Содержание
Биполярные, горизонтальные и амакриновые клетки
Медиаторы в сетчатке
Концепция рецептивных полей
Ответы биполярных клеток
Структура рецептивных полей биполярных клеток
Горизонтальные клетки и ингибирование периферии
Значение структуры рецептивных полей биполярных клеток
Рецептивные поля ганглиозных клеток
Использование дискретных зрительных стимулов для определения рецептивных полей
Организация рецептивных полей ганглиозных клеток
Выводы
Литература
Общая схема связей между клетками сетчатки была составлена на основе морфологических описаний Рамон-и-Кахаля: проводящий путь от фоторецепторов до ганглиозных клеток при помощи биполярных клеток, с регулированием передачи посредством горизонтальных и амакриновых клеток. Исходя из схемы соединений клеток в сетчатке приматов становится понятным, что конечный выходной сигнал является результатом сложного интегративного процесса, происходящего в самой сетчатке. Например, горизонтальная клетка, показанная на рис.2D, получает синаптические входы от многих фоторецепторов и, в свою очередь, образует с ними модулирующую обратную связь.
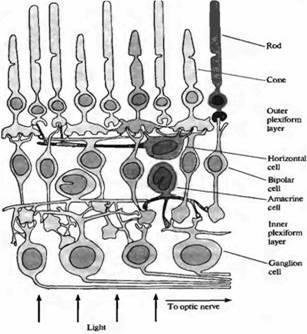
Рис. 1. Основные клеточные типы и соединения в сетчатке приматов представлены здесь для иллюстрации связей между фоторецепторами (палочками и колбочками) и ганглиозными клетками.
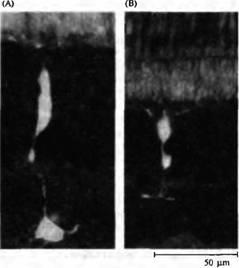 
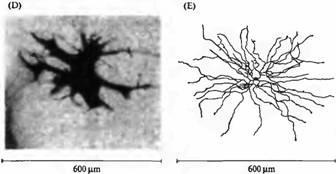
Рис. 2. Биполярные, горизонтальные и амакри новые клетки. (А) Деполяризующаяся биполярная клетка "оп"-центра золотой рыбки с введенным флуоресцентным красителем. (В) Гиперполяризующаяся биполярная клетка "off''-центра золотой рыбки. (С) Выделенная биполярная клетка из сетчатки крысы, окрашенная на про теинкиназу С. (D) Горизонтальная клетка акулы (морской собаки) с введенной пероксидазой хрена. (Е) Накапливающая индоламин амакриновая клетка из сетчатки кролика с введенным красителем Lucifer Yellow.
Горизонтальные клетки также образуют окончания на биполярных клетках. Аналогично некоторые амакриновые клетки (рис.2Е), получающие сигналы от биполярньи клеток, посылают аксоны как обратно на биполярные, так и на ганглиозные клетки. Можно сделать вывод, что горизонтальные и амакриновые клетки осуществляют и модулируют процесс передачи сигнала в сетчатке. Дополнительную сложность создает то, что каждый из основных классов нейронов, показанных на рис.1 и 2, имеет множество морфологических и фармакологических подтипов).
При помощи электрофизиологических, биохимических и анатомических критериев было описано несколько основных классов биполярньи клеток, больше чем 2 типа горизонтальных клеток и по крайней мере 20 типов амакриновых клеток.
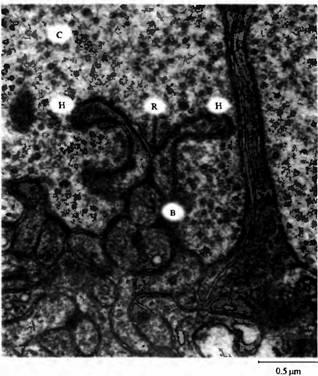
Рис. 3. Лентовидные синапсы, образованные терминалью фоторецептора на окончаниях биполярных и горизонтальных клеток. Пресинаптические везикулы в терминалях колбочек (С) выстроены вдоль ленты (R). Данный тип синапсов приспособлен к постоянному высвобождению квантов глутамата к биполярным (В) и горизонтальным (Н) клеткам в темноте. Через ГАМК-ергические горизонтальные клетки осуществляется обратная связь с фоторецепторами.
