Министерство образования Республики Беларусь
УО МОГИЛЕВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ ПРОДОВОЛЬСТВИЯ
КАФЕДРА ХИМИЧЕСКОЙ ТЕХНОЛОГИИ ВЫСОКОМОЛЕКУЛЯРНЫХ СОЕДИНЕНИЙ
БИОЛОГИЧЕСКАЯ ХИМИЯ
КОНСПЕКТ ЛЕКЦИЙ
для студентов специальностей 49 01 01, 49 01 02, 91 01 01
БЕЛКИ И НУКЛЕИНОВЫЕ КИСЛОТЫ
Могилев, 2004
УКД
ББК
Рассмотрен и рекомендован к изданию кафедрой химической технологии высокомолекулярных соединений
Протокол № __ от «__»_____________2004 г.
Рассмотрен и рекомендован к изданию секцией выпускающих кафедр.
Протокол № __ от «__»_____________2004 г.
Составители: доцент Макасеева О.Н.
Рецензент: доцент Шуляк Т.Л.
Ó Могилевский государственный университет продовольствия
Содержание
1. БЕЛКИ
1.1Строение и общие свойства аминокислот.
1.2Классификация аминокислот.
1.3Кислотно-основные свойства аминокислот.
1.4Спектры поглощения аминокислот.
1.5Химические реакции аминокислот.
1.6Пептиды.
1.7Белки.
1.8Строение белковой молекулы.
1.9Физико-химические свойства белков.
1.10Выделение белков и установление их однородности.
1.11Классификация белков.
2. НУКЛЕИНОВЫЕ КИСЛОТЫ.
2.1Состав нуклеиновых кислот.
2.2Нуклеозиды.
2.3Нуклеотиды
2.4Первичная структура нуклеиновых кислот.
2.5Вторичная и третичная структуры ДНК.
2.6Структура РНК
Рекомендуемая литература
1. БЕЛКИ
Основной структурной единицей белков являются a-аминокислоты. В природе встречается примерно 300 аминокислот. В составе белков найдено 20 различных a-аминокислот (одна из них – пролин, является не амино-, а иминокислотой). Все другие аминокислоты существуют в свободном состоянии или в составе коротких пептидов, или комплексов с другими органическими веществами.
a-Аминокислоты представляют собой производные карбоновых кислот, у которых один водородный атом, у a-углеродного атома замещен на аминогруппу (–NН2
), например:
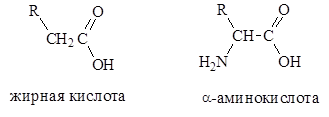
Различаются аминокислоты по строению и свойствам радикала R. Радикал может представлять остатки жирных кислот, ароматические кольца, гетероциклы. Благодаря этому каждая аминокислота наделена специфическими свойствами, определяющими химические, физические свойства и физиологические функции белков в организме.
Именно благодаря радикалам аминокислот, белки обладают рядом уникальных функций, не свойственных другим биополимером, и обладают химической индивидуальностью.
Значительно реже в живых организмах встречаются аминокислоты с b- или g-положением аминогруппы, например:
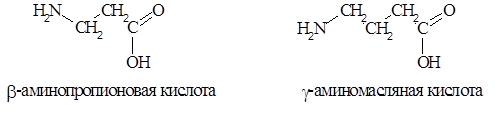
В молекулах всех природных аминокислот ( за исключением глицина) у a-углеродного атома все четыре валентные связи заняты различными заместителями, такой атом углерода является асимметрическим, и получил название хирального атома. Вследствие этого растворы аминокислот обладают оптической активностью – вращают плоскость плоскополяризованного света. Причем, при прохождении через них поляризованного луча происходит поворот плоскости поляризации либо в право (+), либо влево (–). По расположению атомов и атомных группировок в пространстве относительно асимметрического атома различают L- и D-стереоизомеры аминокислот. Знак и величина оптического вращения зависят от природы боковой цепи аминокислот (R-группы).
Число возможных стереоизомеров ровно 2n
, где n – число асимметрических атомов углерода. У глицина n = 0, у треонина n = 2. Все остальные 17 белковых аминокислот содержат по одному асимметрическому атому углерода, они могут существовать в виде двух оптических изомеров.
В качестве стандарта при определении L и D-конфигураций аминокислот используется конфигурация стереоизомеров глицеринового альдегида.
Расположение в проекционной формуле Фишера NH2
-группы слева соответствуют L-конфигурации, а справа – D-конфигурации.
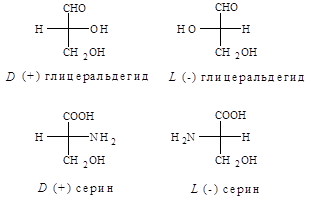
Следует отметить, что буквы L и D означают принадлежность того или иного вещества по своей стереохимической конфигурации к L или D ряду, независимо от направленности вращения.
В составе белков обнаруживаются только L-изомеры аминокислот. D-формы аминокислот в природе встречаются редко и обнаружены лишь в составе белков клеточной стенки (гликопротеинов) некоторых бактерий и в пептидных антибиотиках (грамицидин, актиномицин и т.д.). L-формы хорошо усваиваются растениями и животными и легко включаются в обменные процессы. D-формы не ассимилируются этими организмами, а иногда даже ингибируют процессы обмена. Это объясняется тем, что ферментативные системы организмов специфически приспособлены к L формам аминокислот.
L и D формы аминокислот оказывают различное физиологическое воздействие на организм человека – различаются по вкусу: D-изомеры сладкие, L-формы горькие или безвкусные.
Взаимопревращение D и L-энатиомеров называется рацемизацией. Превращение DÛL – это один из метаболических процессов в живых организмах, причем равновесие этого метаболического процесса сильно смещено в сторону образования L-формы. Когда метаболические процессы после смерти организма прекращаются, процесс DÛL продолжается самопроизвольно с очень малой скоростью, переводя для каждой аминокислоты к соотношению D/L-энантиомеров, характерному для неметаболического равновесия. Для достижения такого равновесия могут потребоваться десятки тысяч лет. Новый метод определения геологического возраста образца основан на измерении соотношения D/L-энантиомеров аспарагиновой кислоты в образцах окаменелых костей. Результаты, полученные методом D /L-датирования, хорошо дополняют другие данные, полученные, например, радиоуглеродным методом.
Кроме 20 стандартных аминокислот встречающихся почти во всех белках, существуют еще нестандартные аминокислоты, являющиеся компонентами лишь некоторых типов белков – эти аминокислоты называют еще модифицированными. Около 150 из них уже выделены. Эти аминокислоты образуются после завершения синтеза белка в рибосоме клеток путем посттрансляционной химической модификации.
Один из примеров особенно важной модификации – окисление двух-SН–групп цистеиновых остатков с образованием аминокислоты цистина, содержащей дисульфидную связь. Так же легко происходит и обратный переход.
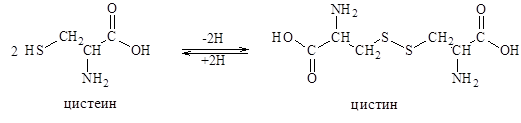
Таким путем образуется одна из важнейших окислительно-востановительных систем живых организмов. В больших количествах цистин содержится в белках злаковых – клейковине, в белках волос, рогов.
