростками.
Метод Гольджи сильно помог изучению строения нервных клеток. Его использование показало, что, несмотря на то, что в мозгу клетки упакованы чрезвычайно плотно и их отростки перепутаны, все же каждая клетка четко отделяется от другой, т.е. мозг, как и другие ткани, состоит из отдельных, не объединенных в общую сеть клеток. Этот вывод был сделан испанским гистологом С. Рамон и Кахалем, который тем самым распространил клеточную теорию на нервную систему.
Отказ от представления о единой сети означал, что в нервной системе сигнал переходит с клетки на клетку не через прямой электрический контакт, как в электрической схеме, а наоборот, через разрыв.
Когда в биологии стал использоваться электронный микроскоп, эти представления о наличии разрыва получили прямое подтверждение. Оказалось, что и терминаль аксона, и тело клетки-мишени имеют свои собственные мембраны в месте контакта. Между мембранами имеется щель, ширина которой составляет примерно 20 нм.
Как же передается сигнал через этот разрыв?
Какие бывают синапсы. Опять «великий спор»
К тому времени, как было обнаружено наличие таких разрывов в цепи «живой сигнализации», было уже достаточно очевидно, что в решении вопроса о том, как проходит сигнал через этот разрыв, выбирать приходится между двумя способами: электрическим и химическим. В дискуссии о том, какой из этих способов реализуется в природе, участвовали уже знакомые нам Дюбуа-Реймон и его ученик Герман. И по этому вопросу их мнения оказались различными. Герман считал, что одна клетка влияет на другую с помощью местных токов, а Дюбуа-Реймон отдавал предпочтение химическому механизму.
Поясним, в чем существо разногласий, используя современные знания о строении синапсов.
Электрическая гипотеза состоит в том, что нервный импульс, дойдя до терминали клетки-отправителя, вызывает ток в синантической щели, который, затекая в клетка-адресат и вызывает ее возбуждение.
Химическая гипотеза может быть изложена так. Импульс, приходящий по аксону, вызывает в конце терминали выделение химического вещества, которое диффундирует через синаптическую щель и достигает мембраны к летки-мишени. В результате меняется проницаемость этой мембраны и возникает ток, который течет через синаптическую щель и через мембрану тела клетки.
Итак, какими же являются синапсы: «электрическими» или «химическими»?
Первые достаточно прямые эксперименты говорили в пользу химической гипотезы. Отто Леви, раздражая блуждающий нерв лягушки* собрал внеклеточную жидкость, омывающую сердце, на которое действовал этот нерв, и подействовал этой жидкостью на сердце другой лягушки. И вот без всякого раздражения нерва сердце второй лягушки тоже стало биться реже. Правда, в данном случае речь шла не о синапсах между двумя нервными клетками, а о синапсе между нервом и сердечной мышцей. Но, проведя в Казани сходный по идее опыт на нервном ганглии, А.В. Кибяков показал, что и между нервными клетками существует химическая передача. Такой опыт свидетельствовал, что из окончаний раздражаемого нерва выделяется какое-то вещество, которое само по себе может действовать на другие нейроны или на мышцу.
Итак, химическая теория торжествовала. Некоторые медиаторы, которые вначале были столь же гипотетичны, как клеточная мембрана, были выделены в чистом виде и их химическое строение было определено. С помощью микроэлектродов, введенных в клетку и аксон, было выяснено, что время, затрачиваемое на выделение медиатора из терминали и его диффузию через щель, составляет примерно 0,6–0,8 мс у теплокровных животных.
Казалось, что химическая передача в синапсах доказана, а электрическая не находила никаких подтверждений. Но как можно было найти электрический синапс? Надо было показать, что есть синапсы, которые не обладают существенными признаками химических.
Первым таким признаком является временная задержка, те самые 0,8 мс; в случае электрического синапса ждать задержки не было оснований. Другим отличительным свойством химических реакций является их сильная температурная зависимость. Чем выше температура, тем быстрее идет реакция. В случае электрических синапсов ожидать такой зависимости также не приходилось. Кроме того, при изучении химических синапсов было показано, что для их работы нужны ионы кальция в среде, омывающей клетки. Замена кальция на магний блокировала передачу сигнала через химический синапс. И вот в 1957 г. с помощью микроэлектродов был открыт синапс, в котором сигнал передавался практически без задержки, передача слабо зависела от температуры и не блокировалась магнием. Был открыт первый чисто электрический синапс. Это показалось странным исключением, да и синапс открыли всего лишь у какого-то речного рака. Но лиха беда начало. Вскоре электрические синапсы открыли у рыб,
кошек,
обезьян.
Как мыуже неоднократно видели это и раньше снова оказался прав Ходжа Насреддин, утверждая, чтооба спорщика правы. Сейчас четко показано, чтосуществуют и химические и электрические синапсы,
Электрические синапсы существуют, но их не может быть
Полученные в эксперименте доказательства передачи сигнала через синапс чисто электрическим путем вступили в противоречие с существовавшими на тот момент теоретическими расчетами. Сложилась парадоксальная ситуация: электрические синапсы есть, существуют, их существование доказано прямыми экспериментами, а расчеты показывают, что они не могут работать!
