КУРСОВАЯ РАБОТА
"Прочно связанные с целлюлозой полисахариды клеточной стенки ксилемы льна"
Введение
Клеточная стенка – один их немногих клеточных компартментов, наличие которого принципиально отличает растительную клетку от животной. Казалось бы, она должна пользоваться особым вниманием биологов растений. Однако нет, пожалуй, другого столь масштабного и столь плохо понятого процесса как формирование клеточной стенки. Причин для такого положения вещей несколько. Важнейшая из них – сложность, ранее недооцененная, этой надмолекулярной структуры, в состав которой входят многочисленные полимеры с разнообразными деталями строения и неравномерной локализацией. Еще одна причина – упрощенное и несколько пренебрежительное отношение к клеточной стенке как к мертвому внеклеточному образованию, существовавшее на протяжении веков. Многим казалось, что клеточная стенка – своеобразный «ящик», в котором находится живая протоплазма. «Неживые части растительных тканей» – формулировка из определения апопласта, в понятие которого входила совокупность клеточных стенок растения.
Клеточная стенка «ожила» в результате исследований последних десятилетий. Из «мертвого деревянного ящика», в котором живет протоплазма, она превратилась в многофункциональный компартмент растительной клетки с исключительно сложными механизмами формирования и функционирования. Растительной клеточной стенке присущи дуалистичекие свойства. С одной стороны, она отделяет одну клетку от другой, создавая условия для компартментации растительного организма, а с другой стороны, именно клеточная стенка объединяет клетки в целый организм, организуя их взаиморасположение и обеспечивая апопластный путь транспорта. Клеточная стенка должна обладать прочностью, чтобы выносить большие механические нагрузки, и в то же время быть способной растягиваться, во всяком случае, на некоторых этапах развития клетки. Клеточная стенка участвует в определении направления и скорости растяжения клетки, в реакции на стресс, в формировании водо- и ион-связывающей способности ткани, в механизмах узнавания клеток, в частности клеток патогенов, в обеспечении прорастания семян, созревания фруктов, опадения листьев, наконец – в образовании регуляторных молекул олигосахаринов. В последние годы достигнуты колоссальные успехи в расшифровке молекулярной структуры компонентов клеточных стенок, прежде всего полисахаридов, которые относятся к числу наиболее сложных молекул биологического происхождения. Необозримая информационная емкость клеточной стенки не нашла пока своей расшифровки, но уже сейчас очевидно, что эта субклеточная структура – значительно более динамичное образование, чем было общепризнанно.
Целлюлоза является веществом, широко распространенным в растительном мире. Вопрос об условиях и механизме биохимического синтеза целлюлозы – один из наиболее сложных и интересных в химии целлюлозы.
Роль целлюлозы, основного компонента клеточной стенки высших растений, играющего роль механического каркаса, непосредственно связана с особенностями химического строения макромолекулы и характером надмолекулярной структуры. Как образование макромолекул целлюлозы, так и формирование надмолекулярной структуры происходит в процессе биохимического синтеза, поэтому проблема исследования образования целлюлозы в природе имеет два аспекта – собственно биохимический, включающий вопрос о характере исходных реагирующих соединений, кинетике и механизме синтеза макромолекул, и структурно-химический механизм образования элементов надмолекулярной структуры и формирования сложной структуры полисахарида как полимера.
Возможность рационального использования целлюлозы различных отраслях народного хозяйства для получения материалов обладающих требуемыми свойствами, непосредственно зависит от выяснения основных вопросов строения целлюлозы и от подробного изучения свойств целлюлозных материалов. Это относится в первую очередь к тем отраслям промышленности, которые основаны на химической переработке целлюлозы (приготовление лаков, пленок, пластических масс, искусственного волокна, бездымного пороха и т.д.), а также к текстильной и бумажной промышленности. Разные представления о строении целлюлозы приводят к разным выводам об оптимальных условиях проведения процессов ее химической переработки и о методах, которые нужно применять для изменения в желаемом направлении физико-химических и механических свойств получаемых продуктов. Поэтому, естественно, вопросам строения целлюлозы посвящались и посвящаются многочисленные исследования.
В настоящее времени отсутствуют эффективные методы нативного выделения прочно связанных полисахаридов из клеточной стенки, что очень сильно препятствует изучению структуры клеточной стенки.
Проведённые в нашей лаборатории исследования позволили разработать метод выделения прочно связанных с клеточной стенкой матриксных полисахаридов в нативной форме, что позволило проводить более результативные исследования.
Целью данной работы является выделение и моносахаридный анализ прочно связанных полисахаридов с целлюлозой из клеточной стенки ксилемы льна.
Задачи
1) Отчистить клеточную стенку от пектиновых и гемицеллюлозных веществ.
2) Получить раствор прочносвязанных с целлюлозой полисахаридов.
3) Разделить фракций по молекулярным массам.
4) Провести моносахаридный анализ.
1. Обзор литературы
1.1 Растительная клеточная стенка
Растительная клеточная стенка была первой субклеточной структурой, увиденной учеными. Именно ее обнаружил в 1665 г. Роберт Гук при наблюдении в микроскоп среза пробкового дерева (см. Полевой, 1989, с. 13). И название «клетка» относилось первоначально именно к этой структуре из-за сходства строения исследуемых под микроскопом частей растений с ячейками («cells») пчелиных сот. Протоплазма была открыта позднее и терминологическая путаница при употреблении слова «клетка» долгое время смущала ученых (Кернер, 1906 с. 26).
В большинстве многоклеточных организмов, клетки погружены в специальный матрикс, который обеспечивает целостность клеток, и оказывает влияние на их форму, развитие и полярность. Но только в растительных клетках он, во-первых, настолько структурирован, что получил название стенки, а во-вторых, отличается огромным разнообразием компонентов, у которых детерминированы разнообразные детали строения и локализации. Ключевые различия между клетками растений и животных в отношении роста, формообразования, питания, развития, репродукции и защитных механизмов связаны именно с наличием клеточной стенки.
Размеры клеточной стенки
Клеточная стенка – слой структурированного материала, расположенного снаружи от плазмалеммы. Ее толщина обычно колеблется в пределах 0,1–10 мкм (Fry, 1988, с. 2).
Клеточная стенка составляет весьма значительную часть растительного организма: ее содержание в травянистых растениях составляет от 30 до 70% сухой массы, а в древесных растениях достигает 80–90%. Основные компоненты растительной клеточной стенки – самые распространенные органические соединения на Земле.
Объем клеточной стенки резко отличается в различных тканях. Однако даже в растущих клетках с тонкой клеточной стенкой ее объем может составлять до 10% объема клетки, а в сформированных тканях – до 90%.
Клеточная стенка – мощный потребитель фотосинтетических ассимилятов. Именно компоненты клеточной стенки, в первую очередь целлюлозу и лигнин, которые в растительном организме практически никогда не расщепляются, можно рассматривать как конечные продукты фотосинтеза. Затраты клетки на создания клеточной стенки необходимы для образования субстратов синтеза ее компонентов, их транспортировки по растению и внутри клетки, формирования сложного ферментативного комплекса, энергетических расходов.
1.2 Состав клеточных стенок
Основным компонентом клеточных стенок, значение которого часто недооценивается, является вода. Матрикс клеточной стенки на 75% состоит из воды и напоминает плотный водный гель (Cosgrove, 1997). Важной характеристикой клеточной стенки является ее рН. Различные методы определения обычно дают значения рН между 4 и 5 (Fry, 1988; Peters, Felle, 1991; Rayle, Cleland, 1992; Schopfer, 1993). Однако уникальность свойств клеточной стенки определяется присутствующими в ней полимерами, среди которых полисахариды, фенольные соединения (лигнин), белки.
Номенклатура полисахаридов построена так, что последним в слове называются формирующие остов молекул моносахара, к корню которых добавляется суффикс – ан. Например, «ксилан», если остов из ксилозы, «глюкан», если остов из глюкозы и т.д. Перед названием остова ставятся названия тех моносахаров, которые встроены в боковые цепочки. Например, «глюкуроноксилан», если к остову из ксилозы присоединены боковые звенья из глюкуроновой кислоты. Правда, структура полисахаридов значительно разнообразнее, чем можно изобразить словесными построениями; с примерами этого мы столкнемся, разбирая ниже структуру отдельных полисахаридов.
Полисахариды клеточной стенки подразделяют на три класса: 1) целлюлоза; 2) связующие гликаны (раньше называли гемицеллюлозы); 3) пектиновые вещества. Эта терминология в настоящее время наиболее распространена, хотя и достаточно условна, поскольку названия основаны на разных принципах, а граница между вторым и третьим классом несколько размыта. К первому типу относится, единственный полисахарид – целлюлоза. Название второго класса полисахаридов клеточной стенки определяется тем, что они связывают между собой микрофибриллы целлюлозы. Этот термин применяют ко всем нецеллюлозным полисахаридам, не относящимся к пектиновым веществам, поэтому в этот класс попадают и те полимеры, функции которых не совсем ясны, например глюко(галакто) маннаны. Раньше все эти полисахариды называли гемицеллюлозы, что означает «полуцеллюлозы» и совсем не отражает их состав и функции. Разграничение этих полимеров с пектиновыми веществами по наличию или отсутствию уроновых кислот также не совсем абсолютно, поскольку некоторые из связующих гликанов содержат уроновые кислоты, хотя и в минорных количествах. Путаница с подобной терминологией полисахаридов клеточной стенки приводит к тому, что исследователи пытаются использовать другой тип классификации – по экстрагируемости полисахаридов различными растворами. Например, «фракция, растворимая в оксалате аммония», или «фракция, растворимая, в уксусно-азотном реактиве». Эта классификация вполне корректна, но, кроме неблагозвучности, обладает тем недостатком, что не дает представления о том, какие именно полисахариды в нее входят, поскольку у различных растительных образцов одним и тем же растворителем могут извлекаться разные полимеры. С другой стороны, один и тот же полимер может попадать в различные фракции из-за наличия или отсутствия у его молекул связей с другими компонентами клеточной стенки. Предпочтительным, конечно, является перечисление индивидуальных полисахаридов, а не несколько безликой группы полимеров, но такая характеристика является достаточно трудоемкой и выполнена лишь для ограниченного круга объектов. И все же отнесение полисахарида к той или иной группе дает его первичную характеристику и ориентиры в возможных функциях.
Целлюлоза
Целлюлоза, вероятно, – самый известный полимер клеточной стенки. Подсчитано, что ежегодно на Земле синтезируется 180 миллиардов тонн целлюлозы (Delmer, 1999). Кроме высших растений, целлюлозу способны синтезировать большинство водорослей, некоторые слизевики (Dictyostelium
)
, ряд видов бактерий (включая цианобактерии, а также Acetobacter
xylinum
,
Agrobacteriem
tumifaciens
), некоторые грибы, а среди животных организмов – оболочники (Tunicata
)
(Saxena, Brown, 2005).
Строение молекул целлюлозы с первого взгляда производит впечатление очень простого, поскольку они представляют собой линейные молекулы гомополимера, состоящего из глюкозы. Все остатки глюкозы находятся в пиранозной форме и соединены между собой первым атомом углерода одной молекулы глюкозы и четвертым атомои углерода следующей молекулы. Тип связи одинаков для всех мономеров – b. Все вместе это записывается как b – (1®4) – D-глюкан. Степень полимеризации молекул целлюлозы составляет порядка 10 000, колеблясь в пределах 300 – 15 000.
Цепочки целлюлозы образуют кристаллическую надмолекулярную структуру: несколько дюжин молекул целлюлозы связаны между собой водородными связями (рис. 11) и силами Ван-дер-Вальса в микрофибриллу, толщина которой составляет 5–10 нм. Отдельная молекула целлюлозы обычно имеет степень полимеризации в несколько тысяч молекул глюкозы и достигает длины 2–3 мкм. Внутри микрофибриллы молекулы целлюлозы начинаются и кончаются в разных местах, поэтому микрофибрилла может достигать сотен микрометров в длину и содержать тысячи индивидуальных цепочек. В качестве аналогии можно привести льняную нитку, которую можно сделать неограниченной длины, хотя состоит она из волокон, каждое их которых – не более нескольких сантиметров в длину. Прочность на разрыв (tensilestrength) микрофибрилл целлюлозы выше, чем у стали.
В молекуле целлюлозы глюкозные остатки расположены в одной плоскости, так что формируется плоская, лентообразная цепочка. Внутри микрофибриллы отдельные молекулы расположены параллельно друг другу, то есть все восстанавливающие концы направлены в одну сторону (целлюлоза I). Подобную конфигурацию непросто объяснить с термодинамической точки зрения, поскольку при антипараллельной ориентации появляется возможность для дополнительной водородной связи, что делает такое расположение более вероятным. При самосборке целлюлозные цепи соединяются именно в антипараллельной ориентации (целлюлоза II).
В микрофибрилле высокоупорядоченные, кристаллические участки чередуются с относительно неупорядоченными, аморфными, которые отличаются по своим свойствам. Степень кристалличности (доля упорядоченных участков) в растительных объектах довольно высока и составляет от 60 до 80%. Аморфные участки – «слабое место» микрофибрилл. Различают также Ia
(triclinicunit) и Ib
(monoclinicunit) структуры целлюлозы I, которые отличаются молекулярной конформацией и упаковкой кристалла (Delmer, 1999). Доля Ia
-типа колеблется от 64% у Valonia
до 20% в волосках семян хлопчатника (Gossipium
hirsutum
) (Brett, 2000).
Термин «целлюлоза» (цит. по Franz, Blashek, 1990) был введен в 1838 году А. Payen и сначала использовался для обозначения целого ряда полисахаридов, поскольку не существовало способов их разделения. Аналогом этого термина в русском языке является «клетчатка». Schulze в 1891 году предложил использовать название «целлюлоза» для полисахаридов, устойчивых к разбавленной кислоте и щелочи и дающих в результате гидролиза глюкозу. Несколько позднее Nageli (с использованием поляризованного света) установил кристалличность целлюлозы, которая окончательно была доказана после появления рентгеновского анализа. Химическая природа целлюлозы как b-D – (1®4) – глюкана была установлена в 1932 году В. Хеуорсом (W.N. Haworth). Микрофибриллы целлюлозы, будучи кристаллическим образованием, расположенным снаружи от плазмалеммы, были одной из первых структур, надежно идентифицированных с помощью электронного микроскопа (Frey-Wisslingetal., 1948; Prestonetal., 1948).
Важно помнить, что микрофибриллы не ветвятся, они представляют из себя только «голые палки». Из них одних не удалось бы сформировать сколь-нибудь прочную объемную структуру. Сцепляют их между собой молекулы так называемых связующих гликанов.
Связующие гликаны
Остов молекул связующих гликанов устроен по тому же принципу, что и молекулы целлюлозы. Но к этому остову с определенной периодичностью добавлены боковые ответвления, которые нарушают линейность молекулы полисахарида. Другим способом получения изгибов молекулы является чередование типов связи мономеров внутри нее. В результате получается изогнутая во многих местах молекула, содержащая линейные участки. Этими линейными участками молекулы связующих гликанов взаимодействуют как с молекулами целлюлозы, так и между собой. В результате формируется прочная сеть, состоящая из двух типов молекул: целлюлозы и связующих гликанов. В роли связующих гликанов у разных организмов выступают различные полисахариды.
Ксилоглюкан. У двудольных основным связующим гликаном является ксилоглюкан
. Судя по названию, ксилоглюкан – молекула, остов которой сформирован из глюкозы, а боковые цепочки – из ксилозы. Однако название очень упрощено, в нем поименованы лишь основные мономеры. На самом деле в состав боковых цепочек ксилоглюкана входят еще галактоза и фукоза, причем у разных организмов могут быть свои нюансы в составе полимера. В целом же он устроен именно по тому принципу, о котором говорилось выше. Остов молекулы построен точно так же, как молекула целлюлозы, а наличие боковых цепочек приводит к изгибам полисахарида.
Остановимся подробнее на структуре полимера, охарактеризовав на его примере способы графической характеристики полисахаридов. Остов молекулы представляет из себя b-D – (1®4) – глюкан, который ветвится путем присоединения к С-6 атомам глюкозилов остатков ксилозы. В свою очередь, остатки ксилозы могут быть соединены с остатками галактозы или арабинозы, а галактоза – с фукозой.
Расположение боковых цепочек ксилоглюкана является регулярным. Обычно в ксилоглюкане можно выделить повторяющиеся блоки, составленные из 6–11 моносахаридов, пропорции которых варьируют у различных организмов и в различных тканях. У однодольных растений, клеточная стенка которых устроена несколько иначе, чем у двудольных, функции связующих гликанов выполняют глюкан со смешанным типом связи и глюкуроноарабиноксилан.
Глюкан со смешанным типом связи.Глюкан со смешанным типом связей – линейная молекула, в которой чередуются участки, где остатки глюкозы связаны b – (1®4) – связью, как в молекуле целлюлозы, и участки, где они связаны b – (1®3) – связью. Это приводит к изгибам молекулы и делает невозможным формирования водородных связей с другими полимерами по всей длине полисахарида. b – (1®3) – связь обычно встречается через два или три остатка глюкозы, связанных b – (1®4) – связью; примерно 10% (1®4) – олигоглюкозидов могут быть более длинными.
Ксилан. Глюкуроноарабиноксилан. Кс
иланы относятся к очень распространенным соединениям, часто являясь ключевыми полисахаридами матрикса вторичных клеточных стенок (например, в древесине). Остов этих молекул состоит из ксилозы – моносахара, конформационно очень схожего с глюкозой, – b-D – (1®4) – ксилан. В боковых цепочках (у С-2 или С-3) присутствует арабиноза и глюкуроновая кислота. Глюкуроновая кислота, присоединенная к ксилану, часто бывает метилированной. Доля ксилозных остатков, к которым присоединены боковые цепочки, состоящие из арабинозы и / или глюкуроновой кислоты, варьирует от 10% до 85%. Чтобы подчеркнуть высокую степень замещенности, ксилан называют глюкуроноарабиноксиланом.
Маннан. Глюкоманнан. Галактоманнан. Галактоглюкоманнан.Неразветвленный маннан – редко встречающийся полисахарид, состоящий из остатков D-маннозы, соединенных b-D – (1®4) связями. Этот полимер способен образовывать агрегаты, напоминающие микрофибриллы целлюлозы.
Глюкоманнаны являются полисахаридами, у которых остов состоит из чередующихся остатков маннозы и глюкозы. Таким образом, его название является иллюстрацией того, что для полисахаридов сложно выработать общие правила наименования (исходя из них, можно было бы предположить, что глюкоза формирует боковые цепочки). b-D – (1®4) – маннозные звенья прерываются b-D – (1®4) – глюкозными участками, доля которых составляет 25–35%.
С другой стороны, существуют сходные полимеры, в остове которых глюкоза отсутствует. В этом случае, название «галактоманнан» сформировано в полном соответствии с правилами номенклатуры полисахаридов, поскольку остов построен из маннозы, а боковые цепочки – из галактозы. Галактоманнаны – известные полисахариды клеточной стенки, присутствующие в семенах некоторых бобовых, где они играют роль запасных веществ.
Наконец, существуют галактоглюкоманнаны, остов которых устроен как глюкоманнан. При этом к некоторым остаткам маннозы присоединены боковые цепочки из одиночной a – (1®6) – галактозы (у С-6). Подобные полисахариды особенно распространенные в клеточной стенке голосеменных.
b – (1®3) – Глюкан (каллоза).Каллоза по химическому составу напоминает целлюлозу и является линейным гомополимером, состоящим из остатков глюкозы. Однако в образовани связи участвует не С-4, а С-3 атом, что приводит к совершенно иным свойствам молекулы. При классификации каллозу обычно объединяют с поперечно-связывающими гликанами, имея в виду, что ее молекула не содержит уроновых кислот, а значит, безусловно, не является пектиновым веществом. Однако правильнее, пожалуй, было бы выделить ее в особый класс полисахаридов клеточной стенки, поскольку она не участвует в образовани поперечных связей между микрофибриллами. Каллоза – своего рода «пожарный» полисахарид. Она образуется при поранении, при формировании срединной пластинки в ходе деления клетки; по-прежнему, обсуждается ее роль как промежуточного или запасного соединения.
Пектиновые вещества
Вся сеть микрофибрилл и связывающих их гликанов погружена в матрикс из пектиновых веществ. Слово «пектин» многим, наверное, знакомо в связи с мармеладом. Одна из статей в области изучения этих соединений так и называлась «Пектины – это не просто желе» (Jarvis, 1984). Пектиновые вещества – одни из самых сложных молекул биологического происхождения, которым приписывают разнообразные функции. Они были открыты в 1790 году, когда французский химик N. Vauquelin выделил из яблок вещество, образующее в водном растворе студень. Термин «пектин» (от греческого pektóV – свернувшийся) был введен в 1825 году H. Braconnot (Комиссаров, Спиридонов, 1998). Исторически сложилось деление пектиновых веществ на растворимые в воде (гидропектин) и нерастворимые – протопектин.
Непременным компонентом пектиновых веществ является галактуроновая кислота, следовательно, эти молекулы имеют заряд. Карбоксильные группы уроновых кислот могут экранироваться различными способами, например, ионами кальция или метиловыми группами, поэтому пектиновые вещества, являясь природными ионообменниками, могут создавать подвижную систему локального изменения рН, заряда и т.д. Это приводит к регулированию действия ферментов, локализованных в клеточной стенке. Кроме того, пектиновые вещества определяют пористость клеточной стенки. Обработкой клеточной стенки пектиназами (но не целлюлазами или протеиназами) можно вызвать увеличение размеров пор. Свободные карбоксильные группы уроновых кислот могут связывать Ca2+
и образовывать таким образом сшивки между молекулами пектиновых веществ. Считается, что пектиновые вещества являются гидрофильным наполнителем простанства между микрофибриллами целлюлозы и предотврашают их коллапс при падении тургорного давления (Jarvis, 1992).
В случае пектиновых веществ до сих пор неясно, что можно считать индивидуальным полимером. Например, существует мнение, что различные типы пектиновых веществ ковалентно связаны друг с другом (Mohnen, 1999; Vinckenetal., 2003). Тем не менее, на сегодняшний день общепринята классификация их как индивидуальных компонентов клеточных стенок.
Полигалактуроновая кислота.Основным компонентом пектиновых веществ является полигалактуроновая кислота – гомополимер из остатков галактуроновой кислоты, находящихся в пиранозной форме и связанных a – (1®4) – связью. Во многом благодаря наличию полигалактуроновой кислоты, клеточная стенка имеет отрицательный заряд и ведет себя как нерастворимый полианион (Nari et al., 1991). Степень полимеризации варьирует от 30 до 200. Соли полигалактуроновой кислоты называют пектатами.
Рамногалактуронан I. Рамногалактуронан I – гетерополимер, остов которого состоит из рамнозы и галактуроновой кислоты. Соотношение Рам: ГалК часто составляет 1:1 и тогда остов состоит из повторяющегося дисахарида (®4)-a-D-ГалКр
– (1®2)-a-L-Рамр
(1®). В других случаях соотношение может составлять 1:2 и более. Звенья рамнозы никогда не располагаются друг за другом, всгда перемежаясь остатками галактуроновой кислоты. К 20–80% остатков рамнозы могут быть присоединены различной длины (от 1 до 50 гликозильных остатков) цепочки арабиногалактанов, арабинанов и галактанов. Ветвление полисахарида происходит только в звене рамнозы (у С-4). Типы боковых цепочек рамногалактуронана в значительной степени определяют пористость клеточной стенки, ее заряд, рН и баланс ионов. Степень полимеризации рамногалактуронана I может сильно варьировать, обычно приводятся значения около 1000.
Рамногалактуронан II. Исходя из названия полисахарида, можно было бы предположить, что остов молекулы построен как в рамногалактуронане I – из чередующихся остатков рамнозы и галактуроновой кислоты, однако он устроен совсем по-другому. По разнообразию мономеров и типов связей между ними рамногалактуронан II – самый сложный их известных в живых организмах полисахаридов. Хотя степень полимеризации его всего около 60, общее число типов связей в этом небольшом полимере – 20.
Рамногалактуронан II содержит 12 различных моносахаридов, а именно галактуроновую кислоту (включая необычную, b-связанную, форму), рамнозу, арабинозу (в том числе, в необычной, пиранозной, форме), галактозу, фукозу, метилфукозу, метилксилозу, апиозу, глюкуроновую кислоту, редкую для клеточной стенки кетозу – кетодезоксиманнооктоновую кислоту, а также присутствующую только в рамногалактуронане II ацеровую кислоту (3-С
-карбокси-5-дезокси-L-ксилоза) и глюкозу. Остов молекулы содержит всего 9 a – (1®4) – D-ГалКр
звеньев, к С-2 и / или С-3 атомам которых присоединены боковые цепочки сложной структуры. Состав этих цепочек охарактеризован достаточно тщательно, однако порядок их расположения на остове молекулы неизвестен и указывается на схемах предположительно.
Содержание рамногалактуронана II в клеточных стенках составляет всего 0,2–3,6% от сухой массы клеточных стенок (Matsunagaetal. 2004), однако, учитывая его сравнительно небольшую молекулярную массу, число молекул этого полимера вполне сопоставимо с числом молекул самых распростаненных полисахаридов. Структура рамногалактуронана II исключительно консервативна и, несмотря на ее сложность, практически не отличается у разных организмов и в различных тканях.
Ксилогалактуронан. Апиогалактуронан.Значительно менее распространенными, чем обсуждавшиеся ранее пектиновые вещества, являются ксилогалактуронан и апиогалактуронан, которые описаны сравнительно недавно, встречаются лишь в некоторых растительных объектах и не считаются типичными. Эти полимеры родственны полигалактуроновой кислоте, которая составляет остов молекулы. Боковые цепочки состоят из одиночных остатков ксилозы или апиозы, присоединенных к С-3 галактуроновой кислоты. Частота ветвления варьирует от 25 до 75% (Scholsetal., 1995).
Арабинан, галактан. В растительных клеточных стенках присутствуют цепочки a-L – (1®5) – арабинана и b-D – (1®4) – галактана. И те, и другие могут быть линейными или разветвленными, однако боковые цепочки, если присутствуют, представлены, как правило, одиночными остатками того же сахара, что и остов. Дискуссионным остается вопрос, всегда ли они являются боковыми ответвлениями рамногалактуронана I, или могут быть «самостоятельными» соединениями. Это же справедливо в отношении арабиногалактана.
Арабиногалактан. В растительных клеточных стеках обнаруживают арабиногалактан двух типов. В первом из них галактозные звенья соединены по С-4, во втором – по С-3 или С-6. Арабиногалактан II типа входит также в состав протеогликанов клеточной стенки. Антитела на арабиногалактан II типа реагируют и с арабиногалактановыми белками, и с рамногалактуронаном I, свидетельствуя, что в состав этих сложных молекул могут входить одинаковые арабиногалактановые звенья.
Прочно связанные полисахариды
Типы первичной клеточной стенки.
Типы вторичной клеточной стенки
Клеточная стенка ксиланового типа
Клеточная стенка желатинозного типа
Характеристика клеточных стенок исследуемых объектов
Клеточная стенка клеток ксилемы льна
Клеточная стенка волокон кокоса
Клеточная стенка волокон банана
Экспериментальная часть
2. Материалы и методы
2.1 Растительный материал
Лен (Linum
usitatissimum
) – лубоволокнистые культуры в которых формирование клеточных стенок проходит исключительно интенсивно, а стадии развития детально охарактеризованы (Горшкова и др., 2003;). Ксилема для анализа была взята ниже точки слома 5 см, растения собирали через месяц после посадки. Лен был выращен на открытом грунте. Ксилему от флоэмы отчищали вручную, старались максимально отчистить от флоэмы, поэтому этот процесс проводился дважды, а те участки, которые плохо поддавались отчистки просто отбрасывались.
Волокна банана.
Волокна кокоса.
2.2 Подготовка ксилемы льна к экспериментальной части
1) Фиксировали замороженную ксилему при 1000
С тридцать минут.
2) Сушили 60 минут при 600
С.
3) Измельчали ксилему на «мельнице».
Отчистка от непрочносвязынных пектиновых и гемицеллюлозных веществ
1) 2 гр. ксилемы заливаем 100 гр. 1% оксалата аммония.
2) Эту колбу с смесью кипятим на водяной бане 60 минут при 970
С, при этом колба закрыта.
3) Охлаждаем колбу.
4) Эту смесь центрифугируем 10 мин на 4000 об/мин и сливаем надосадочную жидкость.
5) Еще раз заливаем 1% оксалатом аммония нашу смесь и кипятим на водяной бане 30 мин., также центрифугируем.
6) Вымываем оксалат аммония водой: заливаем дисцилированную воду в центрифужные стаканчики тщательно перемешиваем и центрифугируем на 4000 об/мин.
7) Так еще три раза. Проверяем рН универсальной лакмусовой бумажкой.
8) Готовим раствор 4Н КОН и 3% борной кислоты: 44,888 гр. КОН и 6 гр. Н3
ВО3
на 200 мл. дисцилированной воды.
9) Заливаем 100 мл раствора к ксилеме и отстаиваем 1 час, затем центрифугируем и выливаем надосадочную жидкость.
10) Промываем водой: заливаем дисцилированную воду в центрифужные стаканчики тщательно перемешиваем и центрифугируем на 4000 об/мин. промываем до тех пор пока рН среды не будет нейтральным.
2.3 Растворение целлюлозы
Получение раствора хлорида лития в
N
,
N
– диметилацетамиде
1) N, N-диметилацетамид (ДМА) сушили над молекулярными ситами 4А.
2) В колбу присыпали 8 гр. хлорида лития и нагревали в термостате до 1800
в течение 4–5 часов, затем охлаждали и приливали 100 мл. высушенного над молекулярным ситом ДМА.
3) Колбу закрывали стеклянной пробкой и раствор перемешивали на шейкере до растворения хлорида лития (оставляли на ночь).
Выделение прочно связанных полисахаридов из растворенной целлюлозы клеточной стенки ксилемы льна
1) 1 гр. «целлюлозы» суспензировали в 10 мл. воды 40–60 мин.
2) Отделяли на воронке Шота со вставленным нейлоновым фильтром.
3) Осадок промывали на фильтре ацетоном.
4) Выдерживали в ацетоне (10 мл., 60 мин.)
5) Осадок промывали на фильтре ДМА
6) Выдерживали в ДМА (10 мл., 40–60 мин.)
7) Оставляли на ночь в свежей порции ДМА
8) На воронке шота отделяли «целлюлозу»
9) Суспензировали ее в 100 мл раствора хлорида лития в N, N – диметилацетамиде.
10) Колбу закрывали стеклянной пробкой и перемешивали до растворения целлюлозы на шейкере (1–2 дня).
11) Раствор целлюлозы по каплям добавляли к 100 мл. дисцилированной воды при перемешивание на магнитной мешалке. Раствор оставляли на ночь.
12) Отделяли выпавшую целлюлозу на воронке Шота с нейлоновым фильтром и промывали ее два – три раза дисцилированной водой (по 50 мл.) Получили Фильтрат 1
13) Фильтрат 1 Заливали в диалезный мешок 12 – 14 тысяч Дальтон.
14) Диалезный мешок клали в 3-х литровую банк с дисцилированной водой и ставили на магнитную мешалку, воду в банке меняли 4 раза.
15) На роторном испарителе концентрировали раствор.
16) Хроматографируем концентрированный раствор. Готовим 40 пробирок, моем их хромкой и сушим в сушильном шкафу при 1200
С.
17) На хроматографе задаем программу (по каплям, 40 пробирок, в одну пробирку 40 капель ≈ 1,8 мл.)
18) В эпиндорф вливаем концентрированный раствор и центрифугируем 5 мин. при 6 000 об/мин.
19) Хроматографирование на Sepharose CL-4B элюент 0,02% NaN3
и АсNa 10 mMpH 5,5, разделение по молекулярным массам.
20) Определяем количество сахаров методом Дюбуа (фенольным методом). 200 мкл образца+200 мкл 5% раствора фенола, помещаем пробирки в холодную воду и добавляем 1 мл. концентрированной серной кислоты. Кипятим на водяной бане 10 минут и охлаждаем. Измеряем оптическую плотность при 490 нм. (Смотри гафик1)
21)Выпавшую целлюлозу суспензировали в 50 мл 0,01 М растворе NaAc, рН 4,5 + 0,05% NaN3
, содержавший 0,1 мл раствора целлюлазы и инкубировали в течение ночи.
22) Раствор фильтровали на воронке Шота и осадок заново суспезировали в новой порции раствора фермента. Мы получили фильтрат 2.
23) Фильтрат 2 обессоливали на колонке Sephadex G-25, элюент 0,02% NaN3
. 0,005 гр. BlueDextran 2000 кД и 0,250 гр. СоCl2
растворяли в 5 мл дисцилированной воды и хроматографируем. BlueDextran 2000 кД начало 25 мл конец 30 мл. СоCl2
начало 60 мл. конец 73 мл.
24) Хроматографирование на Sepharose CL-4B элюент 0,02% NaN3
и АсNa 10 mMpH 5,5, разделение по молекулярным массам.
25) Определяем количество сахаров методом Дюбуа (фенольным методом). 200 мкл образца+200 мкл 5% раствора фенола, помещаем пробирки в холодную воду и добавляем 1 мл. концентрированной серной кислоты. Кипятим на водяной бане 10 минут и охлаждаем. Измеряем оптическую плотность при 490 нм (Смотри график2).
2.4 Моносахаридный анализ
1) Фильтрат 1 и Фильтрат 2 обессоливаем на колонке SepharoseG250, элюент 10мМ пиридин и 10мМ уксусная кислота рН 4,5.
2) Высушиваем при 600
С до полного отсутствия воды.
3) Добавляем 400 мл ТФУ (Трифторуксусная кислота), помещаем в сушильный шкаф на 1 час при температуре 1200
С.
4) Затем опять сушим при 600
С
5) Растворяем водой и определяем моносахаридный соcтав на Daionex(Данные моносахаридного состава смотри в таблице 1).
6) Мы объединили наши пробирки в следующие фракции до целлюлазной обработки: фракция 1 5–10; фракция 2 11–16; фракция 3 17–23; фракция 4 24–26; фракция 5 27–35.
Фракции после целлюлазной обработки: фракция 1 6–10; фракция 2 11–16; фракция 3 17–19; фракция 4 20–24; фракция 5 25–26; фракция 6 27–31.
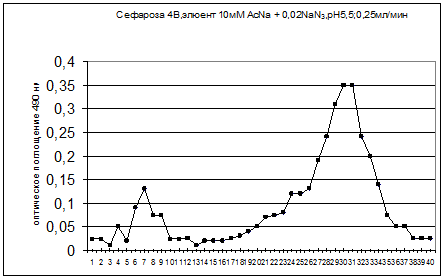
Рисунок 1 – Распределение прочносвязанных полисахаридов во фракциях после растворения целлюлозы до ферментативной обработки.

Рисунок 2 – Распределение прочносвязанных полисахаридов после растворения после ферментативной обработки
Таблица 1. Моносахаридный состав во фракциях
| Рамноза |
Арабиноза |
Галактоза |
Глюкоза |
Ксилоза |
Галактуроновая кислота |
Глюкуроновая кислота |
| Фракция 1, после целлюлазы |
49.290 |
33.680 |
68.173 |
7.624 |
5.855 |
254.246 |
1.166 |
| Фракция 2, после целлюлазы |
61.981 |
43.261 |
99.500 |
6.804 |
13.317 |
311.360 |
3.365 |
| Фракция 3, после целлюлазы |
25.478 |
17.145 |
79.425 |
6.605 |
31.317 |
161.033 |
10.089 |
| Фракция 4, после целлюлазы |
6.349 |
7.295 |
48.161 |
25.828 |
86.452 |
67.533 |
10.577 |
| Фракция 5, после целлюлазы |
4.020 |
14.716 |
40.078 |
40.401 |
210.336 |
3.171 |
5.546 |
| Фракция 6, после целлюлазы |
1.678 |
4.407 |
13.217 |
28.017 |
136.191 |
6.781 |
1.291 |
| Фракция 1, до целлюлазы |
0 |
0 |
0 |
0 |
0.535 |
358.507 |
0 |
| Фракция 2, до целлюлазы |
0.556 |
0.481 |
2.689 |
5.364 |
1.107 |
0.476 |
0 |
| Фракция 3, до целлюлазы |
12.596 |
15.664 |
39.576 |
12.444 |
0 |
21.317 |
1.393 |
| Фракция 4, до целлюлазы |
20.762 |
24.131 |
54.572 |
11.218 |
54.753 |
67.032 |
2.955 |
| Фракция 5, до целлюлазы |
82.195 |
151.536 |
325.976 |
131.493 |
0 |
426.007 |
19.982 |
3. Результаты и их обсуждение
При подготовке ксилемы в первую очередь надо было остановить все процессы происходящие в ней поэтому фиксировали ее. Обработка ксилемы включала два основных этапов. Первое – механическое растирание на мельнице для того что бы получит муку, так как так лучше будет удаляться не прочно связанные вещества с целлюлозой и лучше растворяться. Второе – экстракция пектина и гемицеллюлозы с хелатирующими и щёлочными растворами.
Удаление пектиновых веществ с мы осуществили с помощью оксалата аммония, как известно пектиновые вещества связаны между собой ионами Са, а оксалат аммония является хелатирующим веществом которое связывается с ионами Са и пектиновые вещества отделяются от целлюлозы. Для удаление оксалата аммония и пектиновых веществ из нашей колбы мы центрифугируем их 3 раза с водой.
Обработка клеточной стенки ксилемы льна 4% NaOHпозволяла извлекать гемицеллюлозные вещества, которые не извлекаются оксалатом аммония, нужен более сильный раствор. Мы применяли 4% NaOH который высвобождает целлюлозу от гемицеллюлозных веществ. Затем мы снова удаляли щелочь и гемицеллюлозные вещества с помощью метода центрифугирования при этом проверяли рН универсальной лакмусовой бумажкой, промывали до нейтрального.
Выдерживание в воде «отчищенной» ксилемы необходимо для так называемой активации, без этого этапа целлюлоза не растворяется. Выдерживание в ацетоне и ДМА необходимо для полного удаления воды, так как при наличие воды целлюлоза выподает в осадок. Клеточную стенку волокна растворяли в » 8% растворе LiCl в ДМА, и заново осаждали целлюлозу путём добавления воды. Осаждение в воде необходимо для того что бы фермент полностью расщепил целлюлозу, если опустить этот этап то целлюлоза плохо будет подвергаться гидролизу. 8% раствор LiCl в ДМА мы используем так как они вместе являются хорошими растворителями целлюлозы и не реагируют с ней, к тому от них легко избавиться с помощью хроматографии. Раствор отделяли от целлюлозы (Фильтрат 1). Целлюлозу суспензировали в содержащем целлюлазу ацетатном буфере. В течение 1–2 суток фермент растворял осаждённую целлюлозу (Фильтрат 2). Мы получили раствор прочно связанных полисахаридов с целлюлозой. Далее с помощью ТФУ мы разрушаем все гликозидные связи и при высушивание весь ТФУ испаряется, и остаются мономеры сахаров. Dionex эта ионно-обменная хроматография, на ней мы и определяем моносахаридный состав, заранее вкалываются стандарты моносахоров по которым мы и опознаем конкретный моносахар.
В дальнейшем планируется провести ЯМР анализ и определить тип связующих гликанов.
Выводы
1. Отчистили клеточную стенку от пектиновых и гемицеллюлозных веществ.
2. Получили раствор прочносвязанных с целлюлозой полисахаридов.
3. Разделили фракции по молекулярным массам.
4. Определили моносахаридный состав во фракциях.
Литература
1. Керне, А. Жизнь растений. / А. Керне // СПб.: Просвещение, – 1906. – Т. 2. – 838 с.
2. Полевой, В.В. Физиология растений. / В.В. Полевой // М.: Высшая школа, – 1989. – 464 с.
3. Brett, C.T. Cellulose microfibrils in plants: biosynthesis, deposition, and integration into the cell wall / C.T. Brett // Int. Rev. Cytol. –2000. –V. 199. –P. 61–99.
4. Chivasa, S., Proteomic analysis of the Arabidopsis thaliana cell wall / S. Chivasa, B.K. Ndimba, W.J. Simon, D. Robertson, X.L. Yu, J.P. Knox, P. Bolwell, A.R. Slabas // Electrophoresis. – 2002. – V. 23. – P. 1754–1765.
5. Cosgrove, D.J. Group I allergens of grass pollen as cell wall-loosening agents / D.J. Cosgrove, P.A. Bedinger, D.M. Durachko // Proc. Natl. Acad. Sci. USA. –1997. –V. 94. –P. 6559–6564.
6. Delmer, D. Cellulose biosynthesis: exciting times for a difficult field of study / D. Delmer // Plant Physiol. Plant Mol. Biol. –1999. –V. 50. –P. 245–276.
7. DeWitt, G. Comparative compositional analysis of walls with two different morphologies: archetypical versus transfer-cell-like / G. DeWitt, J. Richards, D. Mohnen, A.M. Jones / Protoplasma. – 1999. – V. 209. – №3/4. – P. 238–245.
8. Frey-Wissling, A. Submicroscopic morphology of protoplasm and its derivatives. / A. Frey-Wissling. // N.Y.: Elsevier. – 1948. – 47 p.
9. Fry, S.C. The growing plant cell wall: chemical and metabolic analysis. / S.C. Fry // London: Longman Sci. and Technic. – 1988. – 333 p.
10. Fry, S.C., Xyloglucan endotransglucosylase, a new wall-loosening enzyme activity from plants / S.C. Fry, R.C. Smith, K.F. Renwick, D.J. Martin, S.K. Hodge, K.J. Matthews // Biochem. J. –1992. –V. 282. –P. 821–828.
11. Gaspar, T., Peroxidases 1970–1980: A survey of their biochemical and physiologic roles in higher plants. / T. Gaspar, C. Penel, T. Thorpe, H. Greppin. // Switzerland: University of Geneva Press. – 1982. – P. 60–121.
12. Gaspar, Y., The complex structures of arabinogalactan-proteins and the journey towards understanding function / Y. Gaspar, K.L. Johnson, J.A. McKenna, A. Bacic, C.J. Schultz // Plant Mol. Biol. –2001. –V. 47. –P. 161–176.
13. Jarvis, M.C. Control of thickness of collenchyma cell walls by pectins / M.C. Jarvis // Planta. – 1992. – V. 187. – P. 218–220.
14. Jose-Estaniol, M., Plant cell wall glycoproteins and their genes / M. Jose-Estaniol, P. Puigdomenech // Plant Physiol. Biochem. – 2000. – V. 38. – №1/2. –P. 97–108.
15. Lamport, D.T.A. The protein component of primary cell walls / D.T.A. Lamport // Adv. Bot. Res. – 1965. – V. 2. – P. 151–218.
16. Lamport, D.T.A., The use of tissue cultures for the study of plant cell walls / D.T.A. Lamport, D.H. Northcote // Biochem. J. –1960. –V. 76. –P. 52.
17. Mohnen, D. Biosynthesis of pectins and galactomannans / D. Mohnen // Comprehensive Natural Products Chemistry / Eds Pinto B.M., Barton D.H.R., Meth-Cohn O. Oxford: Elsevier. – 1999. – P. 497–527.
18. Mohnen, D. Biosynthesis of pectins and galactomannans / D. Mohnen // Comprehensive Natural Products Chemistry / Eds Pinto B.M., Barton D.H.R., Meth-Cohn O. Oxford: Elsevier. – 1999. – P. 497–527.
19. Nishitani, K., Endo-xyloglucan transferase, a novel class of glycosyltransferase that catalyzes transfer of a segment of xyloglucan molecule to another xyloglucan molecule / K. Nishitani, R. Tominaga // J. of Biol. Chem. –1992. –V. 267. –P. 21058–21064.
20. Saxena, I.M., Cellulose biosynthesis: current views and evolving concepts / I.M. Saxena, R.M. Brown // Ann. Bot. – 2005. – V. 96. – №1. – P. 9–21.
21. Schroder, R., Mannan transglycosylase: a novel enzyme activity in cell walls of higher plants /R. Schroder,T.F. Wegrzyn,K.M. Bolitho,R.J. Redgwell // Planta. – 2004. – V. 219. – P. 590–600.
22. Schroder, R., LeMAN4 endo-beta-mannanase from ripe tomato fruit can act as a mannan transglycosylase or hydrolase / R. Schroder,T.F. Wegrzyn, N.N. Sharma,R.G. Atkinson // Planta. – 2006. – V. 224. – P. 1091–1102.
|