Другие примеры аминокислотной модификации-гидроксипролин и гидроксилизин, которые входят в состав коллагена-основного белка соединительной ткани животных.
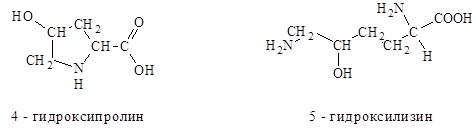
В состав белка протромбина (белок свертывания крови) входит g-карбоксиглутаминовая кислота, а в ферменте глутатионпероксидазе открыт селеноцистеин, в котором ( S ) сера заменена на ( Se ) селен.
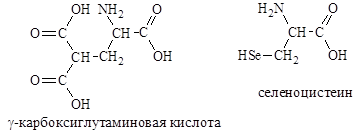
Существует несколько видов классификаций аминокислот входящих в состав белка.
В основу одной из классификаций положено химическое строение радикалов аминокислот. Различают аминокислоты:
1. Алифатические – глицин, аланин, валин, лейцин, изолейцин:
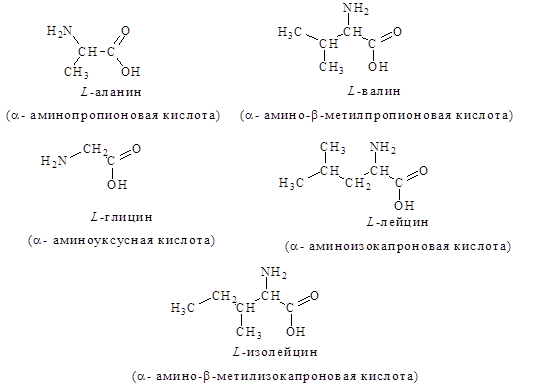
2. Гидроксилсодержащие – серин, треонин:
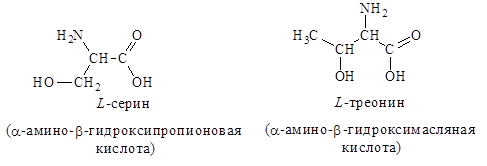
3. Серосодержащие – цистеин, метионин:
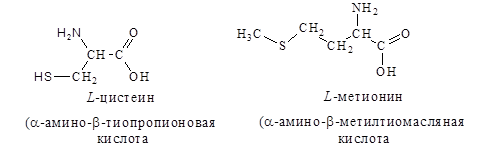
4. Ароматические – фенилаланин, тирозин, триптофан:
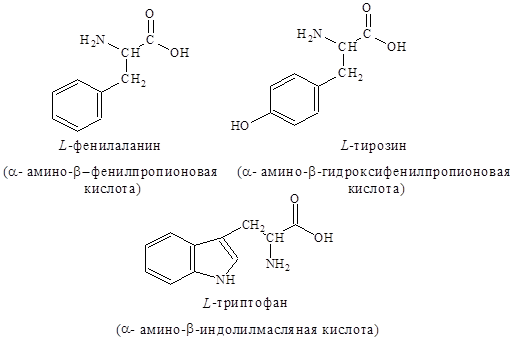
5. С анионобразующими группами в боковых цепях-аспарагиновая и глутаминовая кислоты:

6. и амиды-аспарагиновой и глутаминовой кислот – аспарагин, глутамин.
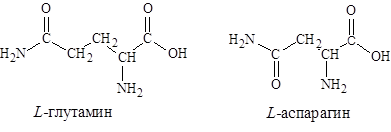
7. Основные – аргинин, гистидин, лизин.
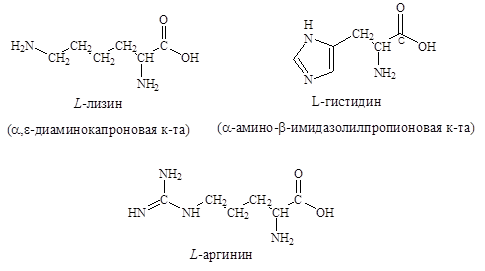
Второй вид классификации основан на полярности R-групп аминокислот. Различают полярные и неполярные аминокислоты. У неполярных в радикале есть неполярные связи С–С, С–Н, таких аминокислот восемь: аланин, валин, лейцин, изолейцин, метионин, фенилаланин, триптофан, пролин.
Все остальные аминокислоты относятся к полярным (в R-группе есть полярные связи С–О, С–N, –ОН, S–H). Чем больше в белке аминокислот с полярными группами, тем выше его реакционная способность. От реакционной способности во многом зависят функции белка. Особенно большим числом полярных групп, характеризуются ферменты. И наоборот, их очень мало в таком белке как кератин (волосы, ногти).
Аминокислоты классифицируют и на основе ионных свойств R-групп (таблица 1). Кислые (при рН=7 R-группа может нести отрицательный заряд) это аспарагиновая, глутаминовая кислоты, цистеин и тирозин. Основные( при рН =7 R-группа может нести положительный заряд) – это аргинин, лизин, гистидин. Все остальные аминокислоты относятся к нейтральным (группа R незаряжена).
Таблица 1 – Классификация аминокислот на основе полярности R-групп.
| Аминокислоты |
Принятые однобуквенные обозначения и символы |
Изоэлектрическая точка, рI |
Среднее содержание в белках,% |
| Англ. |
символ |
Русск. |
1. НеполярныеR-группы
Глицин
Аланин
Валин
Лейцин
Изолейцин
Пролин
Фенилаланин
Триптофан
2. Полярные, незаряженныеR-группы
Серин
Треонин
Цистеин
Метионин
Аспарагин
Глутамин
3. Отрицательно заряженные R-группы
Тирозин
Аспарагиновая к-та
Глутаминовая к-та
4. Положительно заряженныеR-группы
Лизин
Аргинин
Гистидин
|
GLy
ALa
VaL
Leu
Lie
Pro
Phe
Trp
Ser
Thr
Cys
Met
Asn
GLn
Tyr
Asp
GLy
Lys
Arg
His
|
G
A
V
L
I
P
F
W
S
T
C
M
N
Q
Y
D
E
K
R
N
|
Гли
Ала
Вал
Лей
Иле
Про
Фен
Трп
Сер
Тре
Цис
Мет
Асн
Глн
Тир
Асп
Глу
Лиз
Арг
Гис
|
5,97
6,02
5,97
5,97
5,97
6,10
5,98
5,88
5,68
6,53
5,02
5,75
5,41
5,65
5,65
2,97
3,22
9,74
10,76
7,59
|
7,5
9,0
6,9
7,5
4,6
4,6
3,5
1,1
7,1
6,0
2,8
1,7
4,4
3,9
3,5
5,5
6,2
7,0
4,7
2,1
|
По числу аминных и карбоксильных групп аминокислоты делятся на моноаминамонокарбоновые, содержащие по одной карбоксильной и аминной группе; моноаминодикарбоновые (две карбоксильные и одна аминная группа); диаминомонокарбоновые (две аминные и одна карбоксильная группа).