Действительно, как показало серебрение, а потом и электронная микроскопия, непосредственного контакта между клетками все-таки нет: клетки разделены щелью, заполненной жидкостью, через которую ток пойдет не только в клетку-мишень, но и вытечет куда-то «на сторону». Расчеты, проведенные в разных лабораториях мира, дали обескураживающие результаты. Оказалось, что при реальных экспериментально известных значениях сопротивлений мембран, межклеточной среды и размеров синаптических контактов и щелей в клетку-мишень будет затекать не более 0,01% всего тока, вытекающего из терминали. Этот ток к тому же растечется по всему телу клетки и не сможет создать изменения ее потенциала, необходимого для возбуждения или сопоставимого с реально измеряемыми изменениями.
За решение этой задачи в 1965 г. взялась группа молодых сотрудников Теоретического отдела Института биофизики АН СССР.
Первая их идея состояла в том, что в синапсе и сопротивление мембраны, и сопротивление межклеточного вещества могут быть другими, отличаться от сопротивления клеточной мембраны и внеклеточной среды. Ведь известные значения этих параметров были получены совсем на других объектах. Так что в расчетах, о которых говорилось выше, достоверными можно было считать только размеры синаптической области. Да и методы расчетов были весьма приближенными.
Для проверки выдвинутой гипотезы надо было повторить расчеты для разных значений этих неизвестных параметров; кроме того, требовалось усовершенствовать и сам метод расчета, чего никак не удавалось сделать. Тогда решили сделать аналоговую модель синапса.
Эту работу поручили дипломнику, который паял схемы, все время перепаивая их для новых значений параметров, подбирая сопротивления, причем сопротивлений с нужными параметрами, как обычно бывает, в лаборатории не оказывалось, и приходилось соединять несколько имеющихся. Канители было много и работа продвигалась медленно. А тут еще пришло лето и отделу предложили выделить двух сотрудников для поездки на месяц в колхоз. Никому не хотелось на столь долгий срок прерывать работу, и сотрудники предложили другой выход: поехать всем вместе, но зато на короткий срок. Жаль было только, что в полевых условиях нельзя ставить эксперименты и паять новые схемы.
Нужда научит калачи есть
Но этот отрыв от паяльника и сопротивлений оказал благотворное действие. Единственными доступными орудиями научного труда тут были карандаш и бумага. И вот Володе Смоляшгаову удалось заменить физическую модель математической: описать связь всех величин, характеризующих синапс, математическими функциями. Необходимый точный метод расчета был найден.
Вернувшись в лабораторию, засели за математические таблицы, линейки и прочие математические приборы того времени. Результаты работы на такой математической модели казались, на первый взгляд, совершенно невероятными.
Оказалось, что, хотя и существует некоторое оптимальное сопротивление мембраны в синапсе, при котором в клетку-мишень попадала самая большая часть тока, все равно эффективность такого синапса была несравнима с реальной. Если сопротивление мембраны бралось ниже оптимального, то увеличивалась утечка тока через щель, если сопротивление увеличивалось, то падала общая сила токах
вытекающего из терминали.
Другой результат был уже совсем странным. По модели получалось, чтоулучшить связь между клетками можно, если… их лучше изолироватьдруг от друга! Через синапс,щель которого заполнена веществом с высоким удельнымсопротивлением, терминаль можетвозбудить клетку-мишень, если только синапс имеет подходящие размеры: диаметр в несколько микрометров и, как обычно, тонкую щель. Этот результат, немного подумав, легко понять и на качественном уровне. Высокое сопротивление внутрищелевого вещества будет мало мешать току, текущему из терминали в клетку, так как в этом направлении его слой тонок, но зато будет эффективно препятствовать вытеканию тока через щель, так как в этом направлении сопротивление изолирующего слоя велико. Дело, как видите, опять оказалось в геометрии. Беда только, что электрические синапсы с щелью* заполненной жиром, не были известны в природе.
Отрицательный результат – тоже результат
Модель доказала, что электрический синапс не может работать ни при каком сопротивлении мембраны, одинаковом на всей поверхности синапса. И тогда был сделан следующий шаг в единственном возможном направлении: предположили, что мембрана синапса неоднородна по сопротивлению – она имеет «окна» с низким сопротивлением в центральных областях и высокое сопротивление у края синапса.
Эта гипотеза оказалась верной. С усовершенствованием методов электронной микроскопии в разных лабораториях мира было обнаружено, что, действительно, в ЭС используется неоднородная мембрана, но ее неоднородность создается особым способом: с помощьюспециального белка – коннектина. Молекулы этого белка имеются и в мембране терминали, и в мембране клетки-мишени и образуют там особую структуру – коннексон, состоящую из шести молекул и имеющую внутриканал. Когда аксон дорастает до клетки-мишени, дваконнексона соседних мембран соединяются друг с другом ив каждом из них открывается отверстие – канал, который до того закрыт. Этот канал представляет низкое сопротивление для прохождения ионов. В ЭС много коннексонов. Таким образом, ЭС связывает две клетки множеством тоненьких трубочек диаметром около 1 – 1,5 нм, проходящих внутри белковых молекул.
Сделаем небольшое отступление в сторону. Какую же роль сыграло в данном случае математическое моделирование синапса? Ситуация тут достаточно парадоксальная.
Если бы удалось найти параметры, соответствующие передаче тока в реальных синапсах, и модель удовлетворяла бы экспериментальным данным, то результат был бы не столь ценен. Он только означал бы, что ЭС может быть устроен так, как предполагалось в начале, но он мог быть устроен и иначе. Возможно, существуют десятки моделей, которые одинаково хорошо соответствуют эксперименту. Но отрицательный результат моделирования является абсолютным. Он строго говорит, что система не может быть устроена так, как мы предполагаем, и следует искать,
как же она устроена на самом деле.
Замечательно, что и второй результат исследования ЭС на математической модели, несмотря на свою кажущуюся несообразность, также нашел экспериментальное подтверждение. Оказалось, что природа действительно использует заполнение щели изолятором. У птиц в цепочке нейронов, обеспечивающих реакцию зрачка на свет, был обнаружен очень большой по диаметру ЭС, щель которого заполнена миелином, т.е. изолятором. Для такого большого синапса этот способ весьма эффективен, как и предсказывала модель. Таким образом, в природе действительно используются и заполнение синаптической щели изолятором, и неоднородные мембраны.
Химический синапс
В отличие от электрических синапсов, которые так или иначе служат для образования непрерывного пути для электрического сигнала, химические синапсы – это настоящие «разрывы»: электрическому сигналу «напрямую» через химический синапс не пройти.
Рассмотрим для примера работу нервно-мышечного синапса. Как вы знаете, мембрана мышечного волокна возбудима: если прикоснуться к ней электродом, мышца сократится. Оказывается, существует парадоксальное исключение: именно в том месте мембраны, через которое приходит сигнал от мотонейрона к сокращению, мышца не чувствительна к электрическому воздействию! Значит, мотонейрон воздействует на мышцу совершенно иным способом: в его терминали электрический сигнал преобразуется в химический.
Общую схему работы химического синапса мы уже описывали. Когда нервный импульс доходит до терминали мотонейрона под его воздействием в синаптическую щель выделяется особое вещество – медиатор. Для нервно-мышечного синапса позвоночных таким медиатором служит ацетилхолин. Вот к этому-то веществу и чувствительна постсинаптическая мембрана. Стоит только поднести к области синапса пипетку с раствором, содержащим ацетилхолин, и вывести немного ацетилхолина наружу, как в мышечном волокне возникает изменение потенциала, тем более сильное, чем больше ацетилхолина выведено из пипетки. Если эти изменения потенциала достигают порога возбуждения электровозбудимой мембраны, то в ней возникает ПД и мышечное волокно сокращается.
Выделение медиатора
В 1950 г. английские ученые Фетт и Катц, изучая работу нервно-мышечного синапса лягушки, обнаружили, что без всякого действия на нерв в мышце в области постсинаптической мембраны сами по себе через случайные промежутки времени возникают небольшие колебания потенциала, амплитудой примерно в 0,5 мВ, которые они назвали «миниатюрные потенциалы». Когда Фетт и Катц подействовали на синапс ядом, о котором было известно, что он блокирует выделение ацетилхолина из терминалей, миниатюрные потенциалы исчезли.
Получалось, что в химическом синапсе медиатор выделяется и в покое, но изредка и небольшими определенными порциями.
К тому времени электронная микроскопия уже достаточно много знала об устройстве синапсов. В частности, было известно, что в пресинаптических окончаниях имеются какие-то пузырьки диаметром примерно 500 нм. Иногда удавалось увидеть, что мембрана этих пузырьков сливается с мембраной терминали. Катц выдвинул гипотезу, что миниатюрные потенциалы возникают тогда, когда такой пузырек слипается с пресинаптической мембраной, разрывается и выбрасывает в синаптиче-скую щель свой запас ацетилхолина. Частично эту гипотезу удалось проверить цитологам: они сумели выделить везикулы и показать, что в них действительно содержится «цетилхолин.
Итак, в покое, когда мотонейрон не возбужден, в синапсе существует небольшой электрический шум: за счет случайного выброса ацетилхолина возникают миниатюрные потенциалы; средняя частота их – примерно один в секунду. Но амплитуда этих одиночных потенциалов слишком мала, и поэтому мышечное волокно не возбуждается. Когда же к терминали мотонейрона подходит ПД, то происходит массовое опорожнение везикул: за 0,1 мс лопаются примерно 100 пузырьков. И все они выбрасывают свой ацетилхолин в синаптическую щель. В результате в мышечном волокне возникает деполяризация в 30 – 50 мВ, что гораздо выше порогового значения.
Естественно было посмотреть, как влияет изменение МП терминали на частоту миниатюрных потенциалов, т.е. на частоту выброса ацетилхолина. Оказалось, что при деполяризации терминали частота выделения везикул возрастает экспоненциально. Это объясняет, почему под действием нервного импульса выделяется так много ацетилхолина.
Но решение одного вопроса, как всегда, порождает новые: почему деполяризация приводит к тому, что везикулы начинают чаще «прилипать» к мембране?
Целым рядом экспериментов было выяснено, что «виновником» возрастания частоты выделения медиатора являются ионы кальция. Оказалось, что в пресинаптической терминали имеются потенциалзависимые кальциевые каналы, которые открываются при деполяризации терминали, и кальций входит в терминаль.
Этот процесс был очень красиво показан Ллинасом и др. в 1972 г. на крупном синапсе кальмара, где имеется возможность вводить разные вещества в терминаль. Есть такое вещество – экворин. При соприкосновении с ионами кальция он дает световую вспышку. Ллинас и его сотрудники ввели 'экворин в пресинаптическую терминаль. Когда к синапсу приходил нервный импульс, пресинаптическое окончание вспыхивало, показывая, что внутрь него вошли ионы кальция. Но ученые не ограничились этой красивой иллюстрацией уже известных к тому моменту фактов и поставили новый опыт, который дал важный результат. Они ввели через микроэлектрод внутрь окончания ионы кальция, и тут же без всякой деполяризации началось выделение медиатора.
Итак, оказывается, что причиной выделения медиатора является не деполяризация сама по себе, а то, что деполяризация открывает дорогу кальцию внутрь терминали. И если убрать из наружной среды кальций, то как показали эксперименты, химический синапс не сработает ни при какой деполяризации и даже миниатюрные потенциалы исчезнут.
Но есть еще один очень важный вопрос' почему после попадания кальция внутрь терминали медиатор выделяется только очень короткое время, а потом его выделение прекращается? Кальциевые каналы терминали открываются лишь на очень короткое время, но за это время концентрация кальция в терминали повышается в тысячи раз. Куда же девается этот кальций? Оказывается, он быстро выкачивается наружу кальциевым насосом и поглощается митохондриями, которые всегда присутствуют в синаптических окончаниях.
После этих работ был выяснен механизм действия некоторых ядов. Например, из яда паука каракурта был выделен белок – латротоксин, который по существу представляет собой незакрывающиеся кальциевые каналы. Он встраивается в пресинаптическую мембрану и начинает пропускать в терминаль кальций. В результате запасы ацетилхолина в терминали полностью истощаются и нервная система не может вызвать сокращения мышц.
Но что же делает кальций, попав внутрь терминали? На этот вопрос пока нет однозначного ответа. Возможно, кальций просто экранирует отрицательные заряды мембраны везикул и мембраны терминали, что позволяет им слиться; возможно, кальций вызывает сокращение «внутриклеточных мышц», которые подтягивают пузырек к мембране; а может быть, механизм действия кальция окажется совсем иным и неожиданным.
Работа постсинаптической мембраны
Что же делает ацетилхолин с постсинаптической мембраной, к которой он диффундирует? В покое удельное сопротивление постсинаптической мембраны равно примерно 3 ООО Ом-см2
, но при действии ацетилхолина оно снижается до 1 Ом-см2
. Это сразу наводит на мысль, что ацетилхолин открывает какие-то каналы в постсинаптической мембране. И это предположение верно.
В постсинаптической мембране находятся каналы, «ворота» которых управляются не МП, а ацетилхолином, И канал, и ворота являются частями особой сложно устроенной белковой молекулы. Конец такой молекулы, торчащий из мембраны наружу, «узнает» молекулы ацетилхолина. Если с ним связываются две молекулы – ацетилхолина, то открывается канал, через который могут проходить ионы К+
и Ка+
. Иными словами, в мембране открывается «электрическая дырка» со всеми вытекающими последствиями, а именно деполяризацией мембраны. Таким образом, постсинаптическая мембрана преобразует химический сигнал вновь в электрический сигнал – деполяризацию мембраны.
Возникает естественный вопрос: почему эта деполяризация исчезает? Ведь действие химического синапса обычно кратковременно. Значит, ацетилхолин, открывающий каналы в постсинаптической мембране, куда-то девается. Оказывается, медиатор связывается с холинорецептором очень непрочно: открыв ворота канала, он отрывается и вновь уходит в синаптическую щель. А в щели имеется особый фермент, который его разрушает. Так что медиатор – вещество очень «скромное»: сделав свое дело, он тут же уходит. Именно это его свойство обеспечивает кратковременность действия химического синапса.
Есть и другие вещества, которые тоже могут связываться с холинорецептором, но делают это лучше, чем ацетилхолин. Например, растительный яд кураре, которым индейцы смазывали свои стрелы, попав в организм, прочно связывается с холинорецепторами мышцы и занимает «посадочные площадки», предназначенные для ацетилхолина. В результате ацетилхолин не может открыть ворота каналов и любые движения становятся невозможными. На этом же основано и действие некоторых змеиных ядов; так, в частности, действуют яды змей семейства аспидовых. Интересно, что и у растений, и у змей в ходе эволюции вырабатывались яды со сходным механизмом действия. Вещества, сходные с кураре, релаксанты, сейчас используются в медицине во время операций: они расслабляют мышцы.
Сейчас известно довольно много деталей об устройстве и работе «молекулярной машины» – холинорецептора. Холинорецептор состоит из пяти субъединиц – участков. Изучение аминокислотных последовательностей этих единиц показало, что все они произошли в результате модификации одного и того же гена. Соединяясь между собой, эти субъединицы образуют в центре канал. Этот канал имеет примерно квадратное сечение со стороной квадрата 0,65 нм. Он не различает ионов К+
и Ка+
, но не пропускает анионы. Ворота канала открываются на случайные промежутки времени. Проводимость такого канала равна примерно 3-Ю»11
С. Длина холинорецептора в 5–6 раз превышает толщину мембраны, так что белок сильно торчит из нее наружу и внутрь клетки. Ацетилхолин, выброшенный одним пузырьком, открывает около 2 ООО каналов,
Какие синапсы лучше – электрические или химические?
Из нашего рассказа вы поняли, что электрические синапсы устроены просто, химические – несложней, тем более, что мы до сих пор разобрали только один из многих видов ХС – нервно-мышечный. Как же осуществляется разделение труда между этими двумя видами синапсов? Обычно в организме они используются в таких нервных цепях, где их свойства оказываются наиболее полезными.
Например, отличительной особенностью ЭС является быстродействие. Естественно, что ЭС находятся в тех цепях, которые обслуживают срочные реакции организма: отдергивание тела, отпрыгивание, убегание.
Например, у дождевых червей вдоль всего тела проходят гигантские «аксоны». Как видите, они не такие гигантские, как у кальмара, да и устроены они иначе. На самом деле это не аксон, т.е. не отросток одной клетки. Этот «аксон» состоит из множества цилиндрических кусочков. В каждом сегменте тела есть нервная клетка, которая отращивает такой кусочек; затем торцевые мембраны этих цилиндров соединяются коннексонами, так что получается кабель с перегородками, пронизанными каналами коннексонов. В результате импульс бежит по этому составному аксону как по обычному толстому нервному волокну. Эти волокна вызывают быстрое сокращение тела червя, обеспечивая реакции отдергивания от раздражителя или быстрого втягивания в норку. При химических синапсах эта реакция занимала бы несколько десятых долей секунды: ведь задержка между сегментами в ХС холоднокровного составляет несколько миллисекунд, а сегментов может быть несколько десятков и даже сотня; задержка наЭ что отбрасывает рака назад, подальше от опасности. ЭС имеются в системах спасения бегством у некоторых медуз, моллюсков, рыб и других животных.
Вторая характерная особенность ЭС состоит в том, что они пропускают сигнал в обе стороны – они симметричны). Это может способствовать синхронизации, т.е., одновременности возбуждения нейронов* связанных ЭС. Такие синхронизирующие системы довольно распространены в живой природе. В простейшем случае они состоят всего из двух клеток; связь этих клеток через ЭС приводит к тому, что при возбуждении одного из нейронов практически одновременно возникает импульс и во втором. Например, у электрического сома такая система из двух нейронов обеспечивает одновременность разряда электрических органов обоих сторон тела. У другой рыбы, которая умеет, сокращая плавательный пузырь, издавать звуки, напоминающие кваканье все клетки, управляющие мышцами плавательного пузыря, связаны ЭС, что позволяетвозбуждаться практически одновременно.
Электрические синапсы довольно «модны» у беспозвоночных и низших позвоночных. У высших позвоночных большинство синапсов – химические. Использование в организмах ХС связано с их характерной особенностью – преобразованием электрического сигнала в химический и обратно. В них одинаковые электрические импульсы могут вызывать выделение самых разных веществ-медиаторов; например, у кольчатых червей в нервно-мышечных синапсах используется тот же ацетилхолин, что и у позвоночных, а у членистоногих, которые произошли в ходе эволюции от кольчатых червей* используется в таких синапсах совсем другой медиатор – глутамат. С другой стороны, в ХС один и тот же медиатор, действуя на разные клетки-мишени, может открывать совершенно разные каналы. Например, ацетилхолин в одних случаях открывает чисто калиевые или чисто натриевые каналы в других – каналы, пропускающие и калий, и натрий, в третьих – каналы, пропускающие только хлор, и т.д. Поэтому ХС может выполнять более сложные функции, чем ЭС. Так, одно из самых важных явлений в нейрофизиологии – явление торможения – практически всегда осуществляется ХС),.
Химический синапс и торможение
Известно, что человек волевым усилием может остановить безусловный рефлекс. Существует, например, защитный рефлекс отдергивания руки, когда рука касается горячего предмета, чего-то острого и т.д. Однако известен пример римского героя Сцеволы, который положил руку на горящую жаровню и не отдернул ее, преодолев боль. Каждый из нас затормаживает рефлекс отдергивания, когда берут кровь на анализ. Вменее явной форме торможение проявляется почти во всяком поведенческом акте, в том числе ив непроизвольных движениях, торможение участвует и в регуляции работы внутренних органов.
Что же такое торможение? В чем его причина? По этому поводу в литературе до сих пор можно встретить самые фантастические утверждения. Даже в школьном учебнике физиологии говорится, что торможение в данном месте нервной системы вызывается тем, что сильно возбуждается соседний участок. Так что же такое торможение на самом деле?
Прежде всего подчеркнем, что под торможением понимается активный процесс, а не просто отсутствие возбуждения. Заторможенной называют клетку, в которой какой-то механизм противодействует возбуждению. Вспомнив, что такое возбуждение, легко понять, что это за механизм. Мы знаем, что нервная клетка или волокно возбуждаются тогда, когда деполяризуется их мембрана. Значит, противоположный сдвиг мембранного потенциала и будет торможением: такую клетку будет труднее возбудить, так как потребуется более сильное воздействие, чтобы довести ее потенциал до порогового значения»
Как можно гиперполяризовать, т.е. затормозить клетку? Вспомнив опять, «что снаружи, что внутри», легко сообразить, что для этого надо либо усиливать проницаемость мембран для ионов калия, которые будут выносить наружу положительный заряд, либо увеличивать проницаемость мембраны для ионов хлора, которых много в наружной среде. Перемещение отрицательных ионов хлора внутрь клетки даст тот же эффект, что и перемещение положительных ионов калия наружу.
В нервной системе встречаются тормозные синапсы, использующие и калий, и хлор. Как правило, при этом используются особые тормозные медиаторы, которые управляют воротами соответствующих каналов. Например, у позвоночных есть два тормозных медиатора – аминокислота глицин и гаммааминомасляная кислота, которые, в основном, открывают хлорные каналы мембраны. Интересно, что гаммааминомасляная кислота является тормозным медиатором не только у позвоночных, но и у членистоногих.
Существуют и другие способы торможения. Например, известно, что блуждающий нерв тормозит деятельность сердца. Блуждающий нерв выделяет ацетилхолин, точно такой же, как тот, который возбуждает скелетные мышцы, но сердце он тормозит. Оказывается, в случае сердца ацетилхолин действует не прямо на ворота каналов. Он садится на особые рецепторы, активация которых меняет метаболизм сердечных клеток. В результате ряда внутриклеточных реакций возникает особое вещество, которое и открывает изнутри ворота калиевых каналов. Такие синапсы называют «метаболическими».
Итак, мы видели, что в ХС с помощью разных медиаторов могут открываться те или иные каналы клеточной мембраны. Если при этом возникает деполяризация – синапс возбуждающий, если возникает гиперполяризация – синапс тормозный.
О величине синаптических потенциалов
Мы с вами рассмотрели принципы работы возбуждающих итормозных синапсов. Посмотрим теперь, как можно оценить количественно действие химических синапсов.
Из рассказа о потенциале покоя и потенциале действия вы знаете, что для каждого иона существует свой равновесный потенциал, при котором число ионов, входящих в клетку и выходящих из клетки, становится одинаковым. В покое для ионов калия равновесный потенциал равен примерно –80 мВ; при возбуждении, когда в основном открываются натриевые каналы, равновесный потенциал для натрия равен примерно +40 мВ. У постсинаптической мембраны тоже есть свой равновесный потенциал. Его величина зависит от того, какие ионы пропускает эта мембрана. Например, постсинаптическая мембрана возбуждающего синапса, каналы которой в равной мере пропускают и калий, и натрий, имеет равновесный потенциал, лежащий ровно посередине между таковыми для калия и натрия: /2 = –20 мВ. А у тормозного синапса, пропускающего ионы хлора, равновесный потенциал равен примерно –80 мВ.
Пока медиатор не подействовал на постсинаптическую мембрану, ее каналы закрыты и ток через нее не течет. Под действием медиатора открываются каналы для тех или иных ионов и они будут идти через постсинаптическую мембрану тем эффективнее, чем дальше отстоит ее потенциал от равновесного. Можно сказать, что в области постсинаптической мембраны включается источник э.д.с. величиной, где Ус
– равновесный потенциал постсинаптической мембраны, а V – мембранный потенциал клетки в данный момент. Если мембранный потенциал равен равновесному для данного синапса, то ток через синапс не пойдет.
Представим теперь эквивалентную электрическую схему нейрона с действующим на него синапсом. При выделении медиатора будет течь синаптический ток силакоторого по закону Ома для всей цепи равна

Здесь. Добщ – сопротивление всей клеточной мембраны, а Лс
– сопротивление синапса. Этот ток создает падение напряжения на сопротивлении внесинаптическои мембраны:

Этот сдвиг потенциала называют постсинаптическим потенциалом. Если синапс возбуждающий, сдвиг потенциала называют возбуждающим постсинаптическим потенциалом, если же синапс тормозный, то сдвиг потенциала называют тормозным постсинаптическим потенциалом
Давайте попробуем оценить величину ВПСП, создаваемого одним синапсом на мотонейроне). Равновесный потенциал этого синапса лежит в области –20 мВ. Сопротивление мембраны мотонейрона равно примерно 107
Ом. Сопротивление среднего синапса площадью в 1 мкм2
равно примерно 1012
Ом. Отсюда АУ –
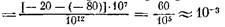 мВ. мВ.
Мы видим, что один синапс создает крайне малый сдвиг потенциала, ведь порог возбуждения 10–15 мВ. Значит, чтобы возбудить мотонейрон, на него должно подействовать много синапсов.
Наша формула годится в том случае, если синаптический ток имеет достаточно большую длительность, тогда все емкости успевают зарядиться и их можно не учитывать. Для кратковременных синаптических токов надо учитывать и емкость мембраны.
Устройства, подобные синапсам
Оказывается, что устройства, подобные синапсам, как электрическим, так и химическим, играют важную роль в жизнедеятельности самых разных тканей и органов.
Например, клетки сердца у самых разных животных связаны каналами из того же белка коннектина, который образует каналы в ЭС. В результате электрический сигнал распространяется по сердечной мышце от клетки к клетке за счет тех же местных токов,
что и по гигантскому «аксону» червя» Связаны коннексонами между собой и клетки гладких мышц разных внутренних органов.
Это еще не самое удивительное: все это возбудимые ткани, в которых должен распространяться электрический сигнал. Но вот уж совсем удивительным было открытие в середине 60-х годов американским биологом Левенштейном и сотрудниками Лаборатории молекулярной биологии им. А.Н. Белозерского МГУ того, что и невозбудимые клетки разных органов тоже связаны высокопроницаемыми контактами. Фактически почти все ткани организма представляют собой не скопление одиночных клеток, а единый коллектив, в котором клетки могут обмениваться через каналы высокопроницаемых контактов разнообразными молекулами. Благодаря этому в тканях возможна своеобразная «клеточная взаимопомощь», Например, если в какой-то клетке плохо работают насосы, ее соседи через каналы высокопроницаемых контактов «делятся» с ней своим ионным запасом и поддерживают ее потенциал покоя.
При изучении высокопроницаемых контактов было выяснено, что коннексоны являются не стабильными трубками, а динамическими структурами: каналы, образуемые коннектином, могут открываться и закрываться под действием разных факторов. Сейчас выяснен молекулярный механизм такого закрывания каналов. Коннексон состоит из 6 субъединиц, которые могут двигаться относительно друг друга, при этом отверстие может закрываться; это устройство очень похоже на устройство диафрагмы фотоаппарата с подвижными лепестками.
Зачем же нужно это свойство коннексонов? Рассмотрим один пример. Обычно в цитоплазме клеток очень мало свободного кальция. По ряду причин более высокие концентрации кальция ведут к гибели клетки, и поэтому у клеток есть ряд защитных механизмов: избытки кальция выкачиваются наружу насосами, поглощаются митохондриями и т.п. Представим себе теперь, что в системе клеток, связанных высокопроницаемыми контактами, какая-то из клеток серьезно повреждена. Защитные механизмы не могут справиться с избытком кальция, поступающего из наружной среды, и клетка погибает. Но когда кальций подходит внутри этой клетки к области клеточного контакта, коннексоны закрываются и соседние клетки отсоединяются от поврежденной. Такая конструкция похожа на систему водонепроницаемых переборок в военных кораблях: если в одном отсеке возникла пробоина, перегородки автоматически закрываются и корабль не тонет. В клеточных системах такое устройство тоже обеспечивает их высокую живучесть. Если предположить, что все сердце представляло бы собой систему клеток, прямо связанных своей цитоплазмой, тогда повреждение могло бы распространяться от клетки к клетке. Один известный ученый сказал: «Клетки сердца работают вместе, а умирают поодиночке». Теперь мы знаем, что это возможно именно благодаря свойствам коннексонов.
Но динамичность коннексонов важна не только для создания живучести. Оказалось, что высокопроницаемые контакты можно найти уже на самых ранних стадиях развития зародышей разных животных; они соединяют между собой клетки, возникающие уже при первых дроблениях яйца, а в ходе дальнейшего развития то появляются, то исчезают. Клетки то влияют друг на друга какими-то веществами, то участки зародыша изолируются друг от друга;
и тогда в этих участках развивается однородная ткань из одинаковых клеток; потом такие участки вновь соединяются контактами с соседями, и вся эта сложная игра контактов важна для регуляции нормального развития.
Все, что мы узнали о высокопроницаемых контактах, невольно наводит на мысль, что передача сигналов в электрических синапсах – это вторичная «профессия» структуры, которая, как и ионные насосы, играет более общую и фундаментальную роль в развитии организмов и функционировании их тканей.
Аналогичная ситуация имеется и с химическими синапсами. Принцип их работы используется в организмах не только для передачи информации, но и в других целях. Оказывается, разнообразные секреторные клетки используют ионы Са++
для регуляции выброса секрета подобно тому, как в химическом синапсе этот процесс используется для выброса медиатора. Кроме того, клетки многих желез являются электрически возбудимыми.
Рассмотрим для примера работу клеток поджелудочной железы, вырабатывающих инсулин, регулирующий содержание сахара в крови.
Оказывается, на поверхности этих клеток имеются специальные рецепторы, реагирующие на глюкозу. Если концентрация глюкозы в крови выше нормы, то под действием этих рецепторов клетки деполяризуются и в них возникают потенциалы действия. Эти потенциалы действия имеют кальциевую природу, они возникают за счет открывания Са-каналов. При этом ионы Са++
входят внутрь клетки, что приводит к выбросу в кровь инсулина, точно так же, как в случае нервных окончаний приводит к выбросу медиатора. Роль кальция в выбросе разных веществ, в частности гормонов, показана и для многих других желез.
Нервная клетка – клетка
Этими словами мы хотим подчеркнуть, что особенные, исключительные свойства нейрона – модификация свойств, присущих и другим клеткам организма, В ходе естественного отбора молекулярные машины и разные клеточные устройства приобретают в разных клетках несколько разные функции, при этом сами устройства могут либо несколько меняться, либо использоваться в том же виде, но для другой цели. Раньше мы уже говорили, что ионные насосы и потенциалы покоя имеются у всех клеток организма, а в нервных и мышечных клетках используются для передачи сигналов.
Рассказ о синапсах дает нам особенно много примеров того, как единство превращается в многообразие. Мы видели, что коннексоны существуют у самых разных клеток организма, в том числе и невозбудимых. Они используются и при регуляции эмбрионального развития, и для передачи молекул от клетки к клетке, а в возбудимых тканях этот межклеточный канал был использован эволюцией для передачи электрического тока, для создания электрических синапсов.
Разнообразные клетки организма выделяют в окружающую среду разные вещества; прежде всего, это клетки желез. Существуют специальные клеточные приспособления для выброса секретируемых веществ: эти вещества упакованы в мембранные контейнеры, а их выброс регулируется ионами Са++
, которые входят в клетку через специальные кальциевые каналы. В результате естественного отбора этот механизм используется нервными клетками в конструкции химических синапсов: в контейнерах содержится медиатор, а дальше выброс его организован так же, как выброс гормонов и других веществ. С этой точки зрения нервные клетки с химическими синапсами – это один из вариантов секреторных клеток, а медиатор – это их секрет, который только не просто выбрасывается в кровь, а поступает к совершенно определенным потребителям через синаптическую щель.
С другой стороны, мы видели, что работа клеток поджелудочной железы похожа на работу нервных клеток. Они тоже имеют потенциал действия, но этот потенциал действия служит для того, чтобы открыть кальциевые каналы и впустить внутрь ионы Са++
. Да и у многих мышечных клеток основная роль потенциала действия состоит в том, чтобы открыть ворота кальциевых каналов для ионов Са++
, запускающих процесс сокращения. Мы еще раз видим близкое сходство механизмов, используемых разными клетками организма в разных целях: для передачи сигнала в ЭС, для выброса секрета в клетках желез, для сокращения в клетках мышц. В конце концов, это совсем не удивительно, ведь все они – потомки одной и той же клетки и имеют идентичный геном.
Большинство клеток организма имеют на своей поверхности специальные белки-рецепторы, чтобы реагировать на действие гормонов, на содержание в крови разных веществ и т.д. Клетки иммунной системы используют такие рецепторы, чтобы отличить свои белки от чужеродных, клетки дыхательной системы – чтобы реагировать на концентрацию углекислоты и т.д. Нервные клетки используют рецепторы постсинаптической мембраны для того, чтобы в ответ на медиатор открыть дорогу тем или иным ионам и изменить мембранный потенциал. Сейчас открыто много метаболических синапсов, где эффект синапса осуществляется белками-рецепторами через изменение внутриклеточных химических реакций, что еще больше роднит их с другими типами клеточных рецепторов.
Часто говорят, что развитие науки приводит к все большей изоляции разных ее направлений, так что настоящий специалист – это человек, который знает все ни о чем или о чем-то бесконечно малом. Как мы видим, в биологии можно найти прямо противоположный пример. Чем глубже мы узнаем механизмы работы клеток и организмов, тем яснее нам становится видно проявление единых общих принципов работы, которые эволюция остроумно приспосабливает для достижения разнообразных целей.
|