В сетчатке были обнаружены практически все известные медиаторы. Глутамат высвобождается фоторецепторами и биполярными клетками. Горизонтальные клетки секретируют ГАМК. Некоторые амакриновые клетки секретируют дофамин, другие ацетилхолин или индоламины. Распределение и функциональная значимость этих медиаторов, рецепторов и их субъединиц, а также переносчиков сигнала были детально изучены методами иммуногистохимии и гибридизации in situ. Некоторые медиаторы-пептиды, например VIP (вазоактивный интестинальный пептид), обнаруженные в глазу, могут играть важную трофическую роль в развитии зрительной системы и быть связаны с развитием миопии (близорукости).
Постоянное высвобождение квантов глутамата фоторецепторами в темноте осуществляется при помощи специального типа нервных окончаний, называемых лентовидными синапсами (ribbon synapses). Пример такого синапса показан на рис.3. Терминаль колбочки содержит везикулы, плотно связанные с мембраной и готовые к высвобождению, которые расположены вдоль длинной плоской органеллы, называемой "лентой" (ribbon). Постсинаптическая мембрана образована терминалями биполярных и горизонтальных клеток, которые формируют инвагинации на окончаниях фоторецепторов.
Техника освещения определенных зон сетчатки привела к появлению такой важной концепции, как рецептивное поле, которая стала ключом к пониманию значения сигналов не только на уровне сетчатки, но и на последующих стадиях в коре. Сам термин "рецептивное поле" был исходно введен Шеррингтоном по отношению к рефлексам и был перенесен в зрительную систему Хартлайном.
Рецептивное поле нейрона зрительной системы может быть определено как зона сетчатки, при падении света на которую может изменяться активность данного нейрона.
Например, регистрация активности одного нейрона зрительного нерва и коры кошки показала, что частота его импульсации увеличивается или уменьшается, если меняется освещение над определенной зоной сетчатки.
Эта зона является рецептивным полем данного нейрона.
---------
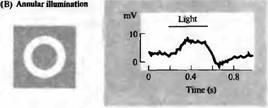
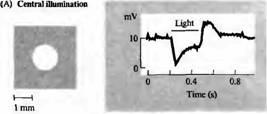
Рис. 4. Карта рецептивных полей биполярных клеток, чувствительных к гиперполяризации (Н). Сигналы регистрировались на биполярной клетке сетчатки золотой рыбки, которая отвечала гиперполяризацией (А) на освещение центра рецептивного поля. Освещение в виде кольца вызывало деполяризацию клетки (В). Рассеянный свет слабо влиял на потенциал клетки. Для биполярной клетки, чувствительной к деполяризации (D), освещение центра рецептивного поля приводит к деполяризации, в то время как освещение в виде кольца вызывает гиперполяризацию.
По определению освещение за пределами рецептивного поля не оказывает влияния на импульсацию. Эта зона сама по себе может быть подразделена на несколько различных регионов, освещение некоторых из которых увеличивает активность, а других подавляет. Это описание рецептивных полей также применимо по отношению к другим нейронам, таким как биполярные и горизонтальные клетки, у которых локальный ответ переменной амплитуды вызывается падением света на сетчатку (что обсуждается далее в этой главе). Как будет показано далее, отдельные вспышки ненаправленного света практически бесполезны при оценке функции зрительной системы.
Каждая биполярная клетка получает сигналы непосредственно от палочки либо от колбочки. Клетки обычно получают сигналы от 15-45 рецепторов. Существует особый тип биполярных клеток, так называемые "карликовые биполяры" (midget bipolar), которые получают сигнал только от одной колбочки. Логично ожидать, что большинство карликовых биполяров находится в области ямки, где острота зрения максимальна. Они передают сигналы с фоторецептора на специальный тип ганглиозных клеток. В других отделах сетчатки биполярные клетки собирают информацию с 15-20 колбочек. Биполярные и горизонтальные клетки отвечают на освещение градуальной де - или гиперполяризацией.
Сигналы и рецептивные поля биполярных клеток зависят от двух механизмов. Во--первых, постоянное высвобождение медиатора в темноте способствует поддержанию ряда биполярных клеток в состоянии постоянной деполяризации, а других - в состоянии гиперполяризации, в зависимости от того, какие - возбуждающие или тормозные - рецепторы они имеют. Во-вторых, свет приводит к гиперполяризации фоторецепторов, тем самым уменьшая выброс глутамата. Уменьшение постоянного освобождения глутамата фоторецепторами при освещении приводит, соответственно, к уменьшению активности биполярных клеток, имеющих возбуждающие рецепторы глутамата, т.е. вызывают их гиперполяризацию. Такие клетки называются Н-биполяры (hyperpolarizing).
Наоборот, уменьшение тонического высвобождения глутамата фоторецепторами при освещении последних приводит к деполяризации тех биполярных клеток, которые экспрессируют тормозные рецепторы глутамата - это D-биполяры (depolarizing). D-биполярные клетки являются одним из редких типов клеток, у которых были обнаружены тормозные эффекты, опосредуемые через рецепторы глутамата. Канеко с коллегами показали, что данный тормозный эффект обусловлен метаботропными рецепторами глутамата, расположенными на мембранах биполярных клеток, которые действуют через G-белок и вторичные посредники, приводя к уменьшению ионной проводимости. Таким образом, в темноте в результате активации этих рецепторов ионные каналы, управляемые циклическими нуклеотидами на D-биполярных клетках, закрываются (в отличие от каналов фоторецепторов). После освещения высвобождение глутамата уменьшается, открываются катионные каналы, и биполярные клетки деполяризуются.
Рецептивное поле гиперполяризующейся Η-биполярной клетки показано на рис.4. Небольшой пучок света, направленный на центральную область поля, вызывает длительную гиперполяризацию. Освещение в форме кольца с темной зоной в центре приводит к деполяризации. Таким образом, рецептивная область подобной клетки состоит из различных по светочувствительности центральной зоны и окружающей ее периферии. Н-биполярная клетка, показанная на рис.4, может быть описана как клетка, имеющая рецептивное поле с "off"-центром, поскольку она деполяризуется, когда поток света исчезает.
D-биполярная клетка имеет рецептивное поле сходной концентрической формы, но освещение центра ее поля приводит к деполяризации, а освещение в форме кольца - к гиперполяризации. Из-за того, что она деполяризуется, когда включается свет, считается, что D-биполярная клетка имеет рецептивное поле с "on"-центром. Терминология "on" - и "off"-клеточных ответов будет активно использоваться нами далее при описании свойств рецептивных полей на следующих этапах зрительной системы. Важным принципом является то, что отдельные фоторецепторы могут одновременно находиться в центре одних рецептивных полей, контролируемых "on" - либо "off"-биполярной клеткой, в то же самое время являясь периферией в других рецептивных полях.
Ответы D - и Η-биполярных клеток при освещении периферии рецептивных зон опосредуются горизонтальными клетками. Каждая горизонтальная клетка получает информацию от большого количества фоторецепторов. Горизонтальные клетки, подобно Н-биполярным клеткам, отвечают на освещение фоторецепторов гиперполяризацией (из-за того, что при освещении уменьшается выделение фоторецепторами глутамата). Другой особенностью горизонтальных клеток является то, что они связаны друг с другом посредством электрических синапсов. Краситель люцеферовый желтый (Lucifer yellow), введенный в одну горизонтальную клетку, распространяется через щелевые контакты в другие нейроны. Таким образом, каждая горизонтальная клетка получает влияние с большой зоны сетчатки, иннервируемой ею и ее ближайшими соседями.
Горизонтальные клетки образуют с биполярными и фоторецепторными клетками тормозные синаптические соединения, в которых медиатором служит ГАМК. Таким образом, деполяризации фоторецепторов в темноте противостоит тормозное влияние горизонтальных клеток. Освещение фоторецепторов приводит к гиперполяризации горизонтальных клеток, и, следовательно, к уменьшению выделения ими ГАМК. Получается, что гиперполяризации фоторецепторов рассеянным светом противостоит соответствующее уменьшение секреции ГАМК в синапсах между фоторецептором и горизонтальной клеткой.
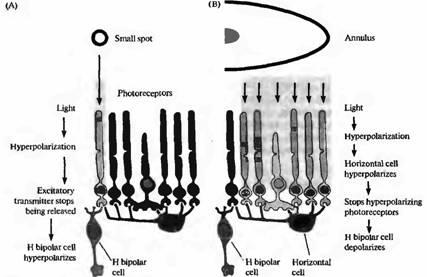
Рис. 5. Соединения между фоторецепторами биполярными и горизонтальными клетками. (А) Свет, падая на отдельную фоторецепторную клетку, вызывает ее гиперполяризацию, результатом чего является прекращение выделения глутамата В результате H биполярная клетка, как показано на рис. 19.18, гиперполяризуется, уменьшая свое возбуждение. (В) Свет, падающий на соседние области в форме кольца, также приводит к тому, что уменьшается высвобождение глутамата фоторецепторами. В результате происходит гиперполяризация горизонтальной клетки, которая предотвращает высвобождение ею тормозного медиатора ГАМК на фоторецептор. Следовательно, фоторецептор, который соединен с Η-биполярный клеткой, деполяризуется (из-за уменьшения тормозного влияния), что вновь приводит к высвобождению медиатора и деполяризации биполярной клетки. При освещении рассеянным светом процессы гиперполяризации и деполяризации гасят друг друга. Таким образом, горизонтальные клетки играют ведущие роли для обеспечения характерных свойств рецептивных полей биполярных клеток.
Было высказано предположение, что определенную роль здесь играет оксид азота, синтезируемый как фоторецепторами, так и горизонтальными клетками, которые также вносят вклад в угнетение высвобождения глутамата фоторецепторами. В итоге отрицательная обратная связь между фоторецепторами и горизонтальными клетками происходит по следующей схеме: свет. > гиперполяризация фоторецепторов > гиперполяризация горизонтальных клеток > деполяризация фоторецепторов.
Соединения клеток в случае центрального "on" - и периферического "off"-ответов биполярных клеток схематично показаны на рис.5. Для простоты центральная зона представлена всего одним фоторецептором, а периферия - несколькими соседними фоторецепторами, связанными с одной горизонтальной клеткой.
Сигнал в ответ на освещение центрального фоторецептора очевиден (рис.5А). Активация фоторецептора приводит к его гиперполяризации и, следовательно, к уменьшению высвобождения глутамата, что вызывает гиперполяризацию биполярной клетки. Горизонтальная клетка также получает стимулы к гиперполяризации, однако, так как они поступают только с одного фоторецептора, они имеют небольшой эффект, проявляющийся только в виде отрицательной обратной связи на центральный фоторецептор.
Ответы на освещение периферии (рис.5В) вовлекают дополнительные механизмы. Горизонтальная клетка, получающая сигналы от нескольких фоторецепторов вокруг центральной зоны, при освещении гиперполяризуется. Гиперполяризация уменьшает высвобождение ГАМК горизонтальными клетками, что приводит к уменьшению тормозного влияния на фоторецепторы и к их деполяризации. Этот эффект деполяризации проявляется минимально по отношению к фоторецепторам периферии, так как они сами по себе гиперполяризованы освещением. Однако центральный фоторецептор не находится под освещением, и поэтому единственным влиянием на него является уменьшение ингибирования со стороны горизонтальной клетки. В результате этого центральная клетка деполяризуется, увеличивается высвобождение ею глутамата, что все вместе приводит к деполяризации Η-биполярной клетки. В обзорах подробно описываются свойства терминалей фоторецепторов, терминалей биполярных клеток, а также синапсы обратной связи между горизонтальными и биполярными клетками.
Какое же физиологическое значение имеет такая структура рецептивных полей биполярных клеток?
D - и Η-биполярные клетки не просто возбуждаются в ответ на освещение. Они начинают анализ информации об элементарных паттернах света. Сигналы биполярных клеток несут информацию о различных падающих на сетчатку пятнах света, окруженных темным полем, либо, наоборот, о темных пятнах, окруженных светом, т.е. они отвечают на контрастные паттерны света и темноты в маленькой области сетчатки.
В дополнение к уже известным нам большим категориям D - и Η-биполярных клеток имеется еще около 11 типов различных биполярных клеток, связанных с колбочками, которые различаются по морфологическим и иммуногистохимическим критериям. Базируясь на них, можно получить три принципиальных типа клеток:
1. Связанные с колбочками D - и Н-биполярные клетки, лучше всего отвечающие на мельчайшие пятна света или темноты.
2. D - и Η-карликовые биполярные клетки центральной области сетчатки, образующие связи только с одной колбочкой.
3. Биполярные D-клетки, связанные с палочками центрального "on"-ответа, лучше всего реагирующие на малые яркие световые пятна.
Эфферентные сигналы сетчатки.
