Введение
Аквакультура в мире в начале XXI века характеризуется прогрессирующим ростом. Это происходит на фоне резкого падения запасов естественных популяций рыб в связи с их сверхинтенсивным промышленным и браконьерским ловом, разделом Каспия на отдельные территории суверенных государств.
С введением новых правил рыболовства на Каспии с 1962 г. промысел рыбы был полностью перебазирован из прибрежья моря в реки. При этом в западно-каспийском районе, в том числе и в реках, промысел осетровых был полностью прекращен. Эта мера сыграла исключительно важную роль в сохранении молоди ценных промысловых рыб, особенно осетровых, восстановлению их запасов, в существенной мере подорванных к этому времени.
Концепцией развития рыбного хозяйства на Каспии, принятой в 50-е годы XX века, было предусмотрено развитие отрасли по трем основным направлениям:
Главное направление — пастбищное осетровое хозяйство — в основном должно было базироваться на искусственном разведении молоди на специализированных осетровых рыбоводных заводах и выпуске ее в море на нагул.
Второе направление — это осуществление комплекса рыбоводно-мелиоративных мероприятий по поддержанию запасов полупроходных видов рыб, размножающихся в дельтах рек.
Третье направление – развитие пресноводной аквакультуры на основе прудового рыбоводства.
Главной проблемой каспийского рыбного хозяйства в настоящее время является критическое состояние запасов осетровых, вызванное масштабным незаконным выловом во всех регионах моря.
По данным правительства Р.Ф. основными целями и задачами развития аквакультуры:
- усовершенствовать и укрепить нормативно-правовые основы для государственного регулирования в области аквакультуры;
- активизировать капиталоемкие научные исследования в области аква- и марикультуры, модернизировать производственные фонды (прибрежную технологическую базу, суда и орудия лова); усилить приоритетное значение аквакультуры в рамках самостоятельного национального проекта или подпроекта в национальном проекте «Развитие агропромышленного комплекса» для оказания всех форм государственной поддержки;
- обновить и реконструировать существующие производственные фонды предприятий аквакультуры.
Для обеспечения душевого потребления рыбы в соответствии с потребительской корзиной в 2015 г. необходимо произвести пищевой рыбной продукции в объеме 2,15 млн. тонн, а при рациональной норме - 2,58 млн. тонн. В настоящее время вылов рыбы в РФ стабилизировался на уровне 3,2 млн. тонн, из которых для пищевых целей используется 1,3-1,5 млн. тонн. Следовательно, производство рыбы в аквакультуре должно составить не менее 1,1 млн. тонн. Достижение указанных объемов возможно только при существенном расширении, модернизации и реконструкции материально-технической базы, что подразумевает придание аквакультуре статуса национального приоритета с обеспечением государственной поддержки и финансирования.
По имеющимся оценкам, для реализации проекта развития аквакультуры РФ на период 2007-2011 гг. требуется финансирование в объеме 91,2 млрд. рублей (примерно 3,50 млрд. долл. США), в том числе 43,2 млрд. рублей (1,65 млрд. долл. США) госбюджетное финансирование и местные бюджеты и 48,0 млрд. рублей (1,85 млрд. долл. США) - внебюджетные средства.
Ориентиры развития аквакультуры в РФ к 2020 г. включают: производство товарной рыбы в аквакультуре - 1,4 млн. тонн и марикультуре-400 тыс. тонн. Выпуск личинок и молоди ценных видов рыб в естественные водоемы к 2020 г. достигнет 80-100 млрд. экземпляров.
Экономический эффект от реализации на российском рынке продукции аквакультуры составит 25 млн. рублей (около 1 млн. долл. США) на 1 тыс. тонн. Аквакультура позволит создать в РФ до 100 тыс. рабочих мест и бупет способствовать расширению занятости в российских региона.
По данным Каспнирха за 2005 г. в Каспийском море выполнены сезонные (весна, лето, осень) тралово-акустическис и сетные съемки по изучению распределения, качественного состава и численности осетровых. Весенняя и осенняя съемки проводились на акватории западной части Северного Каспия. За период исследований проведено 581 траление 24,7- и 9-метровыми тралами и 139 постановок сетных порядков с размером ячеи от 28 до 180 мм. отловлено и проанализировано 1602 экз. осетровых: русского и персидского осетров - 1400 экз.. севрюги — 168 экз.. белуги - 32 экз.. шипа - 2 экз. В весенний период за 37 тралений и 29 постановок сетей было выловлено 4 экз. белуги. В траловых уловах ее отмечали на свале глубин Северного и Среднего Каспия. Диапазон глубин в местах лова изменялся от 4,0 до 26,0 м, температура воды - от 6,1 до 14,2 °С. Улов состоял из особей непромысловой длины. Средние размерно-весовые характеристики белуги в весенний период - 149,5 см и 16,5 кг соответственно. Высокие коэффициенты упитанности (у самцов и самок - 0,66) и зрелости (у самцов - 3.14 %; самок - 6.25 %) свидетельствовали о благоприятных условиях нагула. По результатам летней учетной съемки за 383 траления выловлено 10 экз. белуги, сетными порядками (17 постановок) - 8 экз. Единичные траловые уловы белуги отмечены почти во всех частях Каспийского моря.
Качественная структура белуги в различных частях моря (северная, средняя, южная) не отличалась большим разнообразием. В северной части Каспия (приглубая зона) доля молоди в улове составила 100 %. Длина рыб варьировала от 145,0 до 182,0 см, масса - 21,0-42,0 кг. средние показатели - 181,5 см и 4 ] ,5 кг. В средней части моря длина составила 171,0-295,0 см, масса - 42,0-175,0 кг, средние значения -232,0 см и 90,25 кг, доля молоди в этом районе не превышала 50,0 %. В южной части моря ее доля возросла до 68,4 %, а средние показатели длины и массы снизились до 144,7 см и 30,0 кг. Таким образом, прослеживается закономерность в распределении молоди и взрослых рыб по акватории моря: молодые особи нагуливаются в основном в северной и южной частях моря, а старшевозрастные группы рыб осваивают для нагула Средний Каспий. Средняя длина белуги на всей акватории Каспийского моря - 193,25 см, масса- 55,5 кг: особи промысловых размеров имели длину 214,4 см, массу - 83,1 кг, неполовозрелые особи - 158,0 см и 28,0 кг. Высокий коэффициент упитанности самцов и самок белуги (0,66-0,79) отмечен во всех районах моря, что подтверждает удовлетворительные условия нагула вида.
Данные осенней тралово-сетной съемки в западной части Северного Каспия, как и весенней съемки, свидетельствуют о единичном присутствии в траловых и сетных уловах белуги: за 72 траления и 13 постановок сетей отловлено всего 6 экз. непромысловых размеров (Журавлёва,Ходоревская, Левин и др., 2005 ).
По данным Каспнирха в 2004 г. выпуск молоди осетровых 6-ю рыбоводными заводами Севкаспрыбвода не превысил 62 % от запланированной численности, что составило 33,4 млн шт., в т. ч. .1,5 млн шт. белуги, 25,1 млн шт. осетра и 6,8 млн шт. севрюги.
Численность молоди белуги составила лишь 4,5 % от общего объема выпуска. Следует отметить, что часть молоди выращивалась при плотностях посадки в 2-3 раза ниже норматива (110 тыс. шт./ га): к примеру, на Сергиевском заводе с плотностью 30 тыс. шт./га, на Лебяжьем - 45 тыс. шт./га. Снижение плотности посадки создает более благоприятные условия питания, обеспечивающие увеличение средней массы за нормативные сроки до 5,0-6,0 г. Из рис. 1, представляющего гистограммы массы и длины этой молоди, следует, что в прудах преобладали мальки массой 3,0-4,5 г (56%). Количество молоди массой от 0,4 до 2.9 г, т. е. менее установленного стандарта (3,0 г), оказалось значительно ниже среднемноголетних значений для этого вида и составило 21% общей выборки. Достаточно многочисленной оказалась группа массой 5,0-5,5 г (17,2%). Пик-распределения длины тела белуги был у мальков от 8,5 до 10,5 см. (Григорьева, Михайлова, Крупий и др., 2004).
Глава 1. Биологическая характеристика белуги в связи со средой обитания и образом жизни
1.1 Общая биология вида
Тип:
Хордовые (Chordata)
Под тип:
Черепные (Craniata)
Над класс:
Челюсноротые (Gnathostomata)
Класс:
Рыбы (Pisces)
Над отряд:
Хрящевые ганоиды (Chondrostei)
Семейство:
Осетровые (Acipenseridae)
Род:
Белуги (Huso)
Вид:
Белуга (Husohuso)
Тело массивное, толстое (huso в переводе с латинского – свинья). Рыло короткое тупое. Рот полулунной формы, большой, но, в отличие от Калуги, не переходящий на бока головы. В спинном плавнике 48–81 лучей, в анальном – 22–41. Спинных жучек – 9–17, боковых – 37–53, брюшных – 7–14. В спинном ряду первая жучка наименьшая. Жаберных тычинок 17–36. Спина и бока тела серовато-темные, брюхо белое. Белуга (рис. 1) относится к 120-хромосомной группе осетров (Витвицкая, Тихомиров, Егоров, 2002).

Рис. 1. Белуга
Распространение. Бассейны Черного, Азовского, Каспийского и Адриатического морей. До зарегулирования стока очень высоко поднималась по рекам. По Волге доходила почти до верховьев, встречаясь в Оке, Шексне, Каме, Суре и других притоках. По археологическим материалам вылавливалась даже в р. Москве. В настоящее время ареал ограничен нижними плотинами ГЭС. В Азовском море практически полностью исчезла, раньше, до постройки плотин, в Кубани поднималась до станицы Ладожская и, возможно, выше, в Дону – до Воронежской области. На рисунке 2 обозначены утраченные места обитания, крестиками – места археологических находок остатков белуги (Решетников, 2002).

Рис. 2. Ареал обитания белуги (+ – места археологических находок остатков белуги, черная штриховка – ареал обитания белуги.)
Возраст и размеры. Одна из крупнейших проходных рыб. В прошлом достигала длины более 5 м и массы более 1000 кг. Продолжительность жизни таких крупных экземпляров, видимо, превышала 100 лет. В 1970 г. в промысловых уловах на Волге средняя длина самок белуги составляла 267 см и масса 142 кг, самцов соответственно – 221 см и 81 кг. (Решетников, 2002).
Образ жизни. Проходная рыба. В большинстве крупных рек имеет озимую и яровую формы. В Волгу входит в сентябре-октябре (озимая форма) и в марте-апреле (яровая форма). В этой реке преобладает озимая, зимующая на ямах, раса. В Урале, наоборот около 70% мигрирующих рыб составляет яровая раса, нерестящаяся в год захода в реку. Период размножения приходится на апрель-май. Нерест проходит на пике паводка и начинается при температуре воды 6–7 °С. Оптимальными температурами являются 9–17 °С. Икра откладывается на глубоких местах (от 4 до 12–15 м) с быстрым течением, на каменистых грядах и галечных россыпях. Плодовитость в зависимости от размеров самок колеблется от 200 тыс. до 8 млн. икринок. Икринки крупные, у волжской белуги их диаметр варьирует от 3,6 до 4,3 мм и масса от 26 до 36 мг. Продолжительность эмбрионального периода при температуре воды 11–12 °С составляет в среднем около 200 ч. Молодь белуги и взрослые рыбы после нереста в реке не задерживаются и скатываются в море. Половозрелость у белуги наступает очень поздно: у самцов не ранее 12–14 лет, у самок – к 16–18 годам. Нерест не ежегодный. Белуга очень рано переходит на хищное питание. В Каспийском море ее основными кормовыми объектами являются вобла, судак, сазан, лещ, кильки и другие рыбы. Белуга охотно поедает собственную молодь и других осетровых. В Черном море в ее желудках встречаются в основном хамса и бычки. Молодь белуги длиной до 5 см питается придонными беспозвоночными – мизидами, гаммаридами, олигохетами и др. (Бабушкин, 1953).
Статус вида. Ценнейшая промысловая рыба. Основным бассейном ее добычи всегда был Каспий. Еще в начале 80-х годов прошлого столетия ее здесь вылавливали от 1,6 до 2,0 тыс. т. В результате зарегулирования стока рек практически полностью прекратилось естественное воспроизводство этого вида, и ее запасы целиком поддерживались за счет искусственного разведения на рыбозаводах. В последние годы, особенно после распада СССР, численность и уловы белуги катастрофически снижаются. Белуга, как вид, находящийся под угрозой исчезновения, внесена в "Красную книгу МСОП", а азовский подвид белуги – в "Красную книгу Российской Федерации" (Решетников, 2002).
1.2 Эмбриональное развитие
Эмбриональный период развития осетровых рыб состоит из 5 этапов в 36 стадий:
1 этап – оводнение икринки и появление бластодиска (1–3 стадии).
2 этап – дробление бластодиска до бластулы (4–12 стадии).
3 этап – образование зародышевых пластов – гаструляция (13–18 стадии).
4 этап – дифференциация зародышевых пластов на зачатки основных органов (19–28 стадий).
5 этап – развитие зародышей от начала пульсации сердца до вылупления (29–36 стадии).
1. Этап (1-3 ст.)
Оводнение икринки и появление бластодиска. Выметанные в воду и тут же осемененные яйца опускаются на дно, заносятся под камни и вскоре приклеиваются студенистой оболочкой к грунту. Оплодотворение вызывает кортикальную реакцию, и между цитоплазматической мембраной яйца и оболочкой возникает узкая (в несколько мкм) прослойка выделенною материала кортикальных гранул.
Яйцо внутри высвобождается от оболочек и поворачивается соответственно положению центра тяжести, вегетативным более тяжелым полюсом вниз, а анимальным вверх. В это время в щелевидное пространство под оболочкой выделяется гидрофильный коллоид из лакун на анимальном полюсе. Коллоид, привлекая воду из окружающей среды, набухает, и возникает периветеллиновое пространство. В яйце происходит перемещение пигмента. Он стягивается к центру анимального полюса и светлое пятно исчезает. Вскоре по краю анимальной области появляется светлая полоса – светлый серп.
В это время значительно возрастает прочность яйцевых, оболочек. Увеличение прочности оболочек связано с условиями развития зародышей под камнями на течении, где икра подвергается ударом камешков, песка и других предметов. Как только кортикальная реакция осуществится и между поверхностью цитоплазмы и оболочками возникнет тонкая прослойка выделенного материала, яйцо освобождается внутри оболочек и начинает поворачиваться, в соответствии с положением своего центра тяжести, богатым желтком вегетативным полушарием вниз и анимальной областью, вверх.
Стадия 1 – оплодотворенного яйца в первые минуты после оплодотворения, когда оно по своему виду еще не отличается от неоплодотворенного яйца (рис. 3).
Пигментный рисунок анимальной области не изменен, в центре ее светлое полярное пятно; оболочки плотно прилегают к яйцу и еще не начали набухать. Через несколько минут после осеменения (от 2 до 13 мин.) наружная оболочка яиц становится клейкой и прочной.

Рис. 3
Стадия 2 – оплодотворенного яйца после поворота и образования перевителлинового пространства (рис. 4).
Ко времени завершения поворота в верхней части икринки, между яйцом и оболочкой, становится различимой (если смотреть на яйцо сбоку) узкая щель, которая затем постепенно расширяется. В это время из цитоплазмы под оболочку выделяется осмотически активный коллоид (ранее содержавшийся в сети лакун анимальной области), который набухает, привлекая воду из окружающей среды; поверхность анимальной области яйца уплощается и между нею и оболочками образуется довольно значительное пространство, называемое перевителлиновым.
Своеобразный рисунок анимальной области яйца — светлое полярное пятно в центре и темные пигментные кольца вокруг него — после оплодотворения изменяется. Пигмент постепенно стягивается к центру анимальной области, образуя здесь темное скопление светлое полярное пятно, исчезает или, если оно было очень большим (что часто наблюдается у белуги), сильно уменьшается в размере.
Рис. 4
Стадия 3 – светлого серпа (рис. 5). У края анимальной области нередко заметна светлая, тогда совершенно белая полоса полулунной формы (светлый серп).
Полоска эта постепенно расширяется и превращается в светлый серп. Иногда серп бывает ярким, с четкими границами, в других случаях границы его менее определенны, в некоторых партиях его совсем не удается различить.
Специально проведенные наблюдения и опыты показали, что сторона яйца со светлым серпом становится затем спинной стороной зародыша, причем воображаемая плоскость, проведенная через середину серпа и центры анимальной и вегетативной областей, соответствует будущей среднеспинной плоскости зародыша. Таким образом, зародыш приобретает двусторонне-симметричное строение в самом начале развития, еще до того как яйцо начинает дробиться.

Рис. 5
2 этап (4–12 стадии)
Дробление. Оплодотворенное яйцо многократно делится и превращается в многоклеточный пузырек — бластулу. Клетки, возникающие при дроблении яйца, называются бластомерами.
Деление яйца можно видеть даже невооруженным глазом. Для этого надо смотреть на яйцо со стороны анимальной области. Удобно наблюдать дробление на приклеенной икре.
Стадия 4 – первого деления (рис. 6). Первая борозда делит сначала анимальную область на два бластомера. Затем борозда продвигается очень медленно на вегетативную часть яйца, т. к. огромное скопление желточных зерен и жира затрудняет ее распространение.

Рис. 6
Стадия 5 – второго деления (рис. 7). Анимальная область яйца разделяется на четыре бластомера приблизительно равной величины.
Еще не успеет первая борозда достигнуть экватора, как в центре анимальной области, перпендикулярно к ней, также в виде коротких белых полосок, закладываются борозды второго деления, и постепенно анимальная область зародыша разделяется на четыре части сходной величины. При постоянной температуре борозды второго деления появляются через промежуток времени, равный одной трети периода от осеменения до появления первой борозды.

Рис. 7
Стадия 6 – Третьего деления (рис. 8 - вид сверху).
При третьем делении начинается разделение яйца на 8 бластомеров. Расположение борозд при этом бывает разлит в яйцах удлиненной формы борозды закладываются параллельно первой борозде или под небольшим углом к ней, образуя характерную фигуру в виде буквы (Ж) в яйцах округлой формы они обычно располагаются по радиусам. Рисунок, образуемый бороздами дробления, как правило, не бывает геометрически правильным, бластомеры имеют разные форму и величину.

Рис. 8
Стадия 7 – четвертого деления (рис. 9). Борозды второго деления близки к смыканию; борозды третьего деления приближаются к экватору. При четвертом делении, когда в более узких бластомерах борозды располагаются горизонтально, отделяя к центру анимальной области бластомеры небольшой величины, а в более широких — косо или радиально.

Рис. 9
Стадия 8 – пятого дробления (рис. 10 - вид сверху)

Рис. 10
Стадия 9 – седьмого деления (рис.11).
Борозды полностью разделяют богатую желтком вегетативную область. При этом четко выступает неравномерность дробления, свойственная осетровым: бластомеры, обособляющие в верхней, анимальной части зародыша, на всех стадиях дробления имеют значительно меньшие размеры, чем вегетативные бластомеры.

Рис. 11
Стадия 10 – позднего дробления (рис. 12) Последовательное деление привели к уменьшению размеров бластомеров.

Рис.12
Стадия 11 – ранней бластулы (рис. 13). В анимальной области отдельные бластомеры еще хорошо различимы, при небольшом увеличении. Внутри зародыша между бластомерами образовалась полость дробления бластоцель, зародыш приобретает строение полого шарика – бластулы.

Рис. 13
Стадия 12 – поздней бластулы (рис. 14). В анимальной области отдельные клетки при небольшом увеличении не различимы. Между мелкими анимальными и относительно крупными вегетативными бластомерами имеется зона бластомеров промежуточного размера, носящая название краевой зоны.

Рис. 14
3 этап (13–18 стадии).
Гаструляция. После того как оплодотворенное яйцо, благодаря непрерывному ряду делений, превратилось в многоклеточный пузырек — бластулу, в нем начинаются сложные перемещения: материал краевой зоны вворачивается внутрь, а светлая анимальная область разрастается и покрывает снаружи темные вегетативные клетки. Эти процессы называют гаструляцией, а зародыш в описываемый период — гаструлой.
К концу гаструляции снаружи остается только светлый материал анимальной области бластулы, который теперь покрывает всего зародыша и образует наружный слой стенки гаструлы, а краевая зона и богатые желтком темные клетки вегетативной области все оказываются внутри, где они образуют внутренний, слой зародыша.
Стадия 13 – начало гаструляции (рис. 15). Немного выше экватора на спинной стороне зародыша образовалась узкая, сильно пигментированная полоска.

Рис. 15
Стадия 14 – ранней гаструляции (рис. 16). На месте пигментной полоски образовалось углубление бластопор.

Рис. 16
Стадия 15 –средней гаструлы (рис. 17). На этой стадии светлый анимальный материал покрывает 2/3 поверхности зародыша. Бластопор замыкается в кольцо.

Рис. 17
Стадия 16 –большой желточной пробки (рис. 18). Щель бластопора замыкается в кольцо, окружая темные клетки нижней части зародыша, эта область приобретает вид темной пробки, воткнутой в светлый круглый пузырек, от чего она и получила название желточной пробки
Рис 18
Стадия 17 – малой желточной пробки (рис. 19). Вся поверхность зародыша, за исключением желточной пробки, небольшого размера, покрыта светлым анимальным материалом
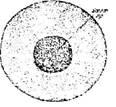
Рис. 19
Стадия 18 – щелевидного бластопора (рис. 20). Темный вегетативный материал полностью покрыт светлым анимальным (процесс гаструляции закончен). Края бластопора сомкнулись, между ними образовалась узкая щель

Рис. 20
4 этап (19–28 стадий).
Дифференциация зародышевых пластов на зачатки основных органов.
По окончании гаструляции у зародыша постепенно формируются зачатки основных систем органов: нервной, пищеварительной, выделительной, кровеносной, а также мускулатуры и хорды. Зародыш на этом этапе развития по-прежнему неподвижен, питается за счет желтка и дышит всей поверхностью тела. Многие из тех изменений, которые происходят с зародышем, можно проследить, наблюдая за развитием при помощи простой лупы. Покровы зародыша очень тонки и полупрозрачны, поэтому внутренние органы просвечивают через них. Обладая острым зрением, можно многое увидеть, рассматривая зародыша даже простым глазом.
Стадия 19 – ранней нейрулы (рис. 21). На спинной стороне зародыша образуется нервная пластинка. Начинают обозначатся нервные валики вокруг головного отдела нервной пластинки, они еще мало приподняты.

Рис. 21
Стадия 20 – широкой нервной пластинки (рис. 22) Нервные валики вокруг головного отдела нервной пластики четко обозначены.
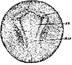
Рис. 22
Стадия 21 – начало сближения нервных валиков. Впервые обозначаются зачатки выделительной системы в виде коротких светлых тяжей, просвечивающих через покровы. Они располагаются по бокам от туловищного отдела нервной пластинки.
Стадия 22 - поздней нейрулы (рис 23). Нервные валики в туловищном отделе сближаются. Зачатки выделительной системы удлинились и видны более четко.

Рис. 23
Стадия 23 - замкнувшейся нервной трубки (рис. 24). Нервная пластинка свернулась в трубку, нервные валики сомкнулись. Шов в области их слияния имеет вид неглубокой бороздки.
Рис. 24
Стадия 24 - появление глазных выростов и утолщения переднего конца зачатков выделительной системы (рис. 25). Шов в области слияния нервных валиков менее заметен. В задней части переднего мозгового пузыря образовались глазные выросты. По бокам от заднего мозгового пузыря возникли зачатки внутреннего уха. Впереди головного мозга, примыкая к нему, обозначается светлая пластинка — зачаток железы вылупления. В передней части закладок выделительной системы образуются утолщения.
Рис. 25
Стадия 25 - сближения боковых пластинок и образования утолщения в области зачатка хвоста (рис. 26) По бокам от переднего конца головного мозга образовались зачатки обонятельных мешков. Впереди мозга менее виден зачаток железы вылупления. Боковые пластинки (из которых позднее образуется сердце, а также выстилка околосердечной полости и полости тела) достигли переднего конца головы; передние суженные выросты их сближаются. В зачатке почки дифференцируются почечные канальцы. В заднем конке зародыша возникло возвышение - еще необособленный зачаток хвоста.
Рис. 26
Стадия 26 — слияния боковых пластинок и начала обособления хвостового отдела зародыша (рис. 27). Сблизившиеся ранее передние выросты боковых пластинок сливаются, и в месте их соединения начинается образование зачатка сердца. Петля, образуемая выводным почечным каналом, значительно удлинилась. Начинается обособление зачатка хвоста, имеющего в это время форму короткой и широкой лопасти

Рис. 27
Стадия 27 - короткой сердечной трубки (рис. 28). Начинается процесс обособления головы - распластанные на брюшном отделе тела зачатки головных органов начинают стягиваться к средне - спинной линии. Образовался зачаток сердца, имеющий вид короткой трубочки. Зачаток хвоста удлинился и слился.

Рис. 28
Стадия 28 - прямой удлиненной сердечной трубки (рис. 29). Зародыш не подвижен, туловищные мышцы еще не реагируют на раздражение сокращением. Голова уже заметно приподнята. В связи с этим зачаток железы вылупления приобрел палочковидную форму.
Рис. 29
5 этап (29–36 стадии).
Развитие зародышей от начала пульсации сердца до вылупления. Уловить первые сокращения сердца нелегко, так как они очень слабы и от одного сокращения до другого проходит много времени.
Но довольно скоро сокращения становятся сильнее, и сердце начинает пульсировать с определенным ритмом. Чтобы наблюдать сердцебиения, нужно положить зародыша на часовое стекло пли в чашечку и в течение нескольких минут под лупой внимательно следить за сердцем.
После того как сердце начнет пульсировать, мышцы туловища тоже приобретают способность сокращаться; если зародыша уколоть в спину иглой, замечаются слабые мышечные подергивания.
В период от начала кровообращения и до вылупления значительно изменяется внешняя форма зародыша. Голова обособляется и немного увеличивается, хвост сильно вырастает в длину, распрямляется и превращается в мощный орган движения личинки. Только брюшной отдел (желточный мешок) изменяется мало; благодаря большому количеству желтка в клетках стенки кишки он сохраняет крупные размеры.
Когда голова, раньше распластанная на желточном мешке, приподнимается, зачаток железы вылупления смещается на ее нижнюю поверхность, а сердце, на более ранних стадиях расположенное впереди головы, теперь оказывается под нею.
Продолжается развитие головного мозга и органов чувств. В том месте, где глазной пузырек соприкасается с покровным эпителием, образуется зачаток хрусталика. Обонятельные ямки углубляются. Слуховые пузырьки увеличиваются, стенка их становится тоньше; в это время они слегка просвечивают через эпителий по бокам от продолговатого мозга.
Очень заметны изменения в хвостовом отделе. Он значительно удлиняется, распрямляется и становится плоским, на спинной и брюшной сторонах его образуется широкая плавниковая оторочка. По мере удлинения хвост, будучи прижат оболочками к желточному мешку, загибается на брюшную сторону; при этом конец его постепенно достигает сердца.
Стадия 29 - образования изгиба сердечной трубки (рис. 30). Зародыш неподвижен, из – за раздражения начинает отвечать слабыми мышечными подергиваниями. Сердечная трубка значительно удлинилась и изгибается. Сердце начинает пульсировать.
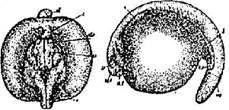
Рис. 30
Стадии 30 - конец хвоста приближается к сердцу (рис. 31). Хвостовой отдел склонен набок, хвост начал распрямляться, уплетаться, вокруг него появляется узкая, еще плохо различимая плавниковая оторочка.

Рис. 31
Стадия 31 - конец хвоста достигает сердца (рис. 32). Хвостовой и спинной отделы наклонены набок. Если снять оболочки, зародыш делает маятникообразные плавательные движения - «плавание на месте». Хвост значительно распрямляется. Плавниковая оторочка хорошо различима. На описываемом этапе развития определять стадию удобнее всего по положению конца хвоста; нужно только иметь в виду, что между разными видами имеются некоторые отличия (так, у стерляди хвост относительно длиннее, чем у белуги, осетра и, в особенности, севрюги); поэтому следует всегда отмечать, о каком виде идет речь.
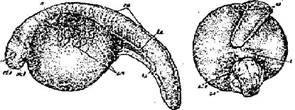
Рис. 32
Стадия 32 - конец хвоста касается головы, плавниковая оторочка шире (рис. 33).

Рис. 33
Стадия 33 - конец хвоста немного заходит за голову, достигая уровня глаз (рис. 34). Голова наклонена на бок, плавниковая оторочка хвоста расширилась. Если снять оболочки хвост полностью распрямляется.

Рис. 34
Стадия 34 – конец хвоста достигает начала продолговатого мозга (рис. 35). Зародыш активно двигается в оболочках, если оболочки снять, способен к слабому поступательному движению.
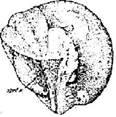
Рис. 35
Стадия 35 – начала выклева конец хвоста достигает почки (рис. 36). Выклев у белуги начинается, когда конец хвоста достигает петли, образованной собирающим и выносящим почечными каналами. В оболочках зародыш активно двигается; если их снять, плавает.
Желточный мешок имеет яйцевидную форму. В глазу может появиться пигментное пятно. Намечается ротовое углубление. Иногда обозначаются зачатки грудных плавников. Кровь бесцветная или желтовато - розовая.
 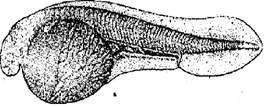
Рис. 36
1.3 Постэмбриональное развитие
Период постэмбрионального развития осетровых рыб (36 – 45 стадии).
Стадия 36 - массовое вылупление (рис. 37). Форма желточного мешка яйцевидная. В глазу четкое пигментное пятно. В жаберной области обозначились складочки 2 первых жаберных карманов. На нижней поверхности головы заметно ротовое углубление. Жаберных щелей и ротового отверстия еще нет. Позади почек едва заметны зачатки грудных плавников. Кровь желтовато-розовая.
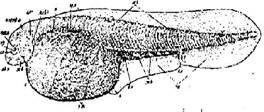
Рис. 37
Стадия 37 - появляются зачатки усиков (рис. 38); прорывается ротовое отверстие. Начинается разделение желточного мешка на желудочный и кишечный отделы. Четко выражены зачатки грудных плавников в виде небольших складочек кожи. Зачатки жаберных лепестков отсутствуют. Появляется зачаток боковой линии сейсмосенсорной системы.
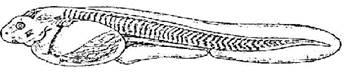
Рис. 38
Стадия 38 - различимы первые меланоциты (рис. 39). Энтодермальная складка, отделяющая желудок от кишечника, неполная. Формируются первые мускульные почки в области спинного и анального плавников. Появляется зачатки жаберных лепестков на жаберной крышке и первой жаберной дуге. Боковая линия сейсмосенсорной системы достигает уровня кювьерова протока.
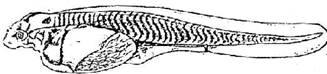
Рис. 39
Стадия 39 - желудок отделен от кишечника; обособились спинной и анальные плавники (рис. 40). Боковая линия сейсмосенсорной системы достигает уровня заднего грудного плавника; появляется добавочный ряд сейсмосенсорной системы.

Рис. 40
Стадия 40 - различим зачаток брюшного плавника в виде узенькой складочки кожи (рис. 41). Брюшные отростки мышечных сегментов в области грудного плавника спускаются ниже его основания. Можно наблюдать первые не регулярные движения нижней челюсти. Боковая линия сейсмосенсорной системы не достигает уровня конца желудка, добавочный ряд заканчивается над грудными плавниками.

Рис. 41
Стадия 41 - края обонятельных лопастей смыкаются, но еще несращены (рис. 42). Боковая линия сейсмосенсорной системы заканчивается над спиральной кишкой, ее добавочный ряд заходит за заднюю границу грудного плавника, образуется короткий зачаток спинного ряда.
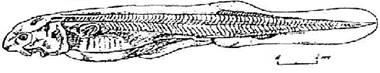
Рис. 42
Стадия 42 – появляется зачаток пилорического придатка (рис. 43).
Лопасти обонятельного органа сращены. Боковая линия сейсмосенсорной системы достигает уровня переднего края брюшного плавника: спинной ряд начинает изгибаться.
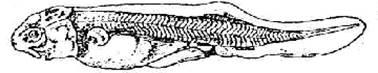
Рис. 43
Стадия 43 - рострум принимает горизонтальное положение (рис. 44).
Брюшной плавник достигает края прианальной складки. Появляются зачатки вторичных лепестков в первой жабре. Боковая линия сейсмосенсорной системы достигает уровня заднего края брюшного плавника, добавочный ряд заканчивается над спиральной кишкой, спинной ряд изогнут, и начинает расти параллельно боковой линии.
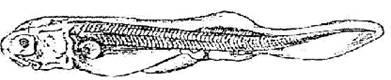
Рис. 44
Стадия 44 - основания усиков выносятся вперед, и концы их не достигают передней границы рта (рис. 45). Массовый выброс пигментных пробок. Лопасти брюшных плавников опускаются ниже края прианальной складки. В спинной плавниковой складке появляется мезенхимная полоска (общий зачаток спинных жучек). Передняя поперечная комиссура сейсмосенсорной системы переместилась дорзально и не видна с брюшной стороны; боковая линия заходит за уровень заднего края брюшного плавника, дополнительный ряд не доходит до первой границы брюшного плавника.
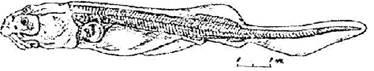
Рис. 45
Стадия 45 — перехода на активное питание (рис. 46). Видны разделительные зачатки жучек в спинной плавниковой складке. Боковая линия сейсмосенсорной системы заходит за уровень .средней части спинного плавника, спинной ряд заходит за уровень заднего края грудною плавника.
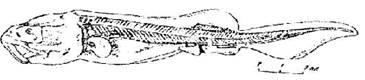
Рис. 46
Личиночный период развития осетровых рыб.
В личиночном периоде обычно выделяют два этапа:
I личиночный этап - этап смешанного питания.
Дыхание становится наружно-жаберным. Появляются зачатки брюшных плавников. Рот нижний, перед ним четыре усика. Голова большая Рыло постепенно становится заостренным. Плавниковая кайма на месте непарных плавников обособляется. Хвостовой плавник большой, гетероцеркальный. Вдоль спинного края плавниковой складки начинаются появляться бугорки - зачатки жучек. Сначала они еще не выходят за пределы складки. На верхней челюсти закладываются зубы. Питание у зародышей смешанное, этап длится 3 дня, анальное отверстие замкнутое, у личинок сформированное. Меланин, имевшийся в кишечнике зародыша выделен.
II личиночный этап - экзогенного питания.
Желточный мешок полностью резорбировался, питание экзогенное. Рот личинки при открывании выдвигается, но ротовой трубки, типичной для более позднего малькового этапа еще нет.
Увеличивается число и длина жаберных лепестков. Жаберная крышка разрастается, покрывая собой наружные жаберные лепестки почти до трети их длины. Образовался спиральный клапан. Длина личинок 21 мм.
Мальковый период развития осетровых рыб.
Личиночные плавниковые складки резорбируются. Вентральная поверхность туловища малька становится плоской, имеются ряды спинных, боковых и брюшных жучек. На голове развиваются покровные типы. Жучки и костные пластинки образуют на теле малька защитный и колючий панцирь. Жаберные крышки разрастаются полностью прикрывая жаберные лепестки. В отличие от личинок челюсти малька не имеют зубов. Зубы исчезают тогда, когда жаберные крышки начинают функционировав как всасывающий аппарат. При этом утрачивается необходимость удерживать добычу зубами. Питание бентосом (Детлаф, Гинзбург, 1954).
Глава 2. Влияние различных факторов среды на белугу
Рыбы, как и все другие организмы, обитающие в водоемах, находятся в тесном взаимодействии с абиотическими факторами среды — совокупностью условий неживой природы, и биотическими факторами среды, включающими в себя разнообразные отношения между организмами. Внешняя среда влияет на все жизненные процессы, происходящие в организме рыбы: дыхание, питание, кроветворение и кровообращение, нервную деятельность, размножение, рост и развитие. Рыба на разных стадиях своего развития и в различные периоды жизни неодинаково реагирует на условия внешней среды. Богатство и многообразие ихтиофауны водоема зависит от многих факторов среды, действующих на организм рыбы, в частности от площади и глубины водоема, характера его грунта, от населяющих его бактерий, растений и животных, от гидрологического и гидрохимического режима и так далее. Чтобы правильно оценить ту огромную роль, которую играют факторы внешней среды в жизни рыб, достаточно рассмотреть лишь наиболее важные из них, с которыми чаще всего приходится сталкиваться специалисту, работающему в области рыбоводства: температуру воды, освещенность, уровень и течение воды, гидрохимический режим, кормовую базу (Иванов, 1988).
2.1 Влияние температуры
Современная биотехнология искусственного разведения осетровых рыб всецело зависит от температурного режима источников водоснабжения. По этой причине сроки работы с производителями, инкубация икры, подращивание личинок осетровых на действующих рыбоводных предприятиях определяются узкими временными границами, не редко выходящими за пределы оптимальных для этих процессов. Для решения данной проблемы необходим перевод начальных звеньев биотехнического процесса искусственного воспроизводства осетровых рыб на управляемый режим (Кокоза, 2004).
Температура воды является одним из факторов, оказывающих большое воздействие на отклонение жизненных функций рыбы, определяющих ее рост и развитие. Этот фактор действует на рыбу как непосредственно — изменяя интенсивность ферментативных процессов, происходящих в организме, активность потребления пищи, характер обмена веществ, ход развития половых желез.Температура является также внешним стимулом, определяющим для физиологически подготовленного организма рыб начало миграций, нереста и зимовки. Температурные условия, при которых все жизненные процессы протекают в организме нормально, принято называть оптимальными. Исходя из оптимальных температурных условий, все виды рыб условно подразделяют на теплолюбивых и холодолюбивых (Иванов, 1988).
Белуга это относительно теплолюбивый вид в связи с этим она может обитать в водоемах, в которых температура воды изменяется в течение года от 0 до 30° С. и даже несколько выше. Весной взрослые особи предпочитают 2-160
С, молодь - 7-200
С. Летом при температуре воды 20-310
С белуга равномерно распределяется по акватории моря, осенью предпочитает 8-150
С. На пастбищах средней части Каспийского моря белуга летом предпочитает температуру 17-190
С, в южной части - 10-290
С, при этом наибольшее количество особей отмечается при 22-290
С. Зимой белуга нагуливается при температуре 4-50
С в средней части и при 10-120
С в южной части моря (Легеза, 1972).
2.2 Влияние освещенности, уровня и течения воды на белугу
Прозрачность воды зависит от содержания в толще воды неорганических и органических взвешенных частиц, а также мельчайших растительных и животных организмов. Чем больше в воде содержится взвешенных частиц и микроорганизмов, тем меньше ее прозрачность и, наоборот. Поздней осенью и зимой вода бывает более прозрачна, чем весной и летом, так как весенний паводок и летние ливневые дожди приносят в водоем большое количество неорганических и органических веществ, в это же время года в воде бурно развиваются одноклеточные водоросли и беспозвоночные животные. От освещенности у большинства рыб зависит строение органа зрения, которое играет значительную роль при их ориентировке во время движения и суточном ритме активности. Этот фактор среды обусловливает возможности охоты хищника за жертвой и защиты жертвы от хищника. Освещенность влияет на развитие рыб. Так, у многих видов рыб в эмбриональный период нарушается обмен веществ, если развитие происходит в несвойственных для них условиях освещенности (Иванов, 1988).
Что касается условий освещения при развитии икры белуги на нерестилищах, то по этому поводу можно только сказать, что, так как икра развивается на значительной глубине (например, на нерестилищах Кубани на глубине от 0,5 до 3,5 м, на Волге от 4 до 9 м) и при слабой прозрачности воды (соответственно, на Кубани от 5 до 15 см, а на Волге от 35 до 50 см интенсивность ее освещения не может быть сколько-нибудь значительной ( Драгомиров, 1961).
Освещенность оказывает также большое влияние на нерестовые миграции проходных и полупроходных рыб из моря в реку, их нерест и миграцию их молоди из реки в море. Для большинства видов наших проходных рыб (осетровые, лососевые) течение воды в реке является стимулирующим фактором для созревания половых продуктов и при нересте (Иванов, 1988).
2.3 Влияние гидрохимических показателей
Развитие осморегуляторной системы обусловлено морфо-функциональным формированием органов, участвующих в водно-солевом обмене (хлоридные клетки жабр, почки, гипоталамо-гипофизарный комплекс, интерреналовая и щитовидная железы и др.). Становление этих органов происходит на ранних этапах онтогенеза осетровых. Именно такое развитие обусловливает расширение степени эвригалинности с возрастом, выявленной у молоди осетровых различных видов. Это явление следует рассматривать как частное проявление общего процесса эврибионтности в ходе онтогенеза. Расширение степени эвригалинности с возрастом проявляется в увеличении диапазона солености, выдерживаемой молодью (Драгомиров, 1961).
Белуга в период до перехода на активное питание (I – III этапы) выдерживает соленость до 5,4‰. Вскоре после начала экзогенного питания(IV этап) степень ее эвригалинности увеличивается, молодь выживает в солености 8,7 – 10,5‰. Стандартная молодь (средний вес 3,5г, средняя длина 100мм) адаптируется к соленности10,5‰ (выживаемость 100%). Высокая выживаемость в морской воде соленостью 12,5‰ наблюдается у молоди белуги более крупных размеров (средняя длина 122 мм, средний вес 5,5г). Молодь осетровых с развитой осморегуляторной системой способна успешно (при 100% выживаемости) адаптироваться к морской воде соленостью 12,5‰. В этом случае при переходе из пресной воды в морскую она может поддерживать относительное постоянство к осмолярности сыворотки крови: в первые 12 часов солевого воздействия осмолярность сыворотки крови поднимается до уровня осмолярности морской среды, превышая исходный уровень на 25 – 28%, но уже в последующие 12 часов осмолярность сыворотки крови начинает снижается, достигая через 24 часа адаптации уровня, превышающий исходный на 14 – 17%, через 72 часа – на 5 – 8%. На ранних этапах онтогенеза, когда происходит интенсивное становление органов, развитие осморегуляторной системы является производным степени сформированности организма в целом, которая в большей мере связанна с размерами и весом рыб, чем с их возрастом. Этим можно объяснить тот факт, что у одновозрастной, но разноразмерной молоди адаптации к одной и той же солености происходит по-разному. Чем крупнее молодь, тем быстрее проходит адаптация. Молодь осетровых одного возраста, но различных размеров и веса, выращенная в одних и тех же условиях, неравноценна по состоянию осморегулярной системы, в результате чего проявляется ее различная реакция на солевое воздействие (Вернидуб, 1952).
Молодь белуги в возрасте 40 суток (средняя длина 13,7см, средний вес 9,5г) способны к осморегуляции, обеспечивающей ее адаптацию к солености 12,5‰. При переводе такой молоди из пресной воды в соленую осмолярность сыворотки крови после ее подъема в первые 12 часов солевого воздействия начинает снижаться и через следующие 12 часов воздействия превышает исходный уровень на 16,3%. На 3 сутки адаптации осмолярность сыворотки крови значительно снижается и достигает уровня, которой превышает исходный на 8,2%. Подобную реакцию на солевое воздействие обнаруживает осморегуляторная система молоди белуги несколько меньших размеров.
Молодь белуги того же возраста и выращенная в тех же условиях, что и первая группа молоди, но меньших размеров, которые считаются стандартными (средняя длина 10см, средний вес 3,5г) неспособна поддерживать относительное постоянство осмолярности сыворотки крови. При переводе такой молоди в воду с соленостью 12,5‰ осмолярность сыворотки крови продолжает подниматься после 12 часов солевого воздействия выше осмолярности воды и молодь погибает (Детлаф. Гинзбург, 1954).
Можно заключить, что у молоди белуги осморегуляторная система оказывается сформированной в значительной степени по достижению ее размеров 12 – 13 см, привесе около 6 г. Сформированная осморегуляторная система молоди осетровых проходных форм характеризуется способностью быстро переходить от гипертонического типа осморегуляции к гипотоническому при смене пресной воды на морскую, в результате чего осмолярность сыворотки крови после ее подъема в первые 12 часов солевого воздействия в течении следующих 12 часов начинает быстро снижаться и на третьи сутки адаптации достигает уровня, близкого к исходному ( Вернидуб, 1952).
Для искусственного разведения создания оптимальных условий развития важнее знать не пороговые значения концентрации кислорода, а критические концентрации, ниже которых начинается угнетение дыхания. Критическая концентрация для мальков осетра и севрюги, в отличие от пороговой, при увеличении температуры сильно возрастает. При 11° C она почти равна пороговой, при 18° C равна уже 4,0—4,5 мг/л, при 25° — 5,0 — 5,5 мг/л, а при 27° — 8,25 мг/л и больше.
По данным О. Л. Гордиенко (1953), успешное выращивание молоди белуги возможно при содержании кислорода в воде от 10,0 до 6,5 мг/л. При снижении содержания кислорода до 3,6—3,88 мг/л состояние молоди становится угнетенным, а при 2,32— 1,41 мг/л наступает гибель.
Одним из важных моментов в характеристике условий развития икры является реакция среды. В экспериментальных условиях оптимальная для развития икры реакция среды измеряется рН от 6,4—6,5 до 7,5, а для мальков рН от 7 до 8. Граница выживаемости несколько шире: для икры рН от 6,1 до 8,0, а для мальков от 6,4 до 8,0. В зонах крайних для икры значений рН—от 6,1 до 6,4 и от 7,5 до 8 — уже заметно возрастает количество аномалий развития (Драгомиров, 1961).
Глава 3. Управление половыми циклами у рыб различными методами
В практике искусственного рыборазведения применяют три метода стимулирования созревания половых продуктов у производителей рыб: экологический, физиологический и эколого-физиологический (Мухачев, 2005).
Экологический метод разработан в 30–е годы академиком А. Н. Державиным. Применяется, главным образом при разведении осетровых, лососевых, окуневых и реофильных карповых рыб (Мухачев, 2005). Он считал, что при выдерживании производителей следует создавать условия внешней среды, соответствующие естественным, в которых происходит развитие половых продуктов. Поскольку в природе икра и сперма созревают во время хода рыбы на нерест против течения воды, то этот фактор А. Н. Державин считал основным, влияющим на ускорение созревания половых продуктов. Он рекомендовал для выдерживания и получения зрелых производителей использовать овальные садки длиной 25 м, шириной 6 м и глубиной до 1,2 м, в которых создавалось течение и имитировались речные условия (быстротоки и т. д.). На дно таких садков насыпается галька. Водоснабжение в садке механическое, расход воды 20 л/с. Улучшение циркуляции воды достигается устройством в средней части садка по его длине бетонной" стены протяженностью 19 м. В каждый садок помещают по 50 рыб, самок и самцов раздельно. Наряду с течением в садках создают благоприятный температурный и кислородный режим. Однако опыт работы с такими садками показал, что в них созревает только одна треть производителей, к тому же трудно определить момент, когда надо брать икру (Мильштейн, 1982).
Физиологический метод. Этот метод основан на инъекционном введении производителям препарата гипофиза. Хорошо зарекомендовал себя этот метод при разведении осетровых и карповых рыб.
Гипофиз — железа, расположенная у основания головного мозга. Гормоны гипофиза влияют на рост и развитие организма, на созревание половых продуктов. Заготавливают гипофизы в осенне-зимний период или ранней весной. Извлекаемый у рыб гипофиз помещают в сосуд с химически чистым ацетоном, который несколько раз меняют в течение 12 ч до полного обезжиривания гипофиза. Количество ацетона должно в 10—15 раз превышать объем гипофизов. Повторное использование ацетона недопустимо.
После обезжиривания гипофизы высушивают в затененном месте на фильтровальной бумаге и хранят в сухих плотно закрытых пробирках или целлофановых пакетах с этикетками.
В связи с постоянным ростом масштабов рыборазведения с каждым годом требуется все больше и больше гипофизов, поэтому учеными предложен ряд его заменителей. К числу таких заменителей относится хориогонин (хориальный гонадотропин). Хориогонин, вырабатываемый на заводах медицинской промышленности, в отличие от гормонов гипофиза можно точно дозировать.
Для инъецирования рыб высушенные гипофизы измельчают в фарфоровой ступке, разбавляют физиологическим раствором (6,5 г химически чистой поваренной соли, растворенной в 1 л дистиллированной воды) и растирают до получения однородной кашицы (суспензии). Полученную суспензию гипофиза вводят рыбе шприцем в мышцы спины. Необходимое количество гипофиза определяют в зависимости от температуры воды, пола, массы рыбы и активности препарата. Инъекции дают хорошие результаты только при нерестовых для данного вида рыб температурах воды и при завершении у производителей IV стадии зрелости половых продуктов.
Активность гипофиза определяют по методу Б. Ф. Гончарова. Он заключается в выдерживании пробы икры, отобранной у самки с помощью щупа, в физиологическом растворе с 0,1 %-ным раствором кристаллического альбумина. При добавлении определенного количества гипофиза у икринок растворяется зародышевый пузырек. Наименьшее количество гипофиза, вызвавшее растворение пузырька, является показателем активности гипофиза. Гипофизы рыб разных семейств обладают специфичностью действия. Так, положительная реакция у осетровых наблюдается только при введении им гипофизов, взятых у особей этого же семейства, или при введении гипофиза леща. У большинства видов лучшие результаты дает двукратное введение гипофиза. При первом (предварительном) инъецировании вводят от 1/8 до 1/3 всей дозы препарата. Оставшуюся часть гипофиза используют при второй (разрешающей) инъекции. Самцов, как правило, инъецируют один раз и вводят им в 2 раза меньше гипофиза, чем самкам (Черномашенцев, Мильштейн, 1983).
Эколого – физиологический метод. Он предусматривает стимулирование созревание половых продуктов у производителей путем комбинированного воздействия на организм рыбы экологических факторов внешней среды и вводимых физиологически активных веществ. Это дает возможность рыбоводу получать в определенный день и даже час необходимое количество икры и спермы, что позволяет планировать работу рыбоводного предприятия по каждому звену биотехнического процесса (Мухачев, 2005).
Примером эколого-физиологического метода является содержание осетровых в садках Куринского типа, бассейнах конструкции Б.Н. Казанского.
В садковом хозяйстве конструкции Б. Н. Казанского имеются земляные пруды для длительного резервирования и расположенные вблизи них бетонные садки-бассейны, предназначенные для кратковременного содержания производителей.
Самок и самцов содержат раздельно. Земляной пруд состоит из двух частей: основной, расширенной, имеющей глубину до 2,5 м, и суженной, более мелкой части с глубиной 0,5—1 м. В этой части пруда создаются условия, имитирующие подход к нерестовому плесу. В расширенной части с большей глубиной условия приближаются к режиму зимовальных ям.
Пруд для самок имеет следующие размеры: длина 130 м (расширенная часть 100 м и суженная 30 м), ширина 20—25 м в расширенной части и 4—6 м в суженной. Дно расширенного участка земляное, а в суженном вымощено мелким гладким булыжником на обедненном бетоне; на месте стыка расширенной и суженной частей рассыпана галька.
Самцов содержат в прудах более простой конструкции. Эти пруды не имеют суженной части. В таком водоеме можно проводить в случае необходимости и зимовку производителей. Длина пруда для самцов 120 м, ширина по дну 5 м, глубина 2,5 м, уклон откоса 1:3.
Водоснабжение прудов механическое, водовпуск имеет вид железобетонного лотка или трубы. Сброс воды осуществляется через водоспускное сооружение, обеспечивающее как полное осушение пруда, так и возможность спуска различных горизонтов воды. Уровень воды регулируется шандорами. Постоянный расход воды 30 л/с может быть увеличен до 300 л/с (Мильштейн, 1982).
Садковое хозяйство куринского типа. Представляет собой земляной пруд размером 75x12 м, разделенный на три участка с помощью бетонного перегораживающего сооружения, в середине которого устроено отверстие для установки затвора. На первом участке длиной 105 м и глубиной 3 м производится длительное выдерживание производителей — от 1 до 1,5 мес. Заполнение водой продолжается 10—12 ч, а сброс 5—6 ч.
При наступлении нерестовых температур производителей переводят на второй участок, представляющий собой овальный бетонный бассейн с вертикальными стенками. В бассейне длиной 7 м, шириной 5 и глубиной 1 м производится предварительное кратковременное выдерживание самок и самцов до инъекции (1—3 сут). Переход из первого во второй участок осуществляется в форме плавного подъема: орудия лова — волокуши, которыми отлавливают производителей, подтягивают по специальным направляющим электролебедками с дистанционным управлением. Второй участок заполняется водой за 30 мин. На третьем участке производителей инъецируют и выдерживают после гипофизарной инъекции. Этот участок имеет 2 бетонных бассейна с вертикальными стенками. Длина бассейна 5 м, ширина 3,5, глубина 1 м. На заполнение и сброс воды отводится по 15 мин. Над бассейном устраивается навес. Пересадка производителей из второго на третий участок, а также доставка их в операционное отделение, где получают икру, производится самоходным электротельфером в люльках.
Ранней весной из отстойника подают более теплую воду, что позволяет инъецировать рыбу в более ранние сроки. В бассейнах производители находятся 1–3 сут. Подача и сброс воды из бассейнов независимые. Вода подается с помощью трубы (флейты), расположенной поперек бассейна. Струи воды из флейты направлены в противоположные стороны. В результате такой подачи воды улучшается кислородный режим.
В бассейн сажают по 50 производителей белуги, по 80 — осетров или шипа и по 100 — севрюги. Расход воды в бассейнах 30 л/с. Третий участок огораживается штакетным забором, вокруг которого сажают деревья (Мильштейн, 1982).
При наступлении нерестовых температур производителям делают гипофизарную инъекцию.
Приготовление препарата гипофиза. Перед инъецированием ацетонированные гипофизы растирают в чистой стеклянной или фарфоровой ступке пестиком в порошок. Порошок развешивают в соответствии с необходимой дозировкой в биологических единицах на аналитических или горизонтальных весах на каждую партию инъецированных производителей — раздельно самцов и самок. То количество препарата гипофиза, которое необходимо для данной порции производителей, помещают в ступку, смачивают физиологическим раствором (0,65 % раствор хлористого натрия) и тщательно перемешивают. Затем к образовавшейся кашице добавляют физиологический раствор в таком же количестве, чтобы на одного производителя приходилось 2 см3
(осетровые) или 1 см3
(карповые, окуневые) суспензии. Суспензию тщательно взбалтывают и переносят в низкую склянку с широким горлышком и притертой пробкой. Перед началом инъекции содержимое склянки еще несколько раз перемешивают путем втягивания жидкости шприцем. После этого суспензию набирают в шприц и вводят сбоку в спинную мускулатуру рыбы передней части тела. Иглу вынимают осторожно. Место прокола кожи прижимают пальцем, а затем немного массируют для предотвращения вытекания введенного препарата. В настоящее время кроме суспензии ацетонированных гипофизов применяют глицериновый гипофизарный препарат.
Проведение гипофизарной инъекции у осетровых рыб. При инъецировании производителей осетровых препаратом гипофиза необходимо учитывать температуру воды. Инъецирование производителей белуги проводят при температуре воды от 7 до 17°С, осетра и шипа - от 9 до 19 С, севрюги при температуре от 15 до 24°С. Доза гипофиза вводимого производителям рассчитывается на I кг массы самки (табл. 1).
Таблица 1
Доза препарата гипофиза для производителей осетровых на 1 кг массы (мг).
| Вид рыбы |
Масса, кг |
Температура воды, °С |
| 7 - 9 |
10 - 12 |
13 - 15 |
16 - 18 |
19 - 24 |
| Белуга |
100 - 140 |
2,5 |
2 |
1,5 |
| Шип |
24 - 35 |
2,5 |
2 |
1,5 |
| Осетр |
20 - 25 |
2 |
2,5 |
2 |
1,5 |
| Севрюга |
10 - 12 |
2,5 |
2 |
1,5 |
После инъекции самцы становятся зрелыми раньше самок, поэтому самцам препарат вводят позднее. Разница во времени созревания половых продуктов при различных температурах следующая: при 14-16°С-4-5 часов, при 17-19°С -3-4 часа. В качестве заменителей препарата гипофиза, для инъецировании производителей осетровых рыб, в настоящее время применяют его искусственные и естественные аналоги - сурфагон и люлеберин (Пономарева, Бахарева, 2001). На современном этапе развития аквакультуры многие ученые рекомендуют использовать эколого–физиологический метод для стимулирования половых продуктов у осетровых в частности белуги.
Глава 4. Биологические основы кормления белуги
Белуга — хищник и предпочитает питаться рыбой. В пищевом комке белуги длиной 21-30 см на северокаспийских пастбищах 85 % приходилось на бычков, у рыб длиной 31-50 см главным объектом питания являлась обыкновенная килька. На втором году жизни белуга почти полностью переходит на хищный образ жизни: доля рыб в пищевом комке достигает 98 % (Рыбы Казахстана,1986).
Спектр питания белуги в естественных условиях очень разнообразен так летом в Северном Каспии основу пищи на западе района составляла вобла, на востоке - судак, бычки и килька обыкновенная. Из ракообразных потреблялись рак, краб и креветки (Ходоревская, Полянинова, 2000).
В Среднем Каспии спектр питания белуги состоял из равных частей рыбного корма (сельдевые - 45,6 %) и ракообразных (креветки, а также Gammaracantusloricatuscaspius, Dikerogammarushaemobaphisи Amathillinacristata). Интенсивность питания была низкой, накормленность составила 8 %оо, что в 2,5 раза ниже среднемноголетней величины. Молодь белуги имела большую накормленность, чем взрослые особи, - 29,8 против 2,8 %оо соответственно. Индекс наполнения у белуги, пойманной на глубине 25-50 м, составил 20,3 против 1,9 %оо у рыб, пойманных на глубине до 25 м. Интенсивность питания самок была значительно выше, чем у самцов, - 14,6 и 3,2 % соответственно (Захарова, Хураськин , Полянинова, 2000).
В Южном Каспии пищевой рацион белуги состоял из рака p. Astacus - 80,3 % и рыбного корма (килька обыкновенная, пузанок, бычки) - 19,2 %. Накормленность хищника в Южном Каспии составляла 49,5 %оо.
Несмотря на то, что запасы кильки обыкновенной в Каспийском море за последние годы увеличились, в пищевом рационе белуги этой рыбы не стало. Частный индекс наполнения желудков килькой обыкновенной в 80-е годы в среднем равнялся 9,4 %оо, в 90-е годы он снизился до 1,1 %. Уменьшение количества кильки в пищевом рационе белуги связано, по-видимому, с разобщенностью площадей нагула хищника и жертвы.
Интенсивность питания белуги в Южном Каспии оказалась самой высокой. Средний индекс составлял в этом районе 49,5 ‰0, в то время как в Среднем Каспии он равнялся 8,0‰0, а в Северном – 6,5‰0. Отсюда следует что для нагула белуги северо-каспийские пастбища теряют первостепенное значение, на первое место выходят пастбища Южного Каспия. У молоди на первом году жизни наблюдается смешанное питание: ракообразные, моллюски и рыба. Сеголетки белуги (табл. 2) в первый месяц пребывания в море (июнь) питаются в основном нектобентическими высшими ракообразными из отряда Mysidacea, только в 1976 и 1978 гг. в рационе питания в большом количестве появились бычки, что было обусловлено снижением численности мизид в маловодный период (Полянинова, Эрексон, 1996).
Таблица 2
Состав пищи сеголетков белуги в первый месяц пребывания на пастбищах Северного Каспия, в % по массе
| Организмы |
1968 г. |
1974 г. |
1975 г. |
1976 г. |
| Mysidae |
100 |
100 |
93,3 |
35 |
| Gobiidae |
- |
- |
- |
65 |
| Прочие |
- |
- |
6,7 |
- |
| Всего |
100 |
100 |
100 |
100 |
У молоди белуги длиной до 10 см пища почти исключительно состояла из мизид. У сеголетков длиной 11-20 см 30 % пищевого рациона составляли бычки, главным образом, бычок-песочник. В разные гидрологические периоды спектр питания белуги менялся слабо. Рыба всегда составляла основу ее рациона.
Скатывающаяся молодь белуги (7,0-15,0 г) в реке питается бокоплавами, мизидами, личинками хирономид, поденок, ручейников, а также олигохетами, икрой и личинками осетровых и других рыб. Крупные мальки белуги поедают молодь севрюги и осетра. У молоди белуги может наблюдаться каннибализм при переходе на активное питание. Индексы наполнения желудков у молоди очень высокие - 280-400 0
/000
. Взрослые рыбы питаются преимущественно рыбой. Характер пищи белуги меняется по сезонам года и в зависимости от мест нагула. Известен случай нахождения в желудке белуги белька тюленя. Белуга косвенно связана с продукцией планктона и бентоса, так как питается рыбами-планктофагами и бентофагами (Полянинова, Эрексон, 1996).
Молодь белуги, выпушенная с заводов, попадая в новые для нее условия - реку, адаптировалась и, не теряя поискового инстинкта, сразу переходила на питание речными кормовыми организмами. В желудках молоди белуги, пойманной через 3 ч после выпуска с Бертюльского ОРЗ, отмечены как прудовые (стрептоцефалюс), так и речные (гаммариды) формы. По мере ската у Сергиевского ОРЗ и в нижерасположенных районах молодь белуги уже полностью перешла на питание речными кормовыми организмами.
Главными пищевыми объектами заводской молоди белуги в реке во все годы исследований служили гаммариды (от 50 до 90% массы пищевого комка). Индекс избирания по гаммаридам был равен 1. Из гаммарид наибольшую значимость в питании белуги имели N. abbreviates и N. maeoticus. Эти виды составляли от 40 до 93% массы пищевого комка и 60-100% по встречаемости. В отдельных районах (Сергиевский ОРЗ) в питании белуги возрастало значение N. similus - до 93% массы пищевого комка. У Четвертой Огневки среди компонентов пиши белуги отмечен N. compressus. Второстепенное значение в питании белуги имели мизиды и рыба (Молодцова, Полянинова, Смирнова, 1992).
На заводах кормление молоди осетровых до массы 3 г следует проводить крупкой стартового комбикорма ОСТ-4 (ОСТ-6). В связи с неустойчивым обеспечением комбикормовых заводов кормовым сырьем целесообразно использовать в качестве добавки в начале выращивания личинок и мальков, живые корма. Состав рецепта ОСТ-4 представлен в таблице 3.
Таблица 3
Состав рецепта стартового комбикорма ОСТ-4 для бестера, белуги и русского осетра
| Компоненты |
Содержание, % |
Заменители |
| 1 |
2 |
3 |
| Мука рыбная |
* |
Нет |
| Мука мясокостная |
* |
Рыбная |
| Мука кровяная |
* |
Рыбная |
| Мука пшеничная |
* |
Нет |
| Шрот соевый |
* |
Соевая мука, ПЗХ, витазар |
| Сухой обрат |
* |
Соевый шрот и др. продукты из сои, витазар, сухое молоко |
| Рыбные гидролизаты |
| Дрожжи |
Нет |
| Витазар |
* |
Фосфатиды |
| Масло подсолнечное |
* |
Нет |
| Жир рыбий |
* |
Нет |
| Премикс |
* |
| Протеин |
48-50 |
| Клетчатка |
1-1,6 |
| Углеводы |
14-18 |
| Жир |
8-11 |
| Общая энергия, мДж/кг |
18-19 |
В настоящее время в НТЦ «Астаквакорм» разработан новый рецепт стартового комбикорма ОСТ-6 с использованием рыбных гидролизатов, что позволяет отказаться от добавок в рацион личинок избытка живых кормов.
В составе стартового и продукционных кормов следует использовать специальный поливитаминный премикс ПО-5 или витаминно-минеральный премикс ВМП ПО-5, содержащий полный набор необходимых витаминов и микроэлементов. Эти премиксы разработаны НТЦ «Астаквакорм».
Норма ввода премиксов ВМП ПО-5 и ПО-5 в кормосмесь комбикормов равна 1%, заменителя - ПФ-2В -1,5%.
Свободным эмбрионам, вставшим на плав, начинают давать немного комбикорма ОСТ-4 в виде пыли еще до рассасывания пигментной пробки с целью выработки положительной пищевой реакции. После рассасывания пигментной пробки начинают давать крупку размером 50-100 микрон. Период адаптации к комбикорму длится 2-3 суток, одновременно с комбикормом следует кормить молодью дафний или артемии салина. Кормление молоди осетровых рыб олигохетами до массы 200-300 г не рекомендуется из-за возможного дисбаланса питательных веществ. При использовании добавок живых кормов кормление комбикормом не прекращают (табл. 4).
Таблица 4
Суточная норма кормления молоди осетровых рыб в зависимости от массы тела и температуры воды комбикормом ОСТ-4
| Масса тела, мг |
Суточная норма, % от массы тела рыб и температуры воды |
| 12-17°С |
17-20°С |
20-24°С |
24-27°С |
| До 60 |
30 |
35 |
35 |
30 |
| От 60 до 300 |
25 |
30 |
30 |
20 - |
| От 300 до 500 |
15 |
20 |
25 |
15 |
| От 500 до 1500 |
12 |
10 |
15 |
10 |
| От 1500 до 3000 |
10 |
8 |
12 |
8 |
В период повышения температуры воды летом до 30°С рекомендуется уменьшить норму дачи кормов на 50% и прекратить добавление жиров в комбикорм. Кратность дачи корма снижается в 2-3 раза. При использовании стартового комбикорма ОСТ-6 с рыбными гидролизатами живые корма не требуются или используются ограниченно.
Желательно для кормления молоди осетровых использовать небольшие автоматические кормораздатчики. Кратность кормления молоди в светлое время суток дана в таблице 5. Весьма эффективными являются ленточные (транспортерные) кормораздатчики непрерывного действия.
Таблица 5
Кратность кормления молоди осетровых рыб.
| Масса рыб, мг |
Ручное кормление |
Кормление с помощью автоматических кормораздатчиков |
| До 60 |
24 |
48 |
| От 60 до 300 |
12 |
36 |
| До 1000 |
8 |
24 |
| До 3000 |
6 |
12 |
Бионормативы кормления и выращивания молоди осетровых рыб массой до 3 г в бассейнах представлены в таблице 6.
По достижению рыбами массы 2 г добавку живых кормов в рацион прекращают и молодь переводят на кормление продукционным комбикормом рецепта ОТ-6, ОТ-7. Использование высокобелковых продукционных комбикормов обеспечивает высокий выход сеголетков, годовиков и двухлетков, оптимальный рост и удовлетворительное физиологическое состояние.
Выращивание крупного посадочного материала массой 500 г проводится в бассейнах типа ИЦА-2 или лотках. Содержание растворенного в воде кислорода должно быть не ниже 7 мг/л. Расход воды устанавливается в соответствии с оптимальным содержанием кислорода (8-10 мг/л). Расход воды в бассейнах для рыб массой от 3 до 500 г составляет 3-0,8 л/мин, на 1 кг рыбы, при недостатке кислорода он увеличивается. Смена воды происходит каж- дые 20- 25 мин., плотность посадки рыб массой 30-200 г составляет 500-400 шт./м2
.
При плотности посадки рыб 200-500 - 250-300 шт./м 2
уровень воды в бассейнах для рыб массой 30-500 г составляет 0,3-0,7 м.
Таблица 6
Бионормативы кормления и выращивания молоди осетровых рыб массой до 3 г в бассейнах
| Элементы биотехники |
Бионормативы |
| Глубина воды в бассейнах, лотках, м |
0,2-0,4 |
| Температура воды, °С, при выращивании: личинок мальков |
16-22
20-24
|
| Продолжительность выращивания молоди до массы Зг, сут. |
30-40 |
| Отход за период выращивания, % |
50 |
Кормовые затраты по комбикорму ОСТ-4 по живым кормам (сухое вещество)
Всего по кормам
|
0,8-1,2
0,5-1,2
1,3-2,4
|
Процесс выращивания посадочного материала массой 500 г может быть прерван вынужденной зимовкой. В период зимовки потеря массы может достигать 15%, отход - 10%.
Для кормления рыб массой от 3 до 500 г следует использовать продукционный комбикорм для осетровых рыб ОТ-6, а также его аналог ОТ-7 (с глютеном) (Пономарев, Гамыгин, Никоноров, и др., 2002).
Глава 5. Транспортировка икры, личинок, молоди и взрослых особей белуги
Интенсификация рыбоводства, расширение видового разнообразия аквакультуры, трансформация технологического процесса выращивания обусловливают необходимость совершенствования средств и методов транспортирования рыб определенных видов и возраста, а также живой икры как внутри хозяйства, так и за его пределами (Привезенцев, 2004).
По мнению Привезенцева Ю.А. (2004) в технологическом процессе выращивания рыбы в прудовом, хозяйстве предусмотрена перевозка рыбы разного возраста из одной категории прудов в другие. Товарная рыба в период ее выращивания в течение 2—3 лет подвергается 5—7 перевозкам. При закупке рыбопосадочного материала, ремонтного материала и производителей в племенных хозяйствах перевозку осуществляют в течение 2—3 суток. Процесс перевозки живой рыбы и икры является одним из сложных и ответственных в технологии выращивания, от которого во многом зависит успех отрасли. При перевозке живой рыбы из одного хозяйства в другое необходимо соблюдать следующие условия: получить разрешение ветеринарного надзора на право перевозки; подлежащую перевозке рыбу обработать дезинфицирующим раствором (солевым или аммиачным); запретить вывоз рыбы из хозяйств, неблагополучных по заболеваниям (краснуха, бранхиомикоз, фурункулез, вертеж лососевых, инфекционная анемия и дискокотилоз форели), рыбу, пораженную триходинозом, хилодонеллезом, дактиологирозом, допускать к перевозке после антипаразитарной обработки; при обнаружении заболевания во время транспортирования доставленную рыбу поместить в изоляторные пруды для лечения или направить на обработку; запретить сбрасывать в водоемы воду, использованную для перевозки рыбы.
Перевозка будет более благополучной при предварительном выдерживании рыбы в течение 2—10 ч без кормления в чистой проточной воде. Истощенная, побитая и вялая рыба плохо переносит транспортирование (Кожин, 1971).
Состояние рыбы при перевозке зависит от качества и объема воды. Необходимо емкости заливать чистой, насыщенной кислородом водой, не содержащей вредных и ядовитых веществ, температурой, равной температуре воды водоема, где выращивалась рыба. Оптимальной температурой для перевозки большинства теплолюбивых рыб в летнее время является 10—12 ° С, холодолюбивых 6—8 °С, а в осенне-весенний период — соответственно 5–6 и 3–5°С. Рыба, перевозимая при более низкой температуре воды, потребляет меньше кислорода, выделяет меньше продуктов обмена, а соответственно ее можно перевозить при более высокой плотности посадки в емкостях. Потребление кислорода в единицу времени также зависит от вида и возраста рыбы. Поэтому при перевозке рыбы важным показателем является соотношение между массой рыбы и объемом воды (Привезенцев, 2004).
Перевозка спермы. В связи с тем, что в семенной жидкости сперматозоиды находятся в активном состоянии, сперму рыб можно перевозить на любые расстояния в сухих стерильных пробирках, установленных в термосе со льдом. При этом необходимо учитывать сроки ее активности. Хранению и перевозке подлежит свежеотобранная сперма, помещенная в сухие пробирки отдельно для каждого самца с плотно пригнанными пробками во избежание попадания воды Пробирки с этикетками заворачивают в марлю и отпускают в термос, наполненный мелко наколотым льдом (Пономарева, Бахарева, 2002).
Перевозка неоплодотворенной икры. Икру закладывают в сухую банку, которую плотно закрывают пробкой и помещают в термос. Желательно иметь мелкую банку, чтобы она полностью была заполнена икрой, без свободного воздушного пространства (Пономарева, Бахарева, 2002).
Перевозка оплодотворенной икры. Икра белуги наиболее чувствительна к механическим воздействиям в стадии начала дробления, продолжающего 25—30 ч при температуре 11—15° С, в стадии гаструляции, продолжающейся 43—50 ч при температуре 11,5—14° С, и непосредственно перед выклевом. Икру осетровых (белуги) обычно перевозят в стадии пигментации глаз у зародышей. При температуре 11 —13° С эта стадия наступает через 60—80 ч после оплодотворения и продолжается 40—50 (до стадии появления S-образного изгиба на сердце зародыша. Для перевозки икры осетровых обычно используют обтянутые марлей деревянные рамки размером 34x28 см, разделение по полам планкой. Рамку выстилают влажной марлевой салфеткой размером 70x50 см. На салфетку в полтора-два слоя раскладывают икру. Вместимость рамки указанного размера 100 тыс. икринок осетровых.
Раскладку икры на рамки производят в воде, налитой плоский сосуд, размеры которого дают возможность свобод помещать рамку на поверхность воды. Чтобы вода не перегревалась, в нее добавляют лед. Рамки с икрой укладывают в стоги по 22—24 штук. Перед упаковкой икры на дно ящика устанавливают крестовину толщиной 4 см, на которую ставят подготовленную стопку рамок. Сверху стопку закрывают специальной рамкой, выстлан ной полиэтиленом. Рамка имеет бортики высотой 5 см. В эту рамку закладывают лед. Для закрепления стопки рамок внутрь ящика ее фиксируют деревянными брусками, а в свободное пространство закладывают лед. Во время транспортировки икры следят за тем, чтобы на дне ящика не скоплялось много воды. При необходимости воду сливают через спускные отверстия, находящиеся на его дне.
При поддерживании в изотермическом ящике постоянно температуры (4—6° С) длительные перевозки икры проходят обычно с незначительным отходом (2–5%).
За 3—4 ч до окончания транспортировки рекомендуется удалить из ящиков часть льда — это приведет к постепенному повышению температуры внутри транспортной тары. Таким образом, можно сократить до 1—1,5 ч время уравнивания температуры в транспортной таре и воды в инкубационном аппарате (Кожин, 1971).
Транспортировка личинок. Чаще всего для транспортировки личинок рыб используют стандартные полиэтиленовые пакеты объемом 40 л, длиной 65 см, с шириной рукава 50 см. Перед перевозкой их упаковывают в стандартную картонную коробку размером 65 х 35 х 35 см. В каждый полиэтиленовый пакет наливают 10 - 12 л воды, помещают личинок, свободное пространство заполняют кислородом и закрывают пакет с помощью зажима Мора или резинового шланга.
Оптимальной температурой для перевозки в летнее время теплолюбивых рыб является 10-12°С. В один пакет помещают, до 15 тыс. осетровых (белуги, осетра, севрюги). Транспортировку личинок рыб из инкубационного цеха можно также производить и в другой таре, например, в молочных флягах с крышкой. При температуре 4 - 5 °С и продолжительности транспортировки 2 часа допустимая плотность посадки 2-3 тыс. экз./л. После помещения личинок воду доливают до горловины фляг и плотно обвязывают двойным слоем марли, на марлю кладут деревянный брусок размером 2 х 2 см и опускают крышку, но не стягивают ее зажимом. Этим достигается постоянная аэрация воды и исключается выброс личинок с водой при толчках во время перевозки.
Перевозка личинок осетровых. Наилучшие результаты и минимальные отходы (0,1 — 1%) наблюдаются при перевозке одно–двухдневных личинок осетровых в течение 20—50 ч при температуре 10—18° С. Перевозка трех-четырехдневных личинок сопряжена с увеличением отходов до 3—10%.
Для перевозки личинок осетровых рыб используют канны из органического стекла, в которых воду аэрируют воздухом. Плотность посадки личинок (табл. 7) рассчитывают или по площади дна канны (4—7 шт/см2
), или по объему воды в ней (160—200 шт/л). В каннах емкостью 50—60 л размещают до 6—12 тыс. личинок средним весом 12—25 мг. Личинок перевозят в полиэтиленовых пакетах, наполненных водой и смесью воздуха с кислородом (рис. 47).
Таблица 7
Нормы посадки личинок осетровых рыб в полиэтиленовые пакеты емкостью 40—45 л
| на 5 тыс. личинок |
на 8-12 тыс. личинок |
| Воды, л |
15 |
15 |
| Кислорода, л |
10 |
15 |
| Воздуха, л |
10 |
15 |
При соблюдении указанных норм и температуре воды до 18°С перевозка личинок в течение 15—20 ч проходит обычно без отходов. Перевозка личинок в полиэтиленовых пакетах удобнее и рентабельнее, чем в каннах. В полиэтиленовых пакетах личинки можно отправлять прямыми рейсами самолетов без сопровождающих (Кожин, 1971)

Рис. 47. Полиэтиленовый пакет для перевозки личинок.
Перевозка живой рыбы в прорезях. Прорезь, предназначенную для перевозки живой рыбы, устанавливают на слабом течении реки или протоки. Рыбу загружают в прорезь с учетом общей ее массы. Для рыбоводных заводов и нерестово-выростных хозяйств разработаны нормы посадки живой рыбы в прорези (табл. 8).
Для предотвращения выпрыгивания рыбы из прорези ее закрывают матами из камыша или мелкоячейной дели, а сверху съемными крышками. Забивать прорезь досками не рекомендуется, так как при выпрыгивании рыба может получить тяжелые травмы.
Отсеки прорезей, в коррых содержится рыба, должны быть обтянуты пологами из мелкоячейной двух- или трехмиллиметровой капроновой ткани «рашель» или килечного капронового хамсороса.
Загруженную живой рыбой прорезь транспортируют к месту назначения катером со скоростью 7—10 км/ч (Кожин, 1971).
Таблица 8
Нормы посадки рыбы в прорези с полезным объектом 30 м3
| Вид рыбы |
Производители, шт. |
Молодь, тыс. шт. |
| Белуга |
5 |
50-60 |
| Осетр |
10 |
50-60 |
| Севрюга |
16 |
50-60 |
| Шип |
10-12 |
50-60 |
| Каспийский лосось |
40 |
| Белорыбица |
20 |
| Сазан |
1500-2000 |
500 - 600 |
| Леш |
2000-2500 |
1000-1500 |
| Судак |
800-1000 |
200-300 |
| Рыбец |
7000 - 7700 |
Транспортировка в живорыбных вагонах. Живорыбные вагоны используют для транспортировки рыбы на расстояния более 1000 км. Для осетровых рыб, которые постоянно держатся у дна, нормы посадки определяют по площади дна баков из расчета посадки их в один слой. В каждый живорыбный вагон помещают по 500 -800 экз. производителей осетра или севрюга при продолжительности транспортировки 4-8 суток и температуре воды 6 - 8 °С. Для поддержания в транспортировочных емкостях удовлетворительных условий содержания рыбы необходима постоянная аэрация воды. При длительных перевозках транспорт помимо основного компрессора должен иметь запасной автономный бензо-компрессор или запас баллонного кислорода. Оптимальным содержанием кислорода в воде во время перевозки рыбы считается 7 - 8 мг/л, его снижение до 3 мг/л свидетельствует о критическом состоянии рыбы. В большинстве специализированных транспортных средств предусмотрена термоизоляция. Перевозка рыбы в живорыбных вагонах может осуществляться при температурах воздуха от -40 до +30 °С (Пономарева, Бахарева, 2002).
Перевозка живой рыбы автотранспортом. На автомашинах рыбу перевозят на небольшое расстояние: доставка рыбы из живорыбных вагонов на рыбоводный завод; доставка молоди рыбы с рыбоводных заводов к месту выпуска; доставка производителей рыб с тоней или от прорезей на рыбоводный завод. Автомашины (ЗИЛ-150), приспособленные для перевозки живой рыбы, имеют баки емкостью до 3000 л, в которых поддерживается благоприятный для живой рыбы температурный и газовый режим. Обогащение воды кислородом происходит при прохождении ее через форсунки, к которым центробежный насос все время подает воду. Центробежный насос приводится в движение при помощи особого механизма от двигателя автомашины и может работать при движении и во время стоянки автомашины. Зимой вода в баке может подогреваться отработанными газами автомашины до температуры 5° С. При перевозке живой рыбы в пределах 50—100 км отношение количества рыбы к количеству воды колеблется около 1 : 1,5 или 1 :2. При более длительной перевозке норма загрузки должны быть снижена до 1 : 3 или даже 1 : 4 (на 1 кг рыбы 3—4 л воды зависимости от вида рыбы и времени перевозки. Живую рыбу на грузовой автомашине можно перевозить в брезентовых баках. Для этого на платформе машины устанавливают деревянный каркас в виде решетчатого ящика, размер которого зависит от размера платформы машины. Внутри деревянного каркаса навешивают брезент, который должен свободно выстилать внутренние стенки каркаса. В верхней части каркаса брезент прикреплен к крючкам решетки при помощи петель или колец. Таким образом, образуется большой кошель-бак, куда и наливают воду примерно до половины бака. Бак во время перевозки рыбы покрывают брезентовой крышкой, чтобы рыба с водой не выбрасывалась из бака (Кожин, 1971).
Глава 6. Биологические основы акклиматизации
Все работы по акклиматизации и зарыблению естественных водоемов и водохранилищ осуществляются в соответствии с Положением о порядке проведения работ по акклиматизации рыб, других водных организмов и зарыблению водоемов РФ под строгим контролем органов рыбоохраны. Мероприятиям по зарыблению и акклиматизации предшествует разработка научными организациями биологических обоснований, освещающих вопросы биологической и хозяйственной целесообразности их вселения. Положение о порядке проведения работ по акклиматизации рыб, других водных организмов и зарыблению водоемов РФ было утверждено приказом Председателя Комитета Российской Федерации по рыболовству 27 декабря 1993 года (Методические указания, 2002).
Акклиматизация водных организмов - это биолого-биотехнический процесс, представляющий собой вселение объектов, доставленных из одних водоемов, регионов, стран в другие, где их ранее не было или они исчезли, с целью их полной или частичной натурализации, а также других форм хозяйственного использования (спортивное рыболовство, озерно-товарное хозяйство, биологическая мелиорация). Акклиматизация проводится в двух формах: полноцикловая и поэтапная.
Полноцикловая акклиматизация - натурализация - это конечная фаза процесса акклиматизации, когда вселенец приспособился к новым условиям, определилась его ниша и взаимоотношения с аборигенами в экосистеме заселяемого водоема, установилось подвижное равновесие численности новой популяции, и появилась возможность ее использования в кормовых и промысловых целях.
Поэтапная акклиматизация - не завершенная акклиматизация, когда некоторые этапы развития вселенца не могут завершиться в условиях водоема вселения и проходят в других водоемах или под протекцией человека.
Зарыбление - частный случай поэтапной акклиматизации, характеризующий тип, метод хозяйственной деятельности, и предусматривающий регулярный выпуск посадочного материала на нагул в уже апробированные водоемы.
Реакклиматизация - интродукция особей вида в целях восстановления популяции в пределах его естественного (в прошлом) ареала, в котором этот вид по каким-либо причинам исчез (Методические указания, 2002).
В текущем этапе курсовой работы я хотел бы акклиматизировать белугу для выращивания в водоемы лиманного типа (лагуны) находящиеся в Азовском море.
Азовское море конечный самый малый мелководный бассейн эстуарного типа в цепи средиземноморских морей. Площадь моря приблизительно 38 тыс. км2
, объём равен 320 км 2
, глубина ср ~ 8,4 м, глубина max =13,6 м. Благодаря малым глубинам оно быстро прогревается летом и охлаждается до дна зимой. Экологическая ёмкость Азовского моря была ограничена, а биотическая довольно велика. Химическая основа кормности этого бассейна обеспечивается систематическим поступлением биогенов извне и внутренними их запасами, накопленными в организме, грунте и толще воды. Из рек Дона, Кубани и других до 1952 года поступало ежегодно около 80 тысяч тонн азотистых соединений и 12-13 тысяч тонн фосфорных. Экологическая емкость моря зависит от гидрологического режима, а биотическая сохраняется высокой. Разрыв между экологической и биотической емкостью приводит к недоиспользованию органического вещества кормов.
Необходимые условия для вселения имеются в Ахтанизовском и Курчанском лиманах.
Ранее такие работы уже проводились учеными АзНИИРХа. Для проверки эффективности использования лиманов в Ахтанизовский лиман в 1973 г. была вывезена заводская молодь осетровых выращенная в прудах Темрюкского осетрового завода, в следующем количестве: осетр 250 тыс. шт., белуга 250 тыс. шт., севрюга 200 тыс. шт.
Изучение биологии молоди в лимане показало высокую эффективность этого метода. Хищные рыбы молодью осетровых не питались. Кормовая база полностью обеспечивала интенсивное питание осетровых. Осетр и севрюга питались ракообразными (мизиды и корофииды), белуга потребляла бычков.
В 1973 г. вес белуги увеличился в среднем с 8,6г при посадке до 680,0 г в конце сезона, осетра с 4,8 г до 90,0 г, севрюги с 4,2 г до 86,0 г.
Для определения количества молоди, обитающей в лимане, были организованы съемки лимана, и наблюдения в гирле за выходом молоди из лимана. По сравнению с прямым выпуском в море численность осетровой молоди в лимане оказалась в 4 раза выше (Бокова, и др., 1979).
Гидрологические, гидрохимические показатели и кормовая база в Курчанском лимане мало, чем отличаются от сих показателей в Ахтанизовском. Из этого следует, что Курчанский лиман может служить водоемом вселения.
Летом в открытой части лимана температура воды на некоторых участках достигает 25 – 27˚С, а у берегов повышается до 30˚С. Зимой вода охлаждается до 0˚С и образуется лед. Соленость колеблется от 13 до 16 ‰. Эти колебания зависят от чередования многоводных и маловодных лет (Анисимова, и др., 1983).
Кормовая база выше описанного водоема полностью устраивает потребности белуги при интенсивном питании. А это значит что затраты на корм (искусственный корм, вселение кормовых объектов) не требуются. Что значительно увеличивает экономическую ценность этой попытки.
Молодь белуги для вселения можно брать на рыбоводных заводах Дона и Кубани.
Сроки вселения должны совпадать с началом интенсивного развития и размножения кормовых объектов, которое приходится на июнь – июль.
По истечению трех лет белуга наберет товарную массу (2−2,5 кг) и можно будет организовывать промысел.
Данный вид вселения относится к частному случаю поэтапной акклиматизации − зарыблению, который описан в начале главы.
Список литературы
1. Анисимова И.М., Лавровский В.В. Ихтиология. − Москва. − изд-во «Высшая школа», 1983. − 255 с.
2. Атлас пресноводных рыб России в двух томах. Т–1– под редакцией Решетникова Ю.С. М. наука, 2002. – 253 с.
3. Бабушкин Н.Я. 1964. Биология и промысел каспийской белуги // Тр. ВНИРО. Т. 52. Сб.1. М.: с. 183-258.
4. Бокова Л.И., Гунько А.Ф. опыт использования лиманов для выращивания заводской молоди осетровых в Азовском бассейне // Осетровое хозяйство внутренних водоемов СССР.−1979.− с. 28-29.
5. Вернидуб М.Ф. Инструкция по определению рыбоводного качества, сбору, оплодотворению, рационализации обесклеивания и инкубации икры и выдерживанию личинок осетровых. − рукопись, 1952.
6. Детлаф Т.А., Гинзбург А.С. Зародышевое развитие осетровых рыб (севрюги, осетра и белуги) в связи с вопросами их разведения. − Москва. − изд-во Академии наук СССР, 1954.−215 с.
7. Драгомиров Н.И. Эколого-морфологические особенности личиночного развития белуги HUSOHUSO (L.). // Труду института морфологии животных им. А. Н. Северцова. − 1961. − вып. 33. − с. 72-185.
8. Захарова Н.А., Хураськин Л.С., Полянинова А.А. 2000. Трофические связи осетровых и каспийского тюленя // Осетровые на рубеже 21 века: Тез. докл. Междунар. конф. Астрахань: Изд-во КаспНИРХа. 54 с.
9. Иванов А.П. Рыбоводство в естественных водоемах. - М.: Агропромиздат, 1988. - 367 с.
10. Кокоза А.А., Искусственное воспроизводство осетровых рыб. Моногр.− Астрахань. − изд-во АГТУ, 2004. −207 с.
11. Курс лекций: «Осетровые мирового океана». Для студентов третьего курса специальности «Водные биоресурсы и аквакультура». Л.В. Витвицкая, А.М. Тихомиров, Н.А. Егоров; АГТУ–Астрахань, 2002.–160 с.
12. Легеза М.И. 1972. Роль абиотических факторов среды при распределении осетровых (сем. Acipenseridae, Pisces) в Каспийском море // Вопр. ихтиологии. Т. 121. Вып.1. С. 13-24.
13. Методическое указание на тему: Биологические основы акклиматизации. − Астрахань. − изд-во АГТУ, 2002.
14. Мильштейн В.В. Осетроводство.: М. агропромиздат 1982.
15. Молодцова А.И. Поляникова А.А. Смирнова. А.В. Кормовая база и питание заводской молоди белуги в естественных условиях реки Волги. //Воспроизводство осетровых, лососевых и частиковых рыб: сб. научных трудов ВНИРО. М. 1992 – с 100–110
16. Мухачев И.С. Биологические основы рыбоводства: учеб. пособие: М–во. образования и науки. Р. Ф. Тюмень. Гос. университет – Тюмень из–во. Тюмень гос. ун–та, 2005– 300 с.
17. Поляникова А.А. Эриксон. Е.П. Питание белуги в Каспийском море в современных условиях // Тез. докл. 6–й Всероссийской конфиренции по проблемам промысла. Мурмнск 1996 с. 119–120
18. Понамарев С.В. Гамыгин Е.А., Никоноров С.И. Понамарева Е.Н. Бахарева А.А. Грозеску Ю.Н. Технологии выращивания и кормления объектов аквакультуры юга России.– Астрахань: «Нова плюс», 2002
19. Понамарева Е.Н. Бахарева А.А. Методы стимулирования половых продуктов у рыб различными способами. Астрахань 2001
20. Понамарева Е.Н. Бахарева А.А. Методы транспортировки икры, спермы, личинок, молоди и производителей рыб. Астрахань 2002
21. Привезенцев Ю.А. Рыбоводство: учебник для студентов вузов/ Ю.А. Привезенцев, В.А. Власов. – М.: Мир, 2004. – 456 с.
22. Рыбы Казахстана.1986. Т. 1. Алма-Ата: Наука. С. 57-71.
23. Рыбохозяйственные исследования на Каспии: Результат НИР за 2005г Романов А.А., Журавлёва О.Л., Ходоревская Р.П., Левин А.В., Лепилина И.11., Коноплёва И.В., Сафаралиев И.А. Астрахань.: Изд–во Капнирх. 2006– 436 с
24. Справочник рыбовода по исскуственному разведению промысловых рыб/ под ред. д. б. н. проф. Н.И. Кожина–М. Пищевая пром.1971–208 с.
25. Ходоревская Р.П., Полянинова А.А. 2000. Оценка условий нагула белуги (Huso huso L.) в северо-западной части Северного Каспия // Морские гидробиологические исследования. М.: ВНИРО. С. 205-208.
26. Черномашенцев. А.П. Мильштейн. В.В. Учебник рыбоводства 1 часть: М. Агропромиздат 1983.
|