The production of 2
H-labeled amino acids by a new mutant of RuMP fucultative methylotroph
В
revibacterium methylicum
Oleg V. Mosin1
1
Department of Biotechnology, M. V. Lomonosov State Academy of Fine Chemical Technology, Vernadskogo Prospekt 86, 117571, Moscow, Russia
Summary
The biosynthesis of 2
H-labeled phenylalanine was done by converse of low molecular weight substrates ([U- 2
H]methanol and 2
H2
O) in a new RuMP facultative methylotrophic mutant Brevibacterium methylicum. To make the process work, adapted cells with improved growth characteristics were used on minimal medium M9 with the maximum content of 2
H-labeled substrates. Alanine, valine, and leucine/isoleucine were produced and accumulated exogeneously in
addition to the main product of biosynthesis. Electron impact mass spectrometry of methyl esters of the N
-Dns-amino acid mixture obtained after the chemical derivatization of growth medium with dansyl chloride and diazomethane, was done to calculate the deuterium enrichment of the amino acids synthesized. The experimental data testified to the character of labeling of amino acid molecules as heterogeneous; however, high levels of deuterium enrichment were detected in all presented molecules - for phenylalanine the enrichment was six, leucine/isoleucine - 5.1, valine - 4.7, and alanine - 3.1 deuterium atoms.
Keywords:
Brevibacterium methylicum - Heavy water - Biosynthesis - 2
H-Labeled amino acids - Phenylalanine - EIMS
Abbreviations:
EI MS: electron impact mass spectrometry; TLC: thin layer chromatography; DNSCl: dansylchloride; DZM: diazomethane; N-NMU: N-nitroso-methylurea; RuMP: rybolose monophosphate; PenP: pentose phosphate; PEP: phosphoenolpyruvate; ERP: erythrose-4-phosphate.
Introduction
Labeling of amino acid molecules with deuterium is becoming an essential part for various biochemical studies with 2
H-labeled molecules and investigation of certain aspects of their biosynthesis(LeMaster, 1990).
For introduction of deuterium into amino acid molecules either chemical or biosynthetical methods may be used. Chemical synthesis of these compounds has one significant limitation; it is a very laborious and costly multistep process resulting in a mixture of dl
-racemates. This major disadvantage, however essentially delaying its development is a difficulty in preparing the appropriate 2
H-labeled amino acids. Chemical synthesis usually results in obtaining a mixture of d,l
-racemates (Daub, 1979). Although chemomicrobiological synthesis overcomes this problem (Walker, 1986), the amount of purified enzymes required is prohibitive (Faleev, 1989). By growing algae on media with 96% (v/v) 2
H2
O, the desired 2
H-labeled biochemicals can be produced both at high yields and enrichments (Cox, 1988), but the process involves algae is limited by the expense of a mixture of 2
H-labeled amino acids isolated from hydrolysates of biomass (Daboll, 1962). The using for this purpose a certain methylotrophs which assimilates MetOH as a source of carbon and energy via RuMP cycle has a great practical advantage because their ability to produce and acumulate a gram quantities of 2
H-labeled amino acids during the growth on media with 2
H2
O and [U -2
H]MetOH and the comparatively low price of [U -2
H]MetOH (Karnaukhova, 1994).
The biosynthesis of 2
H-labeled amino acids usually involves growth of an organism on selective media containing the labeled substrates:e.g
., growth of algae autotrophically on media with content of 2
H2
O 90% and more, is a well established method for biosynthesis of numerous highly deuterated molecules. But this method, while being generally applicable, is limited by the low resistance of plant cells to 2
H2
O and expense of 2
H-labeled amino acids isolated from algae hydrolysates. Alternative and relatively inexpensive objects for biosynthesis of 2
H-labeled amino acids seem certain auxotrophic mutants of methylotrophic bacteria using methanol as a main source of carbon and energy via
the ribulose-5-monophosphate (RuMP) and the serine cycle of carbon assimilation. These bacteria have a big advantage because of their ability to produce and accumulate gram quantities of highly enriched, 2
H-labeled amino acids during growth on minimal salt media with [U- 2
H]methanol and 2
H2
O and the comparatively low price of [U- 2
H]methanol. It is only in recent years that some progress was made in the isolation of a number of versatile the RuMP cycle methylotrophic bacteria, suitable for such studies, though the research that has been done with methylotrophs was limited and suffered from low growth characteristics on 2
H2
O-containing media. Although the production of 2
H-labeled amino acids by obligate methylotroph Methylobacillus flagellatum
described by Karnaukhova involves the growth on media with approximately 75% (v/v) 2
H2
O. We have recently selected a new mutant of facultative methylotroph Brevibacterium methylicum
, realizing the NAD+
dependent methanol gehydrogenase (EC 1.6.99.3) variant of RuMP cycle of carbon assimilation, which seems more convinient for the preparation of 2
H-labeled amino acids than M. flagellatum
because its ability to grow on liquid M9 with 98% (v/v) 2
H2
O (Mosin, 1995).
Thus, we have previously studied the applicability of the RuMP cycle obligate methylotrophic bacterium Methylobacillus flagellatum
for biosynthesis of 2
H-labeled leucine 8)
. This approach is not yet practical for the biosynthesis of 2
H-labeled phenylalanine, mainly because of the absence of suitable methylotrophic producer of this amino acid. After selecting a new the RuMP cycle methylotrophic producer of phenylalanine, leucine auxotroph Brevibacterium methylicum
, we have used this strain for this research.
Material and methods
2
H2
O (99.9 at.% 2
H) was purchased from Russian Scientific Enterprises, Sankt Petersburg. [U -2
H]MetOH (97.5 at.% 2
H) was from Biophysic Center, Pushino. DNSCl of sequential grade was from Sigma Chemicals Corp., USA. DZM was prepared from N-NMU, Pierce Chemicals, Corp., USA. A gram-positive parental strain of RuMP facultative methylotroph Brevibacterium methylicum
# 5662 was obtained from Russian State Scientific Center for Genetics and Selection of Industrial Microorganisms GNIIGENETIKA (Nesvera, 1991).
Basal salt medium M9 (Miller, 1976) with MetOH as a carbon and energy source (2%, v/v) and supplemented with Leu (100 mg/l) was used for bacterial growth. For isotopic experiments M9 was enriched with [U -2
H]MetOH and 2
H2
O of various content (see Table below). The bacterial growth was carried out under batch conditions (Karnaukhova, 1994). The exponentially growing cells (cell density 2.0 at absorbance 540 nm) were pelleted by centrifugation (1200 g for 15 min), the supernatant was lyophilized and used for chemical derivatization.
The amount of Phe was determined for 10 ml aliquotes of liquid M9 by TLC with solvent of iso-PrOH-ammonia (7:3, v/v) using pure commercial available Phe as a standard. The spots were detected by 0.1% ninhydrine solution in acetone, eluted by 0.5% CdCl2
solution in 50% EtOH (2 ml). The absorbance of the eluates was measured at 540 nm, the concentration was calculated using a standard curve.
The samples of lyophilized M9 were dansylated in 1 M sodium hydrohycarbonate-acetone (1:2, v/v) solution (pH 10-11) with tenfold excess of DNSCl, and treated according to Devenyi (1976). The derivatization to methyl esters of N-DNS-amino acids was performed in a standard procedure with DZM (Greenstein, 1976).
EI MS was performed on Hitachi MB 80 spectrometer at ionizing energy 70 eV and an ion source temperature of 180oC.
Results and discussion
Phe is synthesized in most bacteria via shikimic acid pathway (Conn, 1986). The precursors for the biosynthesis of Phe are PEP and ERP. The latter compound is an intermediate in the PenP pathway and, in some methylotrophs, the RuMP cycle of carbon assimilation (Antony, 1982; Kletsova, 1988). It is widely accepted, that the native bacterial strains can not to be a strong producers of Phe owing to the effective mechanisms of its metabolic regulation, although certain bacterial mutants with mutations of prephenate dehydrogenase (EC 1.3.1.12), prephenate hydratase (EC 4.2.1.51), chorismate mutase (EC 5.4.99.5) and a number of other several enzymes are proved to be an active producers of this amino acid (Umbarger, 1978). That is why the best Phe producing strains once selected were the mutants partially or completely dependent on Tyr or Trp for growth. The reports about the other regulative mechanisms of Phe biosynthesis in bacterial cell are quite uncommon, though today it is known a large number of RuMP cycle auxotrophic mutants of methylotrophs, covering numerious steps in aromatic amino acid biosynthesis (Dijkhuizen, 1996). The selection of new producers of Phe has a big importance for studies of their regulating pathways and possible production of 2
H-labeled Phe.
For our studies we have used a new non-traditional producer of phenylalanine: a leucine auxotroph of the facultative methylotrophic bacterium Brevibacterium methylicum
obtaining the NAD+
dependent methanol dehydrogenase (EC 1.6.99.3) variant of the RuMP cycle of carbon assimilation, with maximum productivity of phenylalanine on protonated medium M9 - 0.95 gram per liter of growth medium. According to experiments, various compositions of [U- 2
H]methanol and 2
H2
O were added to the growth media as hydrogen (deuterium) atoms could be assimilated both from methanol and H2
O. The growth characteristics of the non-adapted bacteria and production ofphenylalanine in the presence of increasing content of 2
H2
O are given in Table (Expts. 3-10) relative to the control (1) on protonated medium and to the adapted bacteria (Expt. 10’). The odd numbers of experiment were chosen to investigate whether the replacement of [U -2
H]methanol of its protonated analogue has a positive effect on growth characteristics in the presence of 2
H2
O. The maximum deuterium content was reached under conditions (10) and (10’) in which we used 98% (v/v) 2
H2
O and 2% (v/v) [U -2
H]methanol. In the control, the duration of a lag-
phase did not exeed twenty hours, the yield of microbial biomass (wet weight) and production of phenylalanine were 150 and 0.95 gram per 1 liter of growth medium (see relative values in Table, Expt. 1). The results suggested, that below 49% (v/v) of 2
H2
O (Table, Expts. 2-4) there was a small inhibition of growth indicators compared with the control (1), above 49% (v/v) of 2
H2
O (Table, Expts. 5-8), however growth was markedly reduced, while at the upper content of 2
H2
O (Table, Expts. 9-10) growth was extremely small. With increasing content of 2
H2
O in the media there was a simultaneous increase both of the lag-phase and generation time. Thus, under experimental conditions (10) where we used 98% (v/v) 2
H2
O and 2% (v/v) [U -2
H]methanol, the lag-phase was more than three and the generation time - 2.2 times that on ordinary protonated medium (1). The production of phenylalanine and yield of biomass were decreased on medium with 98% (v/v) 2
H2
O and 2% (v/v) [U -2
H]methanol by 2.7 and 3.3 times respectively; in contrast to the adapted bacteria (10’), the growth characteristics of non-adapted bacteria on maximally deuterated medium were strongly inhibited (Table, Expt. 10). The replacement of protonated methanol by [U- 2
H]methanol caused small alterations in growth characteristics (Table, Expts. 2, 4, 6, 8, 10) relative to experiments, where we used protonated methanol (Table, Expts. 3, 5, 7, 9).
Table.
Isotope components of growth media and characteristics of bacterial growth
  |
Media components, % (v/v)
H2
O 2
H2
O MetOH [U -2
H]
MetOH
|
Lag-
phase (h) |
Yield of
biomass (%)
|
Generation time (h) |
Production of phenylalanine (%) |
| (a) |
98 |
0 |
2 |
0 |
20 |
100.0 |
2.2 |
100.0 |
| (b) |
73.5 |
24.5 |
0 |
2 |
34 |
85.9 |
2.6 |
97.1 |
| (c) |
49.0 |
49.0 |
0 |
2 |
44 |
60.5 |
3.2 |
98.8 |
| (d) |
24.5 |
73.5 |
0 |
2 |
49 |
47.2 |
3.8 |
87.6 |
| (e) |
0 |
98.0 |
0 |
2 |
60 |
30.1 |
4.9 |
37.0 |
The production of L-phenylalanine was linear with respect to the time up to exponentaly growth cells (see Fig.1). During the fermentation the formation rate of L-phenylalanine was about 5 mmol/day. As shown in Fig. 1, the substitution by deuterium atoms pronons of water and methanol caused the decreasing both the production of L-phenylalanine and the yieald of biomass. Hawever, the decreasing of L-phenylalanine production (up to 0,5 g\L) was observed in those experiments (10) (Fig.1) when using non adapted cells on media with 98 % (v/v) 2
H2
O. The growth rate and generation time for adapted cells were found to be the same as in control in ordinary water despite of small increasing of lag-phase. In contrast to adapted cells, the growth of non-adapted species on maximal deuterated media was strongly inhibited by deuterium. These data are shown in Fig. 2.
A smart attempt was made to intensificate the growth and biosynthetic parameteres of cells to grow on media containing highly concentration of deuterated substrates. We employed a "step by step" adaptation method, combined with the selection of clones resistent to deuterium using agaric media supplemented with C2
H3
O2
H 2% (v/v) and with increasing 2
H2
O content starting from pure water up to 98 % (v/v) 2
H2
O. The degree of cell survive on maximum deuterated medium (10), containing 98 v/v.% 2
H2
O and 2 v/v.% C2
H3
O2
H was about 40%. Figure 1 shows characteristic growth and biosynthesis curves for adapted to 2
H2
O (10’’
) and non-adapted (10) cells in conditions compared with the control (1) in H2
O. The transfer of fully deuterated cells to ordinary protonated medium results eventually in normal growth.
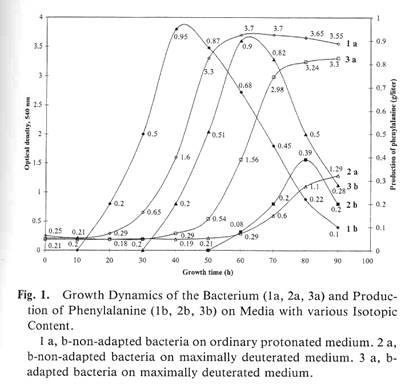
The results on adaptation testified, that the generation time for adapted bacteria was approximately the same as in the control (1) despite the two-fold increase of the lag-phase (Table, Expt. 10’). The yield of microbial biomass and level of phenylalanine production for adapted bacteria on maximally deuterated medium (Table, Expt. 10’) were decreased relative to the control (1) by 13 and 5.3% respectively. Figure 1 shows growth (Expts. 1a, 2 a, 3 a) and production of phenylalanine (Expts. 1 b, 2 b, 3 b) for non-adapted (2) and adapted (3) bacteria on maximally deuterated medium under conditions like the control (1) on protonated medium. As shown from Fig. 1, the curves of phenylalanine production were close to a linear extrapolation with respect to the exponential phase of growth dynamics. The level of phenylalanine production of non-adapted bacteria on maximally deuterated medium was 0.39 g/liter after 80 hours of growth (Fig. 1, Expt. 2 b). The level of phenylalanine production for adapted bacteria under those growth conditions was 0.9 g/liter (Fig. 1, Expt. 3 b). Thus, the use of adapted bacteria in growth conditions to be the same as in the control (1), enabled us to improve the level of phenylalanine production on maximally deuterated medium by 2.3 times. However, phenylalanine is not the only product of biosynthesis; other metabolically related amino acids (alanine, valine, andleucine/isoleucine) were also produced and accumulated in the growth medium in amounts of 5-6 mmol in addition to phenylalanine. This fact required, for the future prospects of the production of labeling molecules of amino acids with deuterium, an efficient separation of 2
H-labeled phenylalanine from other amino acids of growth medium. Recently such separation was done using a reversed-phase HPLC method developed for methyl esters of N
-Dns- and Bzl-amino acids with chromatographic purity of 96-98 and yield of 67-89%.
For evaluation of deuterium enrichment methyl esters of N-DNS-amino acids were applied because the peaks of molecular ions (M+
) allow to monitor the enrichment of multicomponential mixtures of amino acids being in composition with growth media metabolites, therefore EI MS allows to detect samples with amino acids of 10-9
-10-12
mol (Karnaukhova, 1994). N-DNS-amino acids were obtained through the derivatization of lyophilized M9 with DNSCl. To increase the volatality of N-DNS-amino acids, the methylation with DZM was made to prevent the possible isotopic (1
H-2
H) exchange in molecule of Phe. With DZM treatment it occured the derivatization on aNH2
group in the molecule, so that its N-methylated derivative was formed to the addition of methyl ester of N-DNS-Phe.
Mass spectra EI MS of methyl esters of N-DNS-amino acid mixtures, obtained from M9 where used 0; 73.5 and 98% (v/v) of 2
H2
O (Table, Expts. (a), (d), (e)) are shown in consecutive order in Figs. 1-3. The fragmentation pathways of methyl esters ofN-Dns-amino acids by EI MS leads to the formation of the molecular ions (M+
) from whom the fragments with smaller m/z
ratio further are formed. Since the value of (M+
) for Leu is as the same as for Ile, these two amino acids could not be clearly estimated by EI MS. A right region of mass spectra EI MS contains four peaks of molecular ions (M+
) of methyl esters of N-DNS-amino acids: Phe with m/z
412; Leu/Ile with m/z
378.5; Val with m/z
364.5; Ala with m/z
336.4 (see Fig. 1 as an example). A high continuous left background region at m/z
80 - 311 is associated with the numerious peaks of concominant metabolites and fragments of further decay of methyl esters of N-DNS-amino acids.
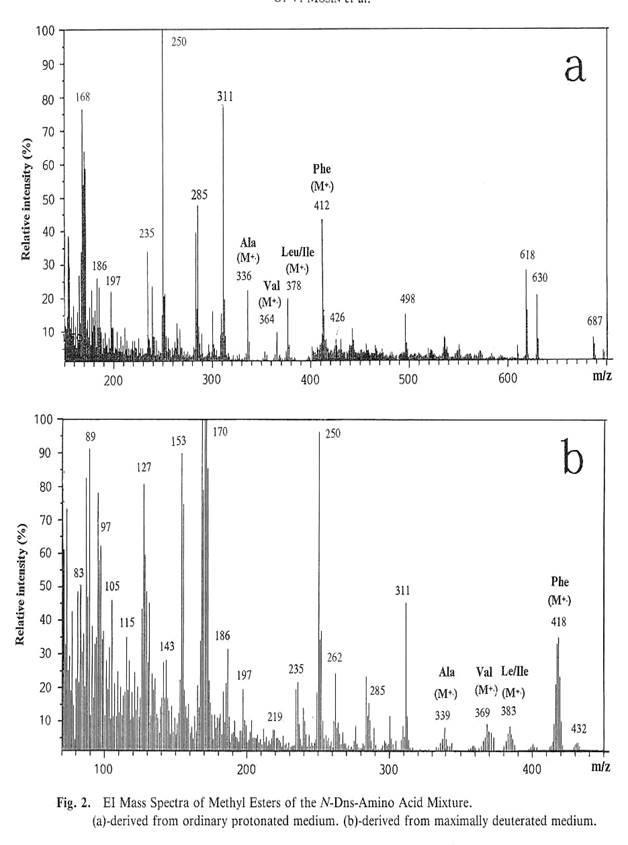
The results, firmly established the labeling of amino acids as heterogenious, juging by the presence of clasters of adduct peaks at their molecular ions (M+
); the species of molecules with different numbers of deuterium atoms were visualised. The most aboundant peak (M+
)in each claster was registered by mass spectrometer as a peak with average m/z
ratio, from whom the enrichment of each individual amino acid was calculated. Thus, in experiment (d) shown in Fig. 2 where used 73.5% (v/v) 2
H2
O the enrichment of Phe was 4.1, calculated at (M+
) with m/z
416.1 (instead of m/z
412 (M+
) for non-labeled compound); Leu/Ile - 4.6 (M+
) with m/z
383.1 instead of m/z
with 378.5 (M+
)); Val - 3.5 (M+
with m/z
368 instead of m/z
(M+
) with 364.5); Ala - 2.5 deuterium atoms ((M+
)with m/z
338.9 instead of m/z
with 336.4 (M+
)).
With increasing of 2
H2
O content in liquid M9, the levels of amino acid enrichment varried propotionaly. As seen in Fig. 3 in experiment (e) where used 98% (v/v) 2
H2
O the enrichment of Phe was six ((M+
) with m/z
418 instead of m/z
412 (M+
)); Leu/Ile - 5.1 ((M+
) with m/z
383.6 instead of m/z
with 378.5 (M+
)); Val - 4.7 ((M+
) with m/z
369.2 instead of m/z
(M+
) with 364.5); Ala - 3.1 deuterium atoms (M+
) with m/z
339.5 instead of m/z
with 336.4 (M+
)). The label was distributed uniformely among the amino acid molecules, in experiment (e) the enrichment of 2
H-labeled amino acids was nevertheless less than we estimated theoretically, because Leu was added in growth medium in protonated form. This effect should be seriously scrutinised before the applying this mutant for the preparation of 2
H-labeled amino acids.
References
1. Antony C (1982) Bacterial Oxidation of Methane and Methanol, The Biochemistry of Methylotrophs, 2 nd edn. Academic Press, London, p 78
2. Boer L de, Harder W, Dijkhuizen L (1988) Phenylalanine and Tyrosine Metabolism in the Facultative Methylotroph Nocardia sp. 239
. Arch Microbiol 149: 459-465
3. Conn EE (1986) Recent Advances in Phytochemistry, The Shikimic Acid Pathway, 2 nd edn. Plenum Press, New York, p 20
4. Cox J, Kyli D, Radmer (1988) Stable Isotope Labeled Biochemicals from Microalgae. Trends Biotechnol 6: 279-282
5. Daboll HF, Crespi HL, Katz JJ (1962) Mass Cultivation of Algae in Pure Heavy Water. Biotechnol and Bioengineering 4: 281-297
6. Daub GM (1979) Syntheses with Stable Isotopes. In: Daub GM Stable Isotopes. Proceedings of the 3d Inter Conf, 1979, Academic Press, NY 1979, pp 31-44
7. Devenyi T, Gergely J (1976) Amino Acids, peptides and proteins, 3 d edn. Mir Publishers, Moscow, p 281
8. Dijkhuizen L (1996) Metabolic Regulation in the Actinomycete Amycolatopsi Methanolica
, a Facultative Methylotroph Employing the RuMP Cycle for Formaldehyde Assimilation. Microbial Growth on C1 Compounds Kluwer Academic Publishers, London, pp 9-15
9. Faleev NG, Ruvinov SB, Saporovskaya MB, Belikov VM, Zakomyrdina LN (1989) Preparation of a-Deuterated Amino Acids by E. coli
Cells Containing Tryptophanase. Izv Akad Nauk USSR Ser Khim (in Russian) 10: 2341-2343
10. Greenstein JP, Winitz M (1976) Chemistry of amino acids, 3 d edn. Mir Publishers, Moscow, p 396
Karnaukhova EN, Reshetova OS, Semenov SY, Skladnev DA, Tsygankov YD (1994) 2
H- and 13
C-Labeled Amino Acids Generated by Obligate Methylotrophs Biosynthesis and MS Monitoring. Amino Acids 6:
165-176
11. Kletsova LV, Chibisova ES, Tsygankov YD (1988) Mutants of the Oligate Methylotroph Methylobacillus flagellatum
KT Defective in Genes of the Ribulose Monophosphate Cycle of Formaldehyde fixation. Arch Microbiol 149: 441-446
12. LeMaster DM (1990) Deuterium Labeling in NMR Structural Analysis of Larger Proteins. Quart Revs Biophys 23: 133-174
13. Miller JH (1976) Experiments in Molecular Genetics. Cold Spring Harbor Laboratory Cold Spring Harbor, New York p 393
14. Mosin OV, Skladnev DA, Tsygankov YD Russian Patent 93055824/13 (Nov. 17, 1995)
15. Nesvera J, Patek M, Hochmannova J, Chibisova E, Serebrijski I, Tsygankov Y, Netrusov A (1991) Transformation of a New Gram-Positive Methylotroph, Brevibacterium
methylicum
, by plasmid DNA. Appl Microbiol Biotechnol 35: 777-780
16. Umbarger HE (1978) Amino Acid Biosynthesis and its regulation. Ann Rev Biochim 47, 533-606
17. Walker TE, Matheny C (1986) An Efficient Chemomicrobiological Synthesis of Stable Isotope-Labeled L-Tyrosine and L-Phenylalanine. J Org Chem 51: 1175-1179.
|