Еще за много лет до того, как от фоторецепторов и биполярных клеток сетчатки могли быть отведены электрические ответы, важная информация была получена при регистрации сигналов от ганглиозных клеток. В этих экспериментах был осуществлен первый анализ выходных сигналов в сетчатке, являющихся результатом происходящих там синаптических взаимодействий. Преимуществом изучения ганглиозных клеток является то, что их сигналы являются потенциалами действия и функционируют по принципу "все или ничего". Поэтому было возможно проведение регистрации сигнала при помощи внеклеточного электрода в то время, когда внутриклеточные электроды еще не существовали и пока не были разработаны соответствующие красители. Кроме того, простыми и удобными для изучения делало их то, что отростки ганглиозных клеток направляются из сетчатки в ЦНС. Именно на них были впервые описаны концентрические поля с "on" - и "off"-центрами. И именно они помогли потом понять смысл сигналов, регистрируемых на горизонтальных и биполярных клетках.
Стивен Куффлер был первым, кто выполнил экспериментальные исследования зрительной системы млекопитающих, обратив внимание на структуру рецептивных полей и их значение для зрительной сигнализации у кошки. В его экспериментах главный интерес состоял в конечных результатах синаптических взаимодействий, нежели в самих синаптических механизмах. Хьюбель впоследствии высоко оценил перспективы таких исследований:
Что особенно для меня интересно - это неожиданность результатов, поскольку никто до Куффлера не дошел до предположения о существовании рецептивных полей в виде структуры "центр - периферия" и что зрительный нерв фактически игнорирует любой раздражитель в виде рассеянного света любой интенсивности.
Принципиально новый подход был обусловлен не только новой техникой исследования, скорее, он обусловлен четкой формулировкой следующего вопроса: каким образом лучше всего стимулировать отдельную ганглиозную клетку? Ответ на этот вопрос привел к использованию отдельных кольцевидных световых пятен для стимулирования отдельных зон сетчатки вместо однородного диффузного освещения.
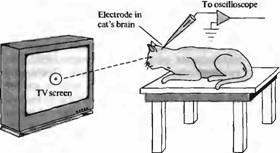
Рис. 6. Стимуляция сетчатки при помощи различных световых образов. Адаптировавшиеся глаза кошки или обезьяны, находящейся под наркозом, направляют в сторону кино - или телевизионного экрана, на котором показаны различные световые образы, генерируемые компьютером и/или спроецированные проектором. При этом при помощи электрода регистрируют ответы отдельного нейрона путей зрительного анализатора. Световые пучки, возникающие в определенной области экрана, увеличивают или уменьшают частоту разрядов данного нейрона. Таким образом может быть очерчено рецептивное поле этой клетки при помощи определения границ зон на экране, представление которых вызывает изменение частоты импульсации. В оригинальных экспериментах Kuffler направлял свет прямо в глаз при помощи специально сконструированного офтальмоскопа.
Эти исследования предвосхитили пионерские работы по исследованию глаза простых беспозвоночных: мечехвоста Limulus и сетчатки лягушек.
Изначальный выбор Куффлером глаза кошки был удачным: у кролика, к примеру, ситуация гораздо более сложная. Ганглиозные клетки сетчатки кролика имеют рецептивные поля, тонко реагирующие на такие сложные элементы, как края светового пятна и определенное направление его движения. Также сложны эти механизмы и у низших позвоночных, таких как лягушки. Можно выявить общую закономерность: чем глупее животное, тем умнее у него сетчатка (Д.А. Бейлор, личное сообщение).
Главной особенностью ранних экспериментов Куффлера было использование интактного не рассеченного глаза, нормальная рефракция которого использовалась для стимуляции определенных зон сетчатки. Удобным методом освещения отдельных отделов сетчатки является общая анестезия животного и помещение его перед телевизионным экраном на расстоянии, на котором его глазом осуществляется нормальная рефракция. При этом при появлении на экране монитора отдельных вспышек, светящихся паттернов или созданного компьютером изображения оно хорошо фокусируется на поверхности сетчатки (Рис.6).
При регистрации сигналов от одной ганглиозной клетки прежде всего важно найти границы ее рецептивного поля. Характерной особенностью большинства ганглиозных клеток и нейронов зрительной системы является то, что они периодически дают редкие разряды в покое, даже при отсутствии освещения. Соответствующие световые стимулы не обязательно вызывают активизацию ганглиозных клеток, они могут просто модулировать спонтанную импульсацию; при этом ганглиозные клетки отвечают либо увеличением, либо уменьшением частоты сигналов.
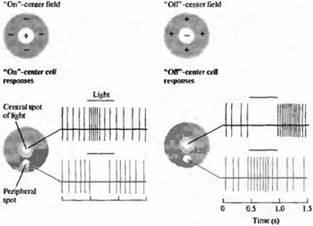 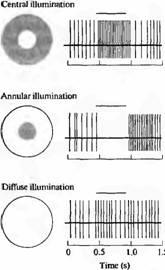
Рис. 7. Рецептивные поля ганглиозных клеток сетчаток кошки и обезьяны, сгруппированные в два основных класса: поля с "on" и поля с "off"-центрами. Клетки с "on"-центральными полями наиболее чувствительны к пятнам света, проецирующимся на центральную зону рецептивного поля. Освещение (показано в виде полоски над сигналами) окружающей области вместе с пятном, а также освещение в виде кольца уменьшают или полностью подавляют разряды, появляющиеся вновь при выключении света. Освещение всего рецептивного поля вызывает слабые ответы из-за того, что центр и окружение проявляют антагонистические эффекты вместе с биполярной клеткой. Клетки с "off" центральными полями уменьшают или прекращают посылать сигналы при освещении центральных зон их рецептивных полей и активируются при выключении света. Свет, падающий на окружающие зоны рецептивных полей с "off"-центром вызывает возбуждение нейрона.
На рис.7 показано, что для ганглиозной клетки маленькое пятнышко света 0,2 мм в диаметре, спроецированное в определенную зону рецептивного поля, способно гораздо лучше вызывать возбуждение, чем рассеянный свет. Более того, то же самое пятнышко света может вызывать и противоположные эффекты, в зависимости от его положения в пределах рецептивного поля. Например, одна зона пятна света возбуждает ганглиозную клетку в течение всего времени освещения. Такого типа "on"-ответ может быстро перейти в "off"-ответ при помощи простого смешения пятна вдоль поверхности сетчатки на 1 мм или менее.
Как и в случае биполярных клеток, существуют два основных типа рецептивных полей ганглиозных клеток: имеющие рецептивные поля с "on" - и "off"-центрами. Рецептивные поля обоих типов имеют приблизительно концентрическую форму.
В случае рецептивных полей с "on"-центром свет вызывает наиболее сильный ответ в том случае, если световое пятно полностью заполняет центр, в то время как для наиболее эффективного угнетения импульсации свет должен покрыть всю периферию в виде кольца (случай кольцевидного освещения показан на рис.7). При выключении угнетающего активность кольцевидного освещения ганглиозная клетка отвечает дополнительным "off"-разрядом. Рецептивные поля с "off"-центром имеют противоположное строение: угнетение происходит при освещении центральной зоны. Для обоих типов зон эффекты света на центр и на периферию антагонистичны. Таким образом, если и центр, и периферия одновременно освещаются, эффекты стремятся погасить друг друга
Палочки и колбочки способны реагировать на освещение ярким и слабым светом.
Зрительные пигменты плотно упакованы в мембранах палочек и колбочек.
Передача сигнала происходит в несколько этапов, с участием G-белка и иГМФ.
В темноте фоторецепторы деполяризованы и постоянно высвобождают медиатор глутамат.
Свет приводит к закрытию управляемых цГМФ катионных каналов, гиперполяризации и уменьшению высвобождения глутамата.
Два основных класса биполярных клеток реагируют на глутамат, высвобождаемый фоторецепторами.
Η-биполярные клетки деполяризованы в темноте и гиперполяризуются на свету.
D-биполярные клетки гиперполяризованы в темноте и деполяризуются на свету.
Рецептивные поля соответствуют зрительным полям или участкам сетчатки, освещение которых вызывает сигналы в клетках зрительной системы.
Фоторецепторы, горизонтальные клетки и биполяры не способны генерировать потенциал действия.
Ганглиозные и амакриновые клетки генерируют потенциал действия.
Биполярные и ганглиозные клетки имеют концентрические рецептивные поля, с "on" - и "off"-центрами и противоположной по функции периферией.
Ганглиозные клетки плохо отвечают на рассеянный свет.
Большие ганглиозные клетки, известные как magnocellular или М-клетки, имеют большие рецептивные поля и хорошо отвечают на движение.
Малые ганглиозные клетки, называемые parvocellular или Р-клетки, имеют небольшие рецептивные поля и способны воспринимать цвет и тонкие детали
1. Baylor, D. 1996. How photons start vision. Proc. Nail. Acad. Sci. USA 93: 540-565.
2. Boycott, В., and Wàssle, H. 1999. Parallel processing in the mammalian retina: The Proctor Lecture. invest. Ophihalmol. Vis. Sci.40: 1313-1327.
3. Dowling, J. E. 1987. The Retina: An Approachable Part of the Brain. Harvard University Press,Cambridge, MA.
|