По способности синтезироваться в организме человека и животных все аминокислоты делятся на заменимые, незаменимые и частично незаменимые.
Незаменимые аминокислоты не могут синтезироваться в организме человека и животных они обязательно должны поступать вместе с пщей. Абсолютно незаменимых аминокислот восемь: валин, лейцин,изолейцин,треонин,триптофан, метионин,лизин, фенилаланин.
Частично незаменимые-синтезируются в организме, но в недостаточном количестве, поэтому частично должны поступать с пищей. Такими аминокислотами являются арганин, гистидин, тирозин.
Заменимые аминокислоты синтезируются в организме человека в достаточном количестве из других соединений. Растения могут синтезировать все аминокислоты.
Кислотно-основные свойства аминокислот связаны с наличием в их структуре двух ионизируемых групп-карбоксильной и аминогруппы, поэтому амнокислоты могут проявлять свойства как кислот, так и оснований, т.е. они являются амфотерными соединениями. В кристаллическом состоянии и в водных растворах a-аминокислоты существуют в виде биполярных ионов, называемых также цвиттерионами. Ионное строение обуславливает некоторые особенности свойств a-аминокислот: высокую температуру плавления (200-300°С), нелетучесть, растворимость в воде и нерастворимость в неполярных органических растворителях. С растворимостью аминокислот в воде связана их всасываемость и транспорт в организме. Ионизация молекул аминокислот зависит от рН раствора. Для моноаминомонокарбоновых кислот процесс диссоциации имеет следующий вид:
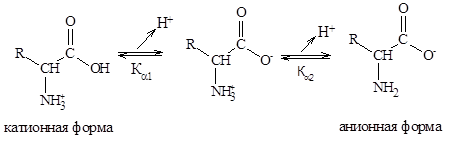
В сильно кислых растворах аминокислоты присутствуют в виде положительных ионов, а в щелочных – в виде отрицательных.
Кислотно-основные свойства аминокислот можно объяснить исходя из теории кислот и оснований Бренстеда-Лоури. Полностью протонированная a-аминокислота (катионная форма) с позиции теории Бренстеда является двухосновной кислотой, содержащей две кислотные группы:недиссоциированную карбоксильную группу (– СООН) и протонированную аминогруппу (NН3
), которые характеризуются соответствующими значениями рКa1
и рКa2
.
Величины рК для аминокислот определяют по кривым титрования. Рассмотрим кривую титрования аланина (рис. 1).
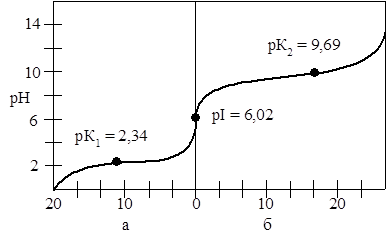
Рис. 1 – кривые, полученные при титровании 0,1М раствора аланина 0,1М раствором HCl (а) и 0,1М растором NaOH (б).
Из кривой титрования аланина следует, что карбоксильная группа имеет рКa1
=2,34, а протонированная аминогруппа рКa2
= 9,69. При рН = 6,02 аланин существует в виде биполярного иона, когда суммарный электрический заряд частицы равен 0. При этом значении рН молекула аланина электронейтральна. Такое значение рН называют изоэлектрической точкой и обозначают рНиэт
или рI. Для моноаминомонокарбоновых кислот изоэлектрическая точка рассчитывается как среднее арифметическое двух значений рКa
. Например для аланина она равна:
рI = Ѕ × (рКa1
+ рКa2
) = Ѕ × (2,34 + 9,69) = 6,02
При значении рН, превышающем изоэлектрическую точку, аминокислота заряжается отрицательно, а при значении рН ниже рI аминокислота несет суммарный положительный заряд. Например, при рН = 1,0 все молекулы аланина существуют в форме ионов
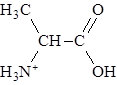
с суммарным зарядом +1. При рН = 2,34, когда имеется смесь равных количеств ионов
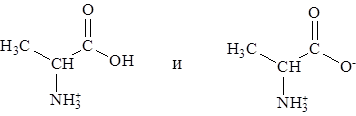
суммарный заряд = +0,5. Аналогичным образом можно предсказать знак и величину суммарного заряда для любой другой аминокислоты при любом значении рН.
Аминокислоты с ионизируемой группой в радикале имеют более сложные кривые титрования, складывающиеся из 3-ох участков, соответствующих трем возможным стадиям ионизации, и, следовательно, они имеют три значения рК (рКa1
, рКa2
и рКR
). Ионизация кислых аминокислот, например аспарагиновой, состоит из следующих последовательных стадий:
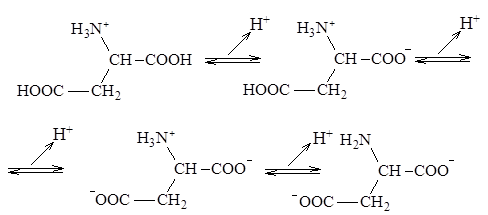
Изоэлектрические точки таких аминокислот определяются также присутствием ионизируемой группой радикала, наряду с a-амино и a-карбоксильными группами. Для моноаминодикарбоновых кислот изоэлектрические точки смещены в кислую область рН и определяются как среднее арифметическое между величинами рК для двух карбоксильных групп (рI аспарагиновой кислоты = 2,97). Для основных аминокислот рI смещены в щелочную область и вычисляются как среднее арифметическое между величинами рК для двух протонированных аминогрупп (рI лизина = 9,74).
Кислотно-основные свойства аминокислот используются для разделения и последующей идентификации аминокислот методами электрофореза и ионообменной хроматографии. Оба эти метода основаны на различиях в знаке и величине суммарного электрического заряда при данном значении рН.
Аминокислоты входящие в состав белка, поглощают свет только в дальней ультрафиолетовой области. Ароматические аминокислоты тирозин, фенилаланин и особенно триптофан поглощают при 260-280 мм. Цистеин обладает слабым поглощением при 240 нм вследствие наличия в нем дисульфидной группы.
Аминокислоты могут участвовать во многих реакциях с участием a-амино, a-карбокси и различных функциональных R-групп. Эти реакции подробно рассмотрены в методическом указании «Химия и биохимия аминокислот и полипептидов». Отметим лишь некоторые реакции, имеющие особо важное значение.
Реакция с нингидрином (рН>5) лежит в основе обнаружения и количественного определения аминокислот и белков.
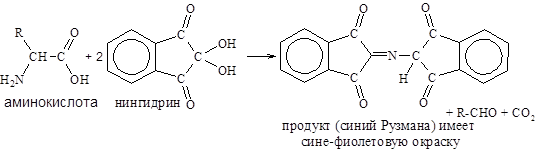
Интенсивность образующейся окраски, оценивают, измеряя поглощение света при длине волны 540 нм (для пролина – 440 нм).
Для идентификации аминокислот служит также получение: 1) фенилтиогитантионовых (ФТТ) производных аминокислот, поглощающих в УФ области; 2) дансильных флуоресцирующих производных (ДНФ) и т.д.
Для пищевиков представляет интерес реакция аминокислот с соединениями, содержащими карбонильную группу С=О, с различными альдегидами и восстанавливающими сахарами (глюкозой, рибозой и отчасти фруктозой). В результате этой реакции происходит разложение как исходной аминокислоты, так и реагирующего с ней восстанавливающего сахара.
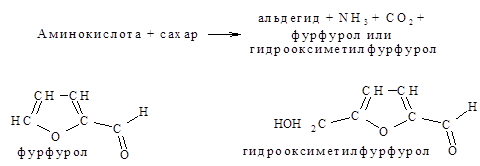
Альдегиды, образовавшиеся из аминокислот, обладают определенным запахом, от которого в значительной степени зависит аромат многих пищевых продуктов.
Лейцин дает запах ржаного хлеба, глицин – карамели, фурфурол – запах зеленого яблока, гидроксиметилфурфурол – запах меда.
Далее фурфурол и гидроксиметилфурфурол реагируют с новой молекулой аминокислоты в результате образуются темноокрашенные соединения – меланоидины. Образование их объясняет наблюдаемое потемнение многих пищевых продуктов во время их изготовления. Особенно интенсивно реакция между аминокислотами и восстанавливающими сахарами происходит при повышенной температуре, имеющей место во время сушки пищевых продуктов, овощей, фруктов, молока, солода. Образование золотисто-коричневой корочки, специфического аромата и вкуса хлеба зависят в основном от меланоидиновых реакций, происходящих при выпечке.
Аминокислоты соединяются друг с другом ковалентной пептидной связью. Образование ее происходит за счет a-аминогруппы (–NH2
) одной аминокислоты и a-карбоксильной (–СООН) группы другой с выделением молекулы воды.
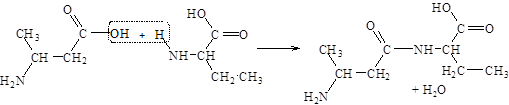
В результате реакции поликонденсации можно получить соединения, составленные из многих аминокислотных остатков – полипептиды. При написании формулы линейных пептидов с известной последовательностью аминокислотных остатков начинают с N-конца (на конце пептида находится свободная a-аминогруппа), используя сокращенные обозначения аминокислот. Названия пептидов складываются из названий соответствующих аминокислот с суффиксом –ил, начиная с N-концевого остатка, – название С-концевой аминокислоты (содержит свободную a-карбоксильную группу) сохраняется. Например, аргинил-аланил-глицин-глутамил-лизин.
Каждый пептид содержит только одну свободную a-амино- и a-карбоксильную группу, которые находятся на концевых остатках аминокислот. Эти группы и R-группы некоторых аминокислот могут быть ионизированы, поэтому пептиды могут нести заряды, и могут быть электронейтральными (т.е. иметь изоэлектрическую точку (ИЭТ). Это свойство пептидов используется для их разделения методами ионной хроматографии и электрофореза. Как и другие соединения, пептиды могут вступать в химические реакции, определяемые наличием у них групп -NH2
, -COOH, и R групп-аминокислот. Одной из важных реакций для пептидов является реакция гидролиза. Реакция гидролиза всех пептидных связей путем кипячения растворов пептидов в присутствии сильной кислоты или щелочи используется при определении их аминокислотного состава и состава белков.
Гидролиз пептидных связей может быть осуществлен также действием некоторых ферментов, которые расщепляют пептидные связи избирательно, с образованием коротких пептидов. Например трипсин гидролизует связи образованные карбоксильными группами лизина, аргинина; химотрипсин-карбоксильными группами фенилаланина, тирозина, триптофана. Такой избирательный анализ оказывается очень полезным при установлении аминокислотной последовательности белков и пептидов.
Кроме пептидов, образующихся в результате частичного гидролиза молекул белка, существует много пептидов, встречающихся в живых организмах как свободные соединения.
Многие природные пептиды отличаются по своей структуре от белков; такие пептиды имеются во всех типах организмов. В структурном отношении пептиды небелковой природы весьма разнообразны: отличаются по размерам, наличию циклических структур, разветвленности, наличию D- и a-аминокислот и, в некоторых отдельных случаях, по уникальному строению пептидной связи. Исходя из принципа взаимосвязи структуры и функций, биологические функции таких пептидов также очень многоплановы. Приведем несколько интересных примеров.
Карнозин и Ансерин. Эти дипептиды найдены в мышечных тканях позвоночных, в том числе и в мышцах человека. Оба они содержат b-аланин – структурный изомер a-аланина.
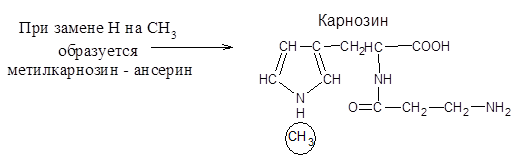
Эти дипептиды служат для поддержания постоянного рН в клетках мышц, т.е действуют как буферы, также они участвуют в сокращении мышц, в процессах окислительного фосфорилирования т.е в образовании АТФ.
Глутатион (g-глутамилцистеинилглицин) – трипептид, присутствует во всех животных, растениях и микроорганизмах.
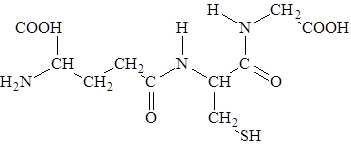
Отличительная структурная особенность глутатиона, состоит в том, что глутаминовая кислота в составе этого пептида представляет для образования пептидной связи g-карбоксильную (а не a-карбоксильную группу). Существуют две формы глутатиона восстановленная (SH-глутатион) и окисленная (S-S-глутатион). Взаимопревращения одной формы в другую катализируется ферментом глутатионредуктазой.
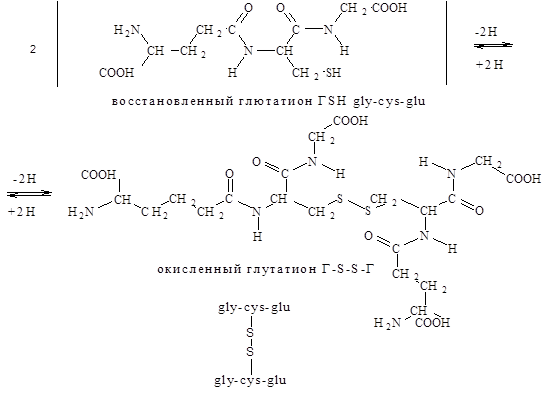
В настоящее время известны лишь некоторые из физиологических функций глутатиона:
1) участие в транспорте аминокислот через клеточные мембраны;
2) поддержание восстановленного состояния железа (Fe+2
) в гемоглабине;
3) входит в состав фермента глутатионпероксидазы, который защищает клетки от разрушающего действия Н2
О2
.
4) участвует в детоксикации ряда чужеродных для живой клетки соединений (галогенсодержащие алифатические или ароматические углеводороды) переводит их в водорастворимые соединения, которые выводятся из организма почками.
5) восстановленный глутатион защищает SH-группы белка от окисления, сам при этом превращается в окисленный.
Глутатион влияет и на технологические свойства зерна и муки. Восстановленный глутатион вызывает восстановление и разрыв дисульфидных связей в молекуле белков клейковины, т.е разрушает ее. Тесто из такой муки обладает плохими структурно-механическими свойствами), оно ослабляется, расплывается из него нельзя получить хлеб нормального качества.) Много глутатиона в старых дрожжах и зародышах зерновых, что следует учитывать в хлебопечении. Восстановленный глутатион способен активировать протеиназы (ферменты ращепляющие белки) зерна и муки, при этом начинается усиленно протекать протеолиз белков клейковины и вызванное им разжижение теста. Глутатион способствует накоплению в пиве азотистых соединений сравнительно большой молекулярной массы, что вызывает образование мути в пиве и ухудшает его потребительские свойства.
С 1981г. разрешено использовать в качестве низкокалорийной добавки для придания продуктам сладкого вкуса аспартам (торговое название). Аспартам в 200 раз слаще сахара и представляет собой метиловый эфир дипептида, состоящего из остатков аспарагиновой кислоты и фенилаланина.
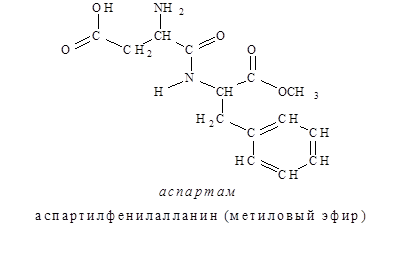
У млекопитающих (в том числе у человека) вырабатываются пептиды обладающие гормональным регуляторным действием, причем диапазон приложения их действия и эффективность в организме очень разнообразны. Например, два циклических нонапептида вырабатывает гипофиз. Окситоцин стимулирует сокращение матки у беременных самок и выделение молока у кормящих самок. Вазопрессин обладает сильным антидиуретическим действием и участвует в контроле кровяного давления. Соматостатин – один из гормонов гипоталамуса – ингибирует синтез гормона роста человека в гипофизе, что приводит к задержке роста и развития тела.
В 1975г. открыта группа пептидов, которые оказывают влияние на передачу нервных импульсов. Их также называют опиатными пептидами, поскольку механизм их действия сходен с механизмом действия морфина и других опиоидов. Они присутствуют в очень малых количествах, как у позвоночных, так и у беспозвоночных. Эти вещества обладают сильным обезболивающим действием, а также участвуют в регуляции настроения и поведения.
Полипептиды, содержащие больше 51 аминокислоты, относятся к белкам. Белки входят в состав всех клеток и тканей живых организмов. Около 50% сухого вещества клетки приходится на белки.
Белки характеризуются определенным элементарным составом. Химический анализ показал наличие во всех белках углерода (50-55%), кислорода (21-24%), азота (15-18%), водорода (6-7%), серы (0,3-2,5%). В составе отдельных белков обнаружены также фосфор, йод, железо, медь и некоторые другие макро и микроэлементы, в различных, часто очень малых количествах.
Белками (протеинами, от греческого protas – первый, важнейший) называют высокомолекулярные природные полимеры, молекулы которых построены из остатков аминокислот.
Поразительно то, что все белки во всех организмах построены их одного и того же набора – 20-ти аминокислот, каждая из которых не обладает никакой биологической активностью. Что же тогда придает белку специфическую активность, одним ферментативную, другим, гормональную, третьим защитную и т.д.
Ответ довольно прост: белки отличаются друг от друга тем, что каждый имеет свою характерную для него аминокислотную последовательность.
Аминокислоты – это алфавит белковой структуры; соединив их в различном порядке можно получить бесконечное число последовательностей, а, следовательно, и бесконечное количество разнообразных белков, выполняющих различные биологические функции.
1. Ферментативная (каталитическая). В биологических системах почти все реакции катализируются специфическими белками – ферментами. В настоящее время открыто около 300 различных ферментов, каждый из которых служит катализатором определенной биологической реакции. Синтез и распад веществ, их регуляция, перенос химических групп и электронов от одного вещества к другому осуществляется с помощью ферментов.
2. Строительная, структурная функция. Белки образуют основу протоплазмы любой живой клетки, в комплексе с липидами они являются основным структурным материалом всех клеточных мембран всех органелл.
3. Двигательная функция. Любые формы движения в живой природе (работа мышц, движение ресничек и жгутиков у простейших, движение протоплазмы в клетке и т.д.) осуществляется белковыми структурами.
4. Транспортная функция. Перенос различных молекул, ионов осуществляется специфическими белками. Например, белок крови гемоглобин переносит кислород к тканям. Перенос жирных кислот по организму происходит с участием другого белка крови-альбумина.
5. Регуляторная функция. Регуляция углеводного, белкового, липидного обменов осуществляется с помощью гормонов, которые по своему строению относятся к белкам (инсулин) или пептидам (окситоцин, вазопрессин и др.).
6. Защитная – эту функцию выполняют иммуноглобулины (антитела). Они обладают способностью обезвреживать бактерии, вирусы, чужеродные белки, попавшие в организм извне. Процесс свертывания крови, защищающий организм от ее потери, основан на превращениях белка – фибриногена. Кератин – белок волосяного защитного покрова.
7. Фоторецепторные белки: например, родопсин, участвующий в зрительных процессах.
8. Резервные белки используются, как запасной материал для питания развивающегося зародыша и новорожденного организма – это белки семян зернобобовых культур, альбумин – яичный белок, казеин молока. Ферретин – белок животных тканей в котором запасено железо. Резервные белки являются важнейшими компонентами растительной и животной пищи.
Имеется много других белков, функции которых довольно необычны. Например, монеллин – белок, выделенный из африканского растения, имеет очень сладкий вкус. Его изучают как вещество нетоксичное и не способствующее ожирению, с целью использования в пищу вместо сахара. Плазма крови некоторых антарктических рыб содержи белок, обладающий свойствами антифриза.
Технология многих производств основана на переработке белков, изменении их свойств; в кожевенной промышленности, при выделке мехов, натурального шелка, выработке сыров, хлеба и т.д.
Для описания строения белковой молекулы были введены понятия о первичной, вторичной, третичной и четвертичной структурах белковой молекулы. В последние годы появились еще такие понятия, как сверхвторичная структура, характеризующая энергетически предпочтительные агрегаты вторичной структуры, и домены – части белковой глобулы, представляющие собой достаточно обособленные глобулярные области.
Количество и последовательность расположения аминокислот, и местоположение дисульфидных связей в полипептидной цепи определяют первичную структуру белка. Между первичной структурой белка и его функцией у данного организма существует самая тесная связь. Для того, чтобы белок выполнял свойственную ему функцию, необходима совершенно определенная последовательность аминокислот в полипептидной цепи этого белка. Даже небольшие изменения в первичной структуре могут значительно изменять свойства белка и соответственно его функции. Например, в эритроцитах здоровых людей содержится белок– гемоглобин с определенной последовательностью аминокислот. Небольшая часть людей имеет врожденную аномалию структуры гемоглобина: их эритроциты содержат гемоглобин, у которого в одном положении вместо глутаминовой кислоты (заряженной, полярной) содержится аминокислота валин (гидрофобная, неполярная). Такой гемоглобин существенно отличается по физико-химическим и биологическим свойствам от нормального. Появление гидрофобной аминокислоты, приводит к возникновению «липкого» гидрофобного контакта (эритроциты плохо передвигаются в кровеносных сосудах), к изменению формы эритроцита (из двояковогнутого в серповидный), а также к ухудшению переноса кислорода и т.д. Дети, родившееся с этой аномалией, в раннем детстве погибают от серповидноклеточной анемии.
Исчерпывающие доказательства в пользу утверждения, что биологическая активность определяется аминокислотной последовательностью, были получены, после искусственного синтеза фермента рибонуклеазы (Меррифилд). Синтезированный полипептид с той же аминокислотной последовательностью, что и естественный фермент, обладал такой же ферментативной активностью.
Исследования последних десятилетий показали, что первичная структура закреплена генетически и в свою очередь определяет вторичную, третичную и четвертичную структуры белковой молекулы и ее общую конформацию. Первым белком, у которого была установлена первичная структура, был белковый гормон инсулин (содержит 51 аминокислоту). Это было сделано в 1953 г. Фредериком Сэнгером. К настоящему времени расшифрована первичная структура более десяти тысяч белков, но это очень небольшое количество, если учесть, что в природе белков около 1012
.
Зная первичную структуру белка, можно точно написать его структурную формулу, если белок представлен одной полипептидной цепью. Если в состав белка входит несколько полипептидных цепей, то их предварительно разъединяют, используя специальные реактивы. Для определения первичной структуры отдельной полипептидной цепи, методами гидролиза с использованием аминокислотных анализаторов, устанавливают ее аминокислотный состав. Затем, применяя специальные методы и реагенты, определяют природу концевых аминокислот. Для установления порядка чередования аминокислот, полипептидную цепь подвергают ферментативному гидролизу, при котором образуются осколки этой полипептидной цепи – короткие пептиды. Эти пептиды разделяют методом хроматографии и устанавливают последовательность аминокислот в каждом. Таким образом, достигается этап, когда последовательность аминокислот в отдельных пептидах (фрагментах белка) известна, но остается невыясненной последовательность самих пептидов. Последнюю устанавливают с помощью так называемых перекрывающихся пептидов. Для этого используются какой-либо другой фермент, расщепляющий исходную полипептидную цепь в других участках, и определяют аминокислотную последовательность вновь полученных пептидов. Пептиды, образованные под действием двух ферментов, содержат одинаковые фрагменты аминокислотных последовательностей., совмещая их устанавливают общую аминокислотную последовательность полипептидной цепи.
Большой вклад в изучение строения белковой молекулы сделали Л. Полинг и Р. Кори. Обратив внимание на то, что в молекуле белка больше всего пептидных связей, они первыми провели кропотливые рентгеноструктурные исследования этой связи. Изучили длины связей, углы под которыми располагаются атомы, направление расположения атомов относительно связи. На основании исследований были установлены следующие основные характеристики пептидной связи.
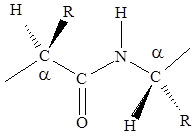
1. Четыре атома пептидной связи и два присоединенных a-углеродных атома лежат в одной плоскости. Группы R и Н a-углеродных атомов лежат вне этой плоскости.
2. Атомы О и Н пептидной связи и два a-углеродных атома и R-группы имеют транс-ориентацию относительно пептидной связи.
3. Длина связи С-N, равная 1,32 Е, имеет промежуточное значение между длиной двойной ковалентной связи (1,21 Е) и однородной ковалентной связи (1,47 Е). Отсюда следует, что связь С-N имеет частично характер двойной связи. Т.е. пептидная связь может существовать в виде резонансных и таутамерных структур, в кето-енольной форме.
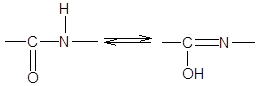
Вращение вокруг связи –С=N– затруднено и все атомы, входящие в пептидную группу, имеют планарную транс-конфигурацию. Цис-конфигурация является энергетически менее выгодной и встречается лишь в некоторых циклических пептидах. Каждый планарный пептидный фрагмент содержит две связи с a-углеродными атомами, способными к вращению. Это связи Сa
–N (угол вращения вокруг этой связи обозначается j) и связь Сa
–С (угол вращения вокруг этой связи обозначается y).
Пептидная связь по своей химической природе является ковалентной и придает высокую прочность первичной структуре белковой молекулы. Являясь повторяющимся элементом полипептидной цепи и имея специфические особенности структуры, пептидная связь влияет не только на форму первичной структуры, но и на высшие уровни организации полипептидной цепи.
Вторичная структура белковой молекулы образуется в результате того или иного вида свободного вращения вокруг связей, соединяющих a-углеродные атомы в полипептидной цепи.
В природных полипептидных цепях обнаружены три основных типа структуры: a-спираль, складчатый лист и статистический клубок. Спиральная структура образуется если в цепи одинаковые углы поворотов (j) для всех связей Сa
–N и углом поворота (y) для всех связей Сa
–С и равны соответственно –48є и –57є. Наиболее часто встречается правозакрученная a-спираль. Эта структура очень стабильна, т.к. в ней почти или полностью отсутствуют стерические затруднения, особенно для R-групп боковых цепей аминокислот. R-группы аминокислот направлены наружу от центральной оси a-спирали. В a-спирали диполи =С=О и >N–Н соседних пептидных связей ориентированы оптимальным образом (почти коаксиальны) для дипольного взаимодействия, образуя вследствие этого обширную систему внутримолекулярных кооперативных водородных связей, стабилизирующих a-спираль. Шаг спирали (один полный виток) 5,4Е включает, 3,6 аминокислотных остатка.

Рисунок 1 – Структура и параметры a-спирали белка
Спиральную структуру могут нарушить два фактора:
1) в наличие остатка пролина, циклическая структура которого вносит излом в пептидную цепь – нет группы –NН2
, поэтому невозможно образования внутрицепочечной водородной связи;
2) если в полипептидной цепи подряд расположено много остатков аминокислот, имеющих положительный заряд (лизин, аргинин) или отрицательный заряд (глутаминовой, аспарагиновой кислот), в этом случае сильное взаимное отталкивание одноименнозаряженных групп (–СОО–
или –NН3
+
) значительно превосходит стабилизирующее влияние водородных связей в a-спирали.
Структура типа складчатого листа также стабилизирована водородными связями между теми же диполями =NН......
О=С<. Однако в этом случае возникает совершенно иная структура, при которой остов полипептидной цепи вытянут таким образом, что имеет зигзагообразную структуру. Углы вращения для связей Сa
-N(
j)
и Сa
-С (
y)
близки соответственно к –120+1350
. Складчатые участки полипептидной цепи проявляют кооперативные свойства, т.е. стремятся расположиться рядом в белковой молекуле, и формируют параллельные
одинаковонаправленные полипептидные цепи или антипараллельные,
которые укрепляются благодаря водородным связям между этими цепями. Такие структуры называются b-складчатые листы (рисунок 2).
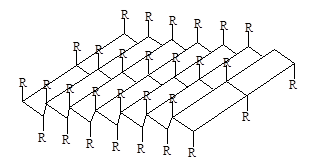
Рисунок 2 – b-структура полипептидных цепей
a-Спиральные складчатые листы – это упорядоченные структуры, в них имеется регулярная укладка аминокислотных остатков в пространстве. Участки белковой цепи с нерегулярной укладкой аминокислотных остатков в пространстве, которые также удерживаются благодаря водородным связям – называются неупорядоченными, бесструктурными – статистическим клубком. Все эти структуры возникают самопроизвольно и автоматически вследствие того, что данный полипептид имеет определенную аминокислотную последовательность, которая предопределена генетически. a-спирали и b-структуры обуславливают определенную способность белков к выполнению специфических биологических функций. Так, a-спиральная структура (a-кератин) хорошо приспособлена к тому, чтобы образовывать наружные защитные структуры-перья, волосы, рога, копыта. b-структура способствует образованию гибких и нерастяжимых нитей шелка и паутины, а конформация белка коллагена обеспечивает высокую прочность на разрыв, необходимую для сухожилий. Наличие только a-спиралей или b-структур характерно для нитевидных-фибрилярных белков. В составе глобулярных-шаровидных белков содержание a-спиралей и b-структур и бесструктурных участков сильно варьирует. Например: инсулин спирализован-на 60%, фермент рибонуклеаза – 57%, белок куриного яйца лизоцим – на 40%.
Сведения о чередовании аминокислотных остатков в полипептидной цепи, а также о наличии в белковой молекуле спирализованных, складчатых и неупорядоченных участков еще не дают полного представления ни об объеме, ни о форме, ни тем более о взаимном расположении участков полипептидной цепи по отношению друг к другу.
Эти особенности строения белка выясняются при изучении его третичной структуры, под которой понимают общее расположение в пространстве в определенном объеме полипептидной цепи.
Третичная структура устанавливается с помощью рентгеноструктурного анализа. Первая модель молекулы белка – миоглобина, отражающая его третичную структуру, была создана Дж. Кендрю с сотрудниками в 1957г. Несмотря на большие трудности к настоящему времени удалось установить третичную структуру более 1000 белков, в том числе гемоглобина, пепсина, лизоцима, инсулина и т.д.
Третичная структура белков образуется путем дополнительного складывания пептидной цепи содержащей a-спираль, b-структуры и участки без периодической структуры. Третичная структура белка формируется совершенно автоматически, самопроизвольно и полностью предопределяется первичной структурой. Основной движущей силой в возникновении трехмерной структуры, является взаимодействие радикалов аминокислот с молекулами воды. При этом неполярные гидрофобные радикалы аминокислот группируются внутри белковой молекулы, в то время как полярные радикалы ориентируются в сторону воды. В какой-то момент возникает термодинамически наиболее выгодная стабильная конформация молекулы – глобула. В такой форме белковая молекула характеризуется минимальной свободной энергией. На конформацию возникшей глобулы оказывают влияние такие факторы как рН раствора, ионная сила раствора, а также взаимодействие белковых молекул с другими веществами.
В последнее время появились доказательства, что процесс формирования третичной структуры не является автоматическим, а регулируется и контролируется специальными молекулярными механизмами. В этом процессе задействованы специфические белки – шапероны. Основными функциями их являются способность предотвращать образование из полипептидной цепи неспецифических (хаотичных) беспорядочных клубков, и обеспечение доставки (транспорта) их к субклеточным мишеням, создавая условия для завершения свертывания белковой молекулы.
Стабилизация третичной структуры обеспечивается благодаря нековалентным взаимодействиям между атомными группировками боковых радикалов следующих типов:
- водородные связи могут возникать между функциональными группами боковых радикалов. Например, между ОН группой тирозина и –N< в кольце остатка гистидина.
- электростатические силы притяжения между радикалами, несущими противоположно заряженные ионные группы (ион-ионные взаимодействия), например отрицательно заряженная карбоксильная группа (– СОО–
) аспарагиновой кислоты и (NН3
+
) положительно заряженной e-аминогруппой остатка лизина.
- гидрофобные взаимодействия обусловлены силами Ван-дер-Ваальса между неполярными радикалами аминокислот. (Например, группами –СН3
– аланина.
Стабилизируется третичная структура и ковалентной дисульфидной связью (–S–S–) между остатками цистеина. Эта связь очень прочная и присутствует не во всех белках. Важную роль эта связь играет в белковых веществах зерна и муки, т.к. оказывает влияние на качество клейковины, структурно-механические свойства теста и соответственно на качество готовой продукции – хлеба и т.д.
Белковая глобула не является абсолютно жесткой структурой: в известных приделах возможны обратимые перемещения частей пептидной цепи относительно друг друга с разрывом небольшого количества слабых связей и образования новых. Молекула как бы дышит, пульсирует в разных своих частях. Эти пульсации не нарушают основного плана конформации молекулы, подобно тому, как тепловые колебания атомов в кристалле не изменяют структуру кристалла, если температура не настолько велика, что наступает плавление.
Только после приобретения белковой молекулой естественной, нативной третичной структуры он проявляет свою специфическую функциональную активность: каталитическую, гормональную, антигенную и т.д. Именно при образовании третичной структуры происходит формирование активных центров ферментов, центров ответственных за встраивание белка в мультиферментный комплекс, центров, ответственных за самосборку надмолекуляных структур. Поэтому любые воздействия (термические, физические, механические, химические), приводящие к разрушению этой нативной конформации белка (разрыв связей), сопровождается частичной или полной потерей белком его биологических свойств.
Изучение полных химических структур некоторых белков показало, что в их третичной структуре выявляются зоны, где сконцентрированы гидрофобные радикалы аминокислот, и полипептидная цепь фактически обматывается вокруг гидрофобного ядра. Более того, в ряде случаев в белковой молекуле обособляются два и даже три гидрофобных ядра, в результате возникает 2-х или 3-х ядерная структура. Такой тип строения молекулы характерен для многих белков, обладающих каталитической функцией (рибонуклеаза, лизоцим и т.д.). Обособленная часть или область молекулы белка обладающая в определенной степени структурной и функциональной автономией называется доменом. У ряда ферментов, например, обособленны субстрат-связывающие и кофермент связывающие домены.
Третичная структура белка имеет прямое отношение к его форме, которая может быть различной: от шарообразной до нитевидной. Форма белковой молекулы характеризуется таким показателем, как степень асимметрии (отношение длинной оси к короткой). У фибриллярных или нитевидных белков степень асимметрии больше 80. При степени асимметрии меньше 80 белки относятся к глобулярным. Большинство из них имеет степень асимметрии 3-5, т.е. третичная структура характеризуется достаточно плотной упаковкой полипептидной цепи, приближающейся по форме к шару.
В биологическом отношении фибриллярные белки играют очень важную роль, связанную с анатомией и физиологией животных. У позвоночных на долю этих белков приходится 1/3 от их общего содержания. Примером фибрилярных белков может служить белок шелка – фиброин, который состоит из нескольких антипараллельных цепей со структурой складчатого листа. Белок a-кератин содержит от 3-7 цепей. Коллаген имеет сложную структуру, в которой 3 одинаковые левовращающие цепи скручены вместе с образованием правовращающей тройной спирали. Эта тройная спираль стабилизирована многочисленными межмолекулярными водородными связями. Наличие таких аминокислот, как гидроксипролина и гидроксилизина также вносит вклад в образование водородных связей, стабилизирующих структуру тройной спирали. Все фибриллярные белки плохо растворимы или совсем нерастворимы в воде, так как в их составе содержится много аминокислот, содержащих гидрофобные, нерастворимые в воде R-группы изолейцин, фенилаланин, валин, аланин, метионин. После специальной обработки нерастворимый и неперевариваемый коллаген превращается в желатин-растворимую смесь полипептидов, который затем используют в пищевой промышленности.
Глобулярные белки выполняют разнообразные биологические функции. Они выполняют транспортную функцию, т.е. переносят питательные вещества, неорганические ионы, липиды и т.д. К этому же классу белков принадлежат гормоны, а также компоненты мембран и рибосом. Все ферменты тоже глобулярные белки.
Белки содержащие две или большее число полипептидных цепей называют олигомерными белками для них характерно наличие четвертичной структуры. Полипептидные цепи (промеры) в таких белках могут быть либо одинаковыми либо разными. Олигомерные белки называют гомогенными, если их протомеры одинаковы и гетерогенными, если их протомеры различны. Например-белок гемоглобин состоит из 4-х цепей: двух -a и двух -b протомеров. Фермент a-амилаза состоит из 2-х одинаковых полипептидных цепей. В олигомерных белках каждая из полипептидных цепей характеризуется своей вторичной и третичной структурой, и называется субъединицей или протомером. Протомеры взаимодействуют друг с другом не любой частью своей поверхности, а определенным участком (контактной поверхностью). Контактные поверхности имеют такое расположение атомных группировок, между которыми возникают водородные, ионные, гидрофобные связи. Кроме того, геометрия протомеров также способствует их соединению. Протомеры подходят друг к другу, как ключ к замку. Такие поверхности называются комплиментарными. Каждый протомер взаимодействует с другим во множестве точек, это приводит к тому, что соединение с другими полипептидными цепями или белками невозможно. Такие комплиментарные взаимодействия молекул лежат в основе всех биохимических процессов в организме. Под четвертичной структурой понимают расположение полипептидных цепей (протомеров) относительно друг друга, т.е. способ их совместной укладки и упаковки с образованием нативной конформации олигомерного белка, в результате чего белок обладает той или иной биологической активностью.
Белки, благодаря присутствию в их составе ионных и полярных группировок (–NH2
; –COOH; –SH; –OH и т.д.) существуют в водных растворах в виде заряженных частиц. В зависимости от соотношения в белке основных (NH-аминных) и кислых (–СООН карбоксильных) группировок и рН среды молекула белка в водном растворе приобретает положительный или отрицательный заряд. Большинство белков животного происхождения содержат в своем составе больше дикарбоновых аминокислот аспарагиновой и глютаминовой и поэтому в водных растворах они заряжаются отрицательно (белки-анионы). Некоторые белки содержат в своем составе значительные количества диаминокислот (аргинина, лизина, гистидина) и поэтому заряжаются положительно (белки-катионы). Одноименный заряд молекул способствует взаимному отталкиванию частиц, что обеспечивает устойчивость их в водном растворе.
Число ионизированных групп в белке может быть увеличено или уменьшено при изменении рН среды. В кислой среде подавляется диссоциация карбоксильных групп и отрицательный суммарный заряд белка уменьшается. Наоборот, в щелочной среде подавляется ионизация аминных групп и положительный суммарный заряд белка уменьшается. При определенном значении рН число положительных зарядов на поверхности белковой молекулы будет равным числу отрицательных зарядов и, в целом заряд молекулы белка станет равным нулю. Состояние белка, при котором суммарный заряд его равен нулю, называется изоэлектрическим состоянием. рН, при котором белок находится в изоэлектрическом состоянии называется изоэлектрической точкой белка и обозначается рI. В изоэлектрической точке отсутствие заряда у молекул белка ослабляет силы отталкивания между белковыми частицами, что благоприятствует агрегации белковых молекул и выпадению белка в осадок, т.е. в изоэлектрической точке раствор белка неустойчив, так как белок теряет один из факторов стабилизации белковых водных растворов – заряд.
При добавлении щелочи или кислоты к белку, выпавшему в осадок в изоэлектрическом состоянии, наступает перезарядка его молекул, и белок вновь переходит в раствор – растворяется.
Белки сохраняют свою нативную структуру, а, следовательно, выполняют свойственную им функцию только при определенных физико-химических параметрах среды. Если каким-то образом разрушить связи стабилизирующие пространственную структуру (вторичную, третичную, четвертичную) белковой молекулы, то упорядоченная, уникальная для каждого белка конформация пептидной цепи нарушается и молекула целиком или в значительной части принимает форму случайного беспорядочного клубка. Такое изменение белка называют денатурацией. При денатурации белка первичная структура сохраняется.
Денатурацию можно вызвать действием физических и химических факторов. Высокая температура (выше 60є) вызывает разрушение в белке гидрофобных и водородных связей.
Скорость тепловой денатурации зависит от температуры, влажности, (сухие белки более устойчивы к денатурации) присутствия солей, их концентрации. Тепловая денатурация сопровождается агрегацией белков, выпадением их в осадок, что представляет собой уже вторичное явление. Типичным примером денатурации может служить свертывание белков куриного яйца при нагревании.
Белки денатурируют и при многих механических воздействиях: растирании, энергичном встряхивании, при облучении ультрафиолетовом, высокочастотном и т. д. При лиофилизации большинство белов не денатурирует, поэтому этот способ сушки используется для получения белковых препаратов.
Действие кислот и щелочей так же вызывает денатурацию вследствие перезарядки и изменения диссоциации ионогенных аминокислот. При сильном подкислении или подщелачивании все диссоциирующие группы белка имеют одноименный заряд (преимущественно NН3
+
-групп при низких значениях рН и –СОО–
– при высоких). Взаимное отталкивание одноименных зарядов вызывает разрыв части слабых связей, в результате чего нарушается нативная структура. Однако имеются белки, которые устойчивы при крайних значениях рН. Например, пепсин при рН 1,5-2 имеет максимальную активность, у аргиназы оптимум рН 9,5-9,7.
Денатурация может быть вызвана катионами тяжелых металлов и некоторыми анионами – иода, тиоционата и др. Считают, что при этом образуются прочные комплексные соединения и происходит нарушение собственных связей в белке ионных, водородных, гидрофобных.
Действие органических растворителей вызывает нарушение гидрофобных взаимодействий в молекулах белка.
Полярные денатурирующие агенты низкие спирты, этиленгликоль, