МИНИСТЕРСТВО НАУКИ И ОБРАЗОВАНИЯ РОССИЙСКОЙ
ФЕДЕРАЦИИ
ГОУ ВПО «НОВОСИБИРСКИЙ ГОСУДАРСТВЕННЫЙ
ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ»
ИНСТИТУТ ЕСТЕСТВЕННЫХ И
СОЦИАЛЬНО-ЭКОНОМИЧЕСКИХ НАУК
КАФЕДРА БОТАНИКИ И ЭКОЛОГИИ
РЕФЕРАТ ПО ФИЗИОЛОГИИ РАСТЕНИЙ
Тема: Катаболизм и стресс у растений
Выполнил:
Калиниченко Н.А.
320 группа
Проверила: д.б.н., профессор
Барахтенова Л.А.
Новосибирск 2007
Содержание:
1. Введение...............................................................................................................3
2. Значение и функции катаболизма для растения...............................................4
3. Биогенный стресс и катаболизм.......................................................................12
4. Заключение.........................................................................................................20
5. Список литературы............................................................................................25
1. ВВЕДЕНИЕ
"В организмах совершается не только процесс создания, т.е. питания и роста, но, рука об руку с ним, идет процесс разрушения и выделения".
"Организм не ассимилирующий теряет в весе, - этим обнаруживается его разрушение; но оно идет и одновременно с ассимиляцией, о чем мы судим по продуктам выделения. Без этого процесса разрушения невозможны проявления жизни. Жизненная деятельность находится, разумеется, в известных пределах, в прямом отношении с этим процессом разрушения. Чем энергичнее трата, тем энергичнее жизненная деятельность. Только совокупность двух процессов, созидания и разрушения, характеризует живое тело. Организм живет только пока разрушается".
К.А. Тимирязев
Обмен веществ слагается из множества одновременно протекающих реакций, среди которых можно выделить два важнейших противоположно направленных процесса: анаболизм и катаболизм - синтез и деструкция. В данном реферате мы рассмотрим различные аспекты катаболитических процессов наблюдаемых у растений при стрессе.
Большинство современных исследований направлено на изучение синтетических процессов, что определяется не только их очевидной важностью, но и появлением и совершенствованием эффективных методов исследования, таких, как радиоактивные индикаторы, хроматография, электрофорез и т.д.
Изучение катаболизма биополимеров и липидов сопряжено с гораздо большими трудностями, чем их синтеза. Например, если поставлена задача изучения метаболизма определенного белка, то его необходимо "подметить" с помощью того или иного изотопа, ввести белок в заданный компартмент клетки и затем прослеживать за особенностями его превращения.
Проведение всех трех операций связано со значительными трудностями. Это главным образом и предопределяет неизмеримо меньшее число публикаций, посвященных процессам катаболизма биополимеров и липидов.
Другая причина - недооценка функциональной роли процессов деградации сложных соединений. Сейчас существует мало публикаций где реакции клеточного метаболизма представлены в виде трех функциональных блоков - катаболизма, анаболизма, а также роста и дифференцировки. [2, стр.8].
2. ЗНАЧЕНИЕ И ФУНКЦИИ КАТАБОЛИЗМА ДЛЯ РАСТЕНИЯ
Можно говорить о множественной роли процессов катаболизма.
Во-первых, роль корректирующего фактора (рис. 1). Он обеспечивает, устранение биополимеров с «некорректной» структурой, вызванной тремя возможными причинами: ошибками в ходе образования этих соединений; большей или меньшей потерей их активности в результате последующей модификации; таким изменением окружающей среды, при котором эти биополимеры или липиды не могут в полной мере выполнять свои функции [2, стр.18].
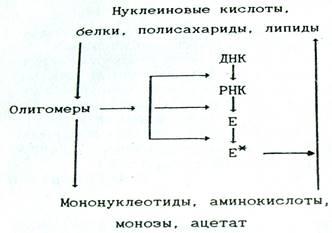
Рис. 1. Схема взаимосвязи катаболизма и анаболизма, отражающая субстратную и регуляторную функции катаболизма
Во-вторых, субстратная роль. Процессы катаболизма обеспечивают синтез биополимеров мономерными субстратами, а синтез липидов - ацетатом, что особенно важно в условиях недостаточного насыщения клеток этими веществами.
В-третьих, энергетическая роль. При дальнейшей деградации мономерных продуктов и ацетата и вовлечении их в процессы дыхания образуются АТФ и НАД(Ф)Н, обеспечивающие протекание анаболических реакций.
В-четвертых, сигнальная функция. Известно, что особенности структуры нуклеиновых кислот, белков и некоторых поли- и олигосахаридов (в том числе входящих в состав гликопротеидов и гликолипидов) используются в клетках для хранения, передачи и реализации информации [2, стр.19].
В процессе деградации биополимеров происходит постепенное уменьшение заключенного в этих молекулах объема информации. Однако трудно себе представить, что в ходе эволюции не были использованы информационные свойства фрагментов молекул биополимеров, образующихся при деструкционном каскаде в результате действия эндо- и экзогидролаз. Исследования в этом направлении показали, что, во-первых, частичная деградация предобразованных молекул является широко распространенным способом формирования функциональных молекул нуклеиновых кислот и белков в реакциях соответственно посттранскрипционного процессинга РНК и посттрансляционной модификации белков.
Во-вторых, накапливается все больше информации о том, что некоторые олигонуклеотиды, олигопептиды и олигосахариды являющиеся продуктами катаболизма соответствующих полимеров обладав свойствами гормонов или активаторов и ингибиторов различных процессов метаболизма и в конечном итоге оказывают влияние на рост и морфогенез растений.
Можно считать, что сигнальные свойства олигомерных промежуточных продуктов катаболизма реализуются путем воздействия на транскрипцию, трансляцию или на предобразованные ферменты связанные с процессами анаболизма, а также на мембранные структуры (см. рис. 1)
Приведенные выше рассуждения о сигнальных свойствах олигомерных производных биополимеров справедливы и в отношении промежуточных продуктов деградации липидов, в состав которых входят полиеновые (ненасыщенные) жирные кислоты — линолевая, линоленовая, а у низших грибов и бактерий - арахидоновая и эйкоэапентаеновая, обладающие соответственно двумя, тремя, четырьмя и пятью двойными связями. Сигнальные свойства полиеновых жирных кислот определяются реакциями оксигенирования молекул по местам двойных связей и последующими реакциями, в результате осуществления которых появляется множество производных жирных кислот. Некоторые из этих продуктов оксигенирования являются физиологически активными веществами.
Деградация мембранных липидов может приводить к драматическим последствиям, в связи с влиянием на физико-химические свойства мембран и условия функционирования катализирующих разнообразные процессы мембранных белков-ферментов, транслокаторных систем, рецепторов и т.д.
Скорость деградации и окислительных реакций в мембранных липидах имеет большое значение для регуляторной и информационной роли мембран в клеточном метаболизме[9, стр.557].
Некоторые из промежуточных продуктов метаболизма биополимеров и липидов в ходе эволюции животных и растительных организмов стали выполнять функции гормонов - физиологически активных веществ, вырабатываемых в одних органах, транспортируемых в другие и изменяющих в них интенсивность и направленность метаболизма и в конечном итоге влияющих на реализацию программ роста, морфогенеза, устойчивости и др.
Принципиальное отличие этих систем у растительных и животных организмов в том, что у первых отсутствуют полипептидные или олигопептидные гормоны, а в роли гормонов стали выступать менее сложные вещества, образующиеся, например, при модификации мономерных продуктов катаболизма белков (триптофан - ауксин, метионин – этилен) нуклеиновых кислот (пуриновые нуклеотиды - цитокинины) или липидов (жирные кислоты - жасмонат) [9, стр.559].
Большинство же физиологически активных олигомерных продуктов катаболизма биополимеров и оксигенированных производных полиеновых жирных кислот выполняет роль не гормонов, а внутриклеточных эндогенных регуляторов, не транспортируемых в другие органы и ткани. Они могут играть роль микровинта в корректировании метаболических процессов, в то время как грубая настройка осуществляется фитогормонами.
Еще один пример использования продуктов деградации биополимеров и липидов, выработавшийся в ходе эволюции, - это появление системы вторичных мессенджеров (инозитолтрифосфатов, диацилглицеролов, циклических нуклеотидов), функционирование которых показано в растениях и образование которых есть результат взаимодействия рецепторов плазмалеммы с гормонами и гидролиза мембранных фосфолипидов (в случае Са 2+
-фосфоинозитольной системы), а также преобразования свободных нуклеотидов (синтез циклических нуклеотидов). Они, в свою очередь, вызывают фосфорилирование белков. Имеется мнение, что в результате фосфолирования белков протеинкиназами, активированными вторичными посредниками, повышается в целом активность ферментов катаболических реакций, фосфорилирование же ферментов анаболических реакций подавляет их активность [9, стр.557].
Особенно сильная активация катаболизма наблюдается при начинающемся стрессе. Известно, что ответная реакция растений на действие эколого-климатических (физических) и антропогенных (преимущественно химических), а также биогенных стрессоров включает несколько стадий:
1) тревоги,
2) адаптации,
3) истощения.
Одной из главных черт первой стадии и является резкое усиление процессов катаболизма липидов и биополимеров[8, стр.281].
Целесообразность усиления катаболизма определяется необходимостью повышения концентраций мономерных субстратов и ацетата для синтеза новых биополимеров и липидов, которые в большей степени соответствуют условиям стресса, а также необходимостью поддержания при этом энергетики клеток за счет сгорания продуктов катаболизма в процессе дыхания. Быстрое повышение концентрации субстратов для синтезов и обеспечение их макроэргическими соединениями и восстановителем создают условия для замещения одних распадающихся форм биополимеров и липидов другими[8, стр.289].
Еще один аспект катаболизма - включение сигнала тревоги - интенсивное образование олигомерных физиологически активных продуктов катаболизма полимерных молекул и оксигенированных производных липидов, в том числе тех фитогормонов, которые являются продуктами катаболизма. Эти сигналы тревоги приводят в действие сложную систему настройки клеток и организма в целом на экстремальные условия существования. Одним из компонентов этой системы настройки является усиление синтеза специфических (шоковых) соединений, способствующих повышению устойчивости клеток. В случае действия патогенов может наблюдаться, наоборот, сильное повышение чувствительности клеток вокруг места внедрения патогена и их гибель, но это повышает устойчивость организма, так как создает препятствия для дальнейшего распространения инфекции[8, стр.296].
Усиление деградации липидов и биополимеров - это лишь одна из многих неспецифических ответных реакций, отражающих структурно-функциональную перестройку растительного организма, попавшего в экстремальные условия.
Вполне естественно, что обнаруживаются и специфические черты ответа, зависящие от вида стрессора (в особенности от биогенного, от патогена), но по мере усиления меры его действия на первый план во все большей степени начинают выступать неспецифические изменения.
К числу наиболее значительных неспецифических изменений можно отнести следующие:
1. Фазность в развертывании во времени ответа на действие стрессора.
2. Усиление катаболизма липидов и биополимеров.
3. Изменение проницаемости мембран клеток для ионов.
4. Повышение в цитоплазме содержания ионов кальция.
5. Подкисление цитоплазмы.
6. Снижение общей интенсивности синтеза биополимеров и липидов.
7. Синтез стрессовых (шоковых) белков.
8. Интенсификация синтеза компонентов клеточных стенок - лигнина, суберина, кутина, каллозы, богатого оксипролином белка.
9. Накопление пролина, и связано это с его свойствами осморегулятора
10. Накопление органических полиаминов.
11. Повышение содержания абсцизовой и жасмоновой кислот.
12. Продукция этилена.
13. Торможение фотосинтеза.
14. Усиление дыхания.
15. Перераспределение углерода из СО2
, усвоенного в процессе фотосинтеза, среди различных соединений - уменьшение включения метки в высокополимерные соединения (белки, крахмал) и сахарозу и усиление (чаще относительно - в процентах от усвоенного углерода) - в аланин, малат, аспартат. Положение о неспецифических изменениях в углеродном метаболизме, вызванных действием различных стрессоров, и о непосредственных причинах этих изменений было впервые выдвинуто автором более четверти века тому назад [1, стр.3].
16. Повышение содержания свободных радикалов.
17. Образование в ответ на действие биострессоров (бактерии, грибов, вирусов, насекомых) элиситоров и фитоалексинов фенилпропаноидной и терпеноидной природы, разнообразных патогениндуцированных белков, хитиназ, β-1, 3-глюканаз, ингибиторов протеиназ[1, стр.4].
Неблагоприятные условия существования - климатические, биогенные, антропогенные- действуют на обмен веществ и ультраструктуру клеток в том же направлении, что и старение. Более того, они ускоряют его. Можно считать, что описания особенностей процессов быстрого старения у таких обычно используемых для этого объектов, как лепестки цветов в значительной степени относятся и к растениям, подвергшимся действию неблагоприятных факторов. Это в первую очередь касается отсеченных лепестков, часто используемых в качестве модели для изучения старения. Дело в том, что отделение какого-либо органа или вычленение ткани растения само по себе вызывает у них состояние стресса. Это продемонстрировано на отделенных листьях и особенно полно на отсеченных корешках проростков.
Например,в некоторых работах показано, что фазы стресса (реакция тревоги и реакция адаптации) реализуются в процессе адаптивного старения отсеченных корней [2, стр.120]. Отсечение корня от проростка вызывает в начальный период (1-2 ч после отсечения) ряд изменений в метаболизме, которые можно отнести к реакции тревоги или предадаптации. Это - падение мембранного потенциала, усиление генерации активных форм кислорода, повышение выхода из клеток ионов калия (вероятно, по калиевым каналам), увеличение содержания свободных жирных кислот, повышение степени ненасыщенности жирнокислотных остатков мембранных фосфолипидов, активация митохондриального дыхания.
Все эти изменения рассматриваются как основа последующих восстановительных процессов, которые можно отнести к стадии адаптации. Во время стадии адаптации происходят восстановление мембранного потенциала, снижение генерации активных форм кислорода, поглощение ионов калия (возможно, вследствие активации протонных насосов). Одновременно повышается степень насыщенности жирнокислотных остатков мембранных фосфолипидов, что является основой снижения степени проводимости мембран для ионов [3, стр.57].
Старение организмов приводит к постепенному уменьшению величины отношения анаболизм/катаболизм. В различных органах растений степень усиления катаболизма с увеличением общего возраста организма может отличаться в зависимости от собственного возраста, например, у метамерных органов (листьев). Усиление катаболизма старых листьев способствует «перекачке» образующихся мономеров (и продуктов их превращения в легко транспортируемые формы) и реутилизации органических соединений в более молодых или репродуктивных органах. Среди неспецифических ультраструктурных изменений, вызванных стрессом и старением, можно отметить изменение тонкой гранулярной структуры ядра, уменьшение числа полисом и диктиосом, набухание митохондрий и хлоропластов, уменьшение в хлоропластах числа тилакоидов, перестройку цитоскелета [3, стр.157].
В изучении процессов деградации у растений важную роль играет выбор объекта исследований. Например, сильная интенсификация катаболизма биополимеров и липидов наблюдается у прорастающих семян; в органах растений с быстрым темпом старения (например, у лепестков некоторых видов растений); у затемненных листьев; в участках корней, заканчивающих рост растяжением [4, стр.197]; в тканях и клетках, разрушающихся в результате механического повреждения, подвергающихся действию бактерий, грибов, вирусов, эколого - климатических или антропогенных стрессоров. Во многих случаях используются изолированные протопласты, культуры клеток или каллусных тканей растений.
Анализ событий, совершающихся в клетках с момента начала действия того или иного стрессора, заставляет считать, что имеется небольшое число наиболее лабильных (чувствительных) компонентов клеточных структур и звеньев метаболизма, которые выполняют роль пускового механизма, вызывающего последующие глобальные нарушения, проявляющиеся в виде неспецифических изменений в обмене веществ.
По всей вероятности, функцию спускового крючка, на которых "нажимают" разнообразные стрессоры и который приводит в действие сложный механизм ответа, в большинстве случаев выполняют мембранные системы.
Мембраны - это "ахиллесова пята" клеток, в первую очередь повреждающаяся под влиянием экстремальных факторов. Совершенно очевидно, что во многих случаях, например, при изменении температуры, роль сенсорного компонента мембран играют липиды, от состава и соотношения которых зависит степень вязкости мембран, определяющая структуру и функции мембранных белков.
Изменение состояния мембран приводит к включению процесса деградации липидов, от которого зависит реализация программы формирования стресса растительных клеток[4, стр.197].
3. БИОГЕННЫЙ СТРЕСС
И
КАТАБОЛИЗМ
В предыдущем разделе уже затрагивался вопрос о влиянии патогенов на образование углеводных элиситоров и белковый обмен. В последние годы появляется все больше информации о метаболическом взаимодействии патогенных организмов - бактерий и грибов - и тканей различных органов растения-хозяина. Первый этап этого взаимодействия - конформационное узнавание поверхностей органов и клеток патогена и хозяина. Второй - экскреция клетками патогенных бактерий и грибов ферментов, гидролизующих биополимеры и липиды покровных тканей растения-хозяина, разрыхляющих их и обеспечивающих более интенсивное проникновение патогена в ткани хозяина и обильное питание, необходимое для развития патогена. Третий - образование в ходе деградации биополимеров и липидов различных физиологически активных промежуточных продуктов - элиситоров, выполняющих роль сигнальных веществ - стимуляторов ответной реакции клеток хозяина. Элиситоры непосредственно или с помощью посредников влияют на генетический аппарат клеток хозяина, вызывая (четвертый этап) синтез веществ, способствующих или повышению устойчивости к патогену, или вызывающих сверхчувствительность и гибель клеток, но тем самым создающих механический барьер, препятствующий распространению инфекции по тканям растения (рис. 2). Промежуточные продукты катаболизма клеток хозяина могут выступать и в роли эффекторов метаболизма и развития патогена [5, стр.44].
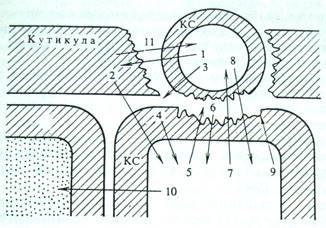
Рис. 2. Схема взаимодействия клетки патогена с растением- хозяином
1 — кутиназа; 2 — продукты деградации компонентов кутикулы (возможно, обладающие сигнальными свойствами); 3 — β-глюканаза и другие гликозилазы, экскретируемые патогеном; 4 — элиситоры — фрагменты клеточной стенки (КС) хозяина; 5 — хитиназы и другие гликозилазы, действующие разрушающе на КС патогена; 6 — элиситоры — фрагменты КС патогена; 7 — фитоалексины — ингибиторы протеиназ, кутиназ, гликозилиз и других ферментов патогена; 8 — токсические вещества патогена; 9 — укрепление КС хозяина за счет активации пероксидаз и усиления синтеза лигнина. Отложение оксипролиновых белков, лектинов; 10 — вещества — индукторы сверхчувствительности и некроза соседних клеток; 11 — продукты деградации кутина, действующие на клетку патогена
Наружным покровом растений является кутикула, состоящая главным образом из гетерополимера кутина, погруженного в воск. Обнаружено более 20 мономеров, из которых состоит кутин: различной длины насыщенные и ненасыщенные жирные кислоты и спирты, в том числе гидроксилированные и эпоксидированные, дикарбоксиловые кислоты и т.д. В кутине большинство первичных спиртовых групп участвует в образовании эфирных связей, так же как часть вторичных спиртовых групп, обеспечивающих сшивки между цепями и точки ветвления в полимере. Другой "барьерный" полимер - суберин, состоит, по-видимому, из фенольных и алифатических доменов, первые из которых близки по своему составу к лигнину, а вторые - к кутину. Отличия алифатического домена в том, что свободные жирные кислоты являются главным компонентом субериновых восков, в то время как в кутине их очень мало. Кроме того, в суберине присутствуют главным образом С22
и С24
жирные спирты, в то время как в кутине – С26
и С 28
[5, стр.44].
Оказалось, что многие патогенные грибы могут выделять ферменты, гидролизующие кутин и суберин. При гидролизе лигниноподобной фракции суберина образовывались кумаровая и феруловая кислоты, причем большая часть фракции оставалась негидролизованной. Продуктами кутиназной реакции были различные оксигенированные жирные кислоты и спирты [5, стр.45].
По всей вероятности, в спорах грибов кутиназа содержится в очень небольших количествах, и при контакте с кутикулой растений гидролизу подвергается лишь малая часть кутина. Однако образующиеся активные сигнальные молекулы - 10,16-дигидрокси-С16
- и 9,10,18-тригидрокси-С18
-кислоты - транспортируются в прорастающую спору и индуцируют образование больших количеств дополнительной кутиназы, начинающей интенсивное разложение кутина и облегчающей инфицирование растения. Было обнаружено, что лаг-период появления кутиназной м-РНК после начала действия ди- и триоксикислот составляет всего 15 мин, а появления кутиназы - в два раза больший [5, стр.45]. Ингибирование кутиназы с помощью химических препаратов или антител предотвращало инфекцию.
Одной из важных задач исследователей молекулярных взаимодействий между патогеном и тканями растения-хозяина были поиски сигнальных веществ, индуцирующих защитную реакцию инфицированных клеток и системную - в удаленных от места инфекции местах.
Вполне вероятно, что продукты деградации кутина (оксигенированные жирные кислоты и спирты) могут выступать в роли не только индукторов образования кутиназы у патогена, но и элиситоров синтеза защитных веществ в клетках растения-хозяина (см. рис.2).
К числу высокоэффективных элиситоров защитного ответа инфицированных растений относятся олигомерные продукты деградации полисахаридов клеточных стенок хозяина или патогена[5, стр.45].
Гидролитические ферменты (в том числе гликопротеины) патогенных грибов активируют защитную реакцию хозяина, освобождая биологически активные углеводы из клеточных стенок хозяина или патогена.
Было найдено, что суспензионные клетки табака вырабатывали фитоалексины (капсидиол и др.) в ответ на обработку целлюлазой, что свидетельствует об элиситорных свойствах линейных β-1,4-глюкановых фрагментов целлюлозы. В этой связи представляют интерес данные об активирующем действии дисахарида целлобиозы на синтез целлюлозы из 14
С-глюкозы или из меченой УДФГ в клетках волосков семян хлопчатника. Так как в клетках не содержится эндогенной целлобиозы, то, по-видимому, экзогенная целлобиоза служит миметиком олигосахаридов более высокой степени полимеризации[9, стр.562].
В формировании ответной реакции растений на патогены принимают участие также полиеновые жирные кислоты.
Оказалось, что элиситорным эффектом обладает не белковая часть липопротеинов, а их липидная часть, представляющая собой не свойственные для высших растений арахидоновую (эйкозатетраеновую) и эйкозопентаеновую кислоты. Они вызывали образование фитоалексинов, некротизацию тканей и системную устойчивость растений к различным патогенам. Продукты липоксигеназного превращения в тканях растений упомянутых выше С20
жирных кислот (гидроперокси-, гидрокси-, оксо-, циклические производные, лейкотриены), образующиеся с помощью имеющегося в клетках хозяина ферментного липоксигеназного комплекса (субстратами которого могут быть как C18
, так и С20
полиеновые жирные кислоты), оказывали сильнейшее влияние на защитную реакцию растений. Это объясняется, по-видимому, тем, что в неинфицированных растениях нет оксигенированных производных 20-углеродных жирных кислот, и их появление в результате инфицирования приводит к драматическим результатам, например к гибели клеток и образованию некрозов, что создает барьер для распространения инфекции. Имеются данные, что индуцирование патогеном липоксигеназной активности приводило к формированию ответной реакции растения и в том случае, когда элиситор не содержал С20
жирных кислот и субстратом липоксигеназной активности могли быть только собственные полиеновые жирные кислоты, а продуктами - октадеканоиды, а не эйкозаноиды. В связи с этим представляют большой интерес сведения о том, что жасмонат может индуцировать синтез ингибиторов протеиназ[10, стр.562]. Знаменательно, что глюкан и Са2+
усиливали влияние арахидоната и эйкозапентаеноата. Так как ЭГТА (специфический лиганд Са2+
) ингибировал синтез фито-алексинов, то можно сделать предположение, что ионы кальция играют важную роль в регуляции осуществления защитной функции растений[5, стр.44].
Не исключено, что сигнальными веществами являются и продукты деградации белков клеточных стенок, богатых оксипролиновыми остатками и содержащих олигогликозильные ответвления.
Под влиянием воздействия многообразных элиситоров в инфицированных растениях начинают вырабатываться защитные вещества, повышающие устойчивость клеток к инфекции. К ним относятся в первую очередь фитоалексины - растительные антибиотики, представляющие собой соединения фенилпропаноидного и терпеноидного характера. Например, у картофеля под влиянием Phytophtorainfestans образуются ришитин и любимин[2, стр.123].
Элиситоры индуцируют образование большого количества белков, как правило, не характерных для неинфицированных тканей.
1. Патогениндуцированные белки, представляющие собой группу щелочных и кислых белков с относительно небольшой молекулярной массой (10-20 кДа), функции которых в большинстве случаев не выяснены.
2. Хитиназы и β -глюканазы, накапливающиеся в вакуоле и клеточных стенках. Интересно, что b-глюканаза "созревает" в две стадии. Первая заключается в удалении с N-конца олигопептида (насчитывающего 21 остаток аминокислот) и в N-гликозилировании С-конца белка, вторая - в отщеплении от образовавшегося промежуточного белка олигопептида с 22 аминокислотными остатками, включая углеводную ветвь, присоединившуюся на предыдущей стадии.
3. Ингибиторы протеиназ, вырабатываемые как в результате механического повреждения тканей (например, листогрызущими насекомыми), так и в результате инфицирования патогенами. Их синтез вызван фрагментами пектиновых веществ клеточных стенок. Интересно, что индукция ингибиторов протеиназ сопровождалась фосфорилированием белков плазмалеммы клеток хозяина.
4. Серусодержащие белки - тионины, высокотоксичные для грибов. Как уже отмечалось выше, наблюдается также значительное усиление
образования оксипролиновых белков (в том числе ферментов, например пероксидазы, от активности которой зависит синтез лигнина), входящих в состав клеточных стенок.
Интенсивное новообразование различных белков является отражением перестройки метаболизма инфицированных растений, приводящей к нарастанию устойчивости к патогену[9, стр.557].
Хитиназы из зерна пшеницы, ячменя и других растений обладали свойствами эндохитиназ, в то время как бактериальные ферменты проявляли экзохитиназную активность.
Хитин (поли-М-ацетилглюкозамин) является компонентом клеточных стенок грибов и членистоногих. В них содержатся и хитиназы, которые наряду с хитин-синтетазными комплексами определяют особенности структуры хитинсодержащих клеточных стенок. Однако хитиназа обнаруживается и у организмов, не содержащих хитина: у почвенных бактерий (экзохитиназа, отщепляющая по очереди концевые N-ацетилглюкозные остатки) как инструмент добывания пищи и у растений (эндохитиназа) как инструмент защиты от грибной инфекции и от некоторых насекомых. Интересно, что хитиназа растений как индивидуальный белок обладает также свойствами лизоцима. Основными продуктами деградации хитина были хитобиоза, хитотриоза и хитотетраоза.
"Антигрибные" хитиназы, по-видимому, широко распространены в царстве растений, в стеблях и листьях индуцируясь этиленом или атакой патогенов, а в семенах запасаясь как средство повышения устойчивости к грибам почвы. Хитиназы растений действуют прямо на растущие кончики гифов гриба, вероятно, вместе с другими гидролазами подавляя рост гифов и ограничивая инфицирование растений[8, стр.297].
Специальные исследования показали, что при взаимодействии бактерий и тканей хозяина различные ферменты деградации появляются не одновременно. Например, пектилметилэстераза присутствовала и в неинокулированных бактерией Erwiniacarotovorasubspatroseptia тканях клубней картофеля[8, стр.298], тогда как полигалактуроназная, пектатлиазная, целлюлазная, протеазная и ксиланазная активности появлялись соответственно через 10,14,16,19 и 22 ч после инокуляции.
Накопление патогениндуцированных белков и устойчивость к инфекции проявляются и в соседней неинфицированной ткани. Из этого следует, что защитные белки индуцируются подвижными веществами, которые образуются в местах инфекции и затем передвигаются в непораженные ткани листьев, вызывая в них эффект защиты.
Активный в качестве индуктора компонент не инактивируется протеазами, его положительный заряд и тепловая стабильность могут свидетельствовать, что он представляет собой маленький пептид или аминокислоту. Малые гликопептиды, присутствующие в инфицированных вирусом табачной мозаики листьях табака, могут отвечать за индукцию системной устойчивости растений [8, стр.298].
Уже отмечалось, что под влиянием инфекции наблюдаются лигнификация, суберинизация клеточных стенок, накопление в них гидроксипролиновых белков, каллозы, что создает дополнительный барьер для патогенов. Происходит также накопление оксигенированных производных ненасыщенных жирных кислот, летучих ароматических гексаналей, таннинов, О-хинонов [8, стр.287]. Так как избыточное образование всех этих соединений связано с активацией имеющихся в клетках или с синтезом новых ферментов, то не были неожиданными факты индукции экспрессии патогеном большого количества генов[8, стр.288].
По всей вероятности, круг защитных веществ может быть со временем значительно расширен.
Реакция растений на действие вирусов, так же как на патогенные грибы и бактерии, может варьировать между иммунностью и восприимчивостью [6, стр.328]. Большинство растений устойчиво к вирусной инфекции благодаря природной селекции в ходе эволюции. Интересно, что свойство устойчивости распространяется из инфицированных в непораженные клетки и ткани. Вирусреплицирующая способность растений может быть усилена некоторыми фитогормонами (кинетином, ИУК), полианионами (дрожжевой РНК, поли-И, поли-Ц, сополимером этиленмалеинового альдегида, полиакриловой кислотой). Устойчивость развивалась градуально и была чувствительна к актиномицину Д.
Изучению молекулярных основ взаимодействия патогенов и растения-хозяина, в том числе роли продуктов частичной деградации биополимеров и липидов в выработке устойчивости, в настоящее время уделяется все большее внимание. В дополнение к журналу, посвященному этому направлению исследований - "RhysiologicalandMolecularPlantPathology", недавно начал издаваться специальный журнал "MolecularPlant - MicrobeInteractions". Можно надеяться, что это приведет к подъему уровня работ не только в области молекулярной фитопатологии, но и в теории стресса в целом[8, стр.297].
4. ЗАКЛЮЧЕНИЕ
Уже давно внимание биологов и медиков привлекало интереснейшее явление: в результате действия экстремальных условий, изоляции или отмирания растительных или животных тканей образуются вещества, способные действовать на здоровые клетки и ткани, их рост и развитие, устойчивость и т.д. К ним можно отнести и "раневые гормоны" и "некрогормоны", существование которых было постулировано в начале нашего века Г. Габерляндтом [2, стр.122]. Не вызывает сомнений, что среди этих веществ имеются продукты распада сложных соединений. Некоторые из них способны выделяться в окружающую среду и оказывать действие на другие организмы в экосистемах[4, стр.197].
Рассмотрение особенностей катаболизма биополимеров и липидов в растениях в условиях стресса оказалось сложной задачей, причем не столько из-за избытка, сколько из-за недостаточности сведений, касающихся многих промежуточных этапов деградации этих соединений. Значительная часть литературы посвящена ферментам, катализирующим главным образом, стартовые реакции процессов катаболизма. Хорошо изучены и реакции дальнейшего превращения образующихся при деградации мономерных продуктов. В то же время весьма немногочисленны результаты исследований промежуточных (например, олигомерных) продуктов, их структуры, характера дальнейших превращений, биологической активности и механизма их действия на обмен веществ клеток, что объясняется как методическими трудностями, так и недостаточным вниманием физиологов и биохимиков растений к этой важной области метаболизма.
Можно считать доказанным, что интермедиаты катаболизма биополимеров и липидов играют большую роль в работе сложного регуляторного механизма клетки и в коррекции метаболизма в соответствии с изменившимися условиями, в частности в формировании ответной реакции клеток растений на действие различных стрессоров[4, стр.198].
Необходимо иметь в виду, что сведения о роли продуктов катаболизма в регуляции обмена веществ получены на основании изучения действия экзогенных соединений. Однако нельзя считать, что реакция клеток на эти соединения будет тождественна реакции на их появление или повышение содержания в том или ином компартменте внутри клеток. Более того, представляется вероятным, что клетка реагирует на многие экзогенные биополимеры и липиды и продукты их деградации как на "обломки кораблекрушения", как на сигналы тревоги, идущие от разрушенных или находящихся в состоянии сильного стресса соседних клеток. Эти органические соединения, взаимодействуя с поверхностью клеточной мембраны, вызывают ее включение в цепь усиления сигнала тревоги, состоящую из различных вторичных посредников (в том числе упоминавшихся в предыдущих разделах). По всей вероятности, на внешней стороне плазмаллемы имеются разнообразные рецепторы, с которыми способны связываться вышеупомянутые "обломки кораблекрушения". Можно предполагать, что это реликтовые формы конструкций, с помощью которых осуществлялось получение информации об изменениях гуморальной обстановки вокруг клетки. Некоторые из этих рецепторных конструкций совершенствовались в ходе эволюции многоклеточных высших растений и стали играть роль специализированных высокоэффективных рецепторов гормонов. За другими могла сохраниться изначальная функция взаимодействия с достаточно сложными органическими соединениями (индукторами защитных реакций, в особенности против патогенов), но само появление этих соединений в окружающей здоровую клетку среде приобрело значение неблагополучия, сигнала тревоги. Судя по имеющимся данным, клетка воспринимает различные экзогенные соединения как знак тревоги при значительно меньших концентрациях, по сравнению с теми же веществами эндогенного происхождения[4, стр.199].
Имеются основания считать, что растительные клетки обладают сходной системой коррекции метаболизма. Если принять это положение, то необходимо дополнить схему, приведенную на рис. 1 (отражающую роль промежуточных продуктов катаболизма как эндогенных регуляторов внутри тех клеток, где они образовались), фрагментом, объясняющим роль этих продуктов в качестве экзогенных эффекторов метаболизма для других клеток (рис. 4). Этот фрагмент включает взаимодействие с рецепторами клеточной мембраны и участие систем вторичных посредников в передаче сигналов в генетической аппарат.
В большинстве случаев имеется информация о действии на функции клеток лишь одного из промежуточных продуктов катаболизма, реже в сочетании с уже хорошо изученным "метаболическим репером", например ауксином. В то же время необходимо учитывать сложность влияния на обмен веществ клеток совокупности образующихся при катаболизме сигнальных молекул, изменения их набора или соотношения концентраций. Например, при изучении образования фитоалексинов растениями под влиянием патогенного гриба Phytophtorainfestans. Оказалось, что элиситорная активность продуцируемых грибом арахидоновой и эйкозапентаеновой кислот значительно повышалась в присутствии водорастворимых b-1,3-глюканов, тоже появляющихся в результате взаимодействия патогена и хозяина. Это позволяет говорить о взаимной корректировке (в данном случае усилении) биологического действия продуктов катаболизма различных классов биополимеров и липидов[4, стр.199].

Рис. 4.
Схема действия некоторых органических (сигнальных) веществ, освобождаемых поврежденной или находящейся в состоянии сильного стресса клеткой, на соседние клетки
В одном из своих выступлений академик А.Л. Курсанов сравнил проблему регуляции функций растений с горной вершиной, которую предстоит покорить и только у подножия которой находятся исследователи.
В настоящей работе предпринята попытка, используя данные, полученные из различной литературы, показать, что в покорении вершины регуляции имеется еще один перспективный путь - изучение промежуточных продуктов катаболизма биополимеров и липидов, их биологической активности и механизмов их действия на физиолого-биохимические процессы.
Разнообразие и высокая специфичность ферментов, отвечающих за реакции деградации биополимеров и липидов, обнаруживаемые в последнее время тонкие механизмы регуляции активности этих ферментов или ферментных систем, все большая информация о биологической активности образующихся соединений позволяют считать, что направленное исследование роли промежуточных продуктов катаболизма в регуляции анаболических процессов и связанных с ними функций растений Должно привести в ближайшие годы к новым интересным результатам.
Многие из этих физиологически активных продуктов найдут применение в сельском хозяйстве, медицине и биотехнологии как активаторы или ингибиторы в качестве самостоятельных препаратов или в комплексе с другими, уже применяющимися соединениями[4, стр.199].
Список литературы:
1. Войников В.К., Иванова Г.Г.
Физиологический стресс и регуляция активности генома клеток эукариотов // Успехи соврем, биологии. 1988. Т. 105, № 1. С. 3-14.
2. Дин Р.
Процессы распада в клетке. М.: Мир, 1981. 120 с.
3. Заленский О.В.
Эколого-физиологические аспекты изучения фотосинтеза. Л.: Наука, 1977. с.57-60 (Тимирязевские чтения).
4.Ильинская Л.И., Васюкова Н.И., Озерецковская О.Л.
Биохимия аспекты индуцированной устойчивости и восприимчивости растений М.: 1991 с. 197-199 (Итоги науки и техники. Защита растений; Т. 7).
5. Курсанов А.Л.
Взаимосвязь физиологических процессов в растении. М.: Издательство АН СССР, 1960. с.44-45 (20-е Тимирязев. чтение).
6.Одум Ю.
Экология. М.: Мир, 1986. Т. 1. 328 с.
7. Вознесенский В.Л.
Фотосинтез и дыхание растений в разных условиях среды // Фотосинтез и продукционный процесс / Под ред. А.А. Ничипоровича. М.: Наука, 1988. С. 132-137.
8. Блехман Г.И., Шеламова Н.А.
Синтез и распад макромолекул в условиях стресса // Успехи соврем, биологии. 1992. Т. 112, № 2. С. 281-297.
9. Бурлакова Е.Б., Хохлов А.П.
Изменение структуры и состава липидной фазы биологических мембран при действии синтетических антиоксидантов: Влияние на передачу информационного сигнала на клеточном уровне // Биол. мембраны. 1985. Т. 2, № 6. С. 557-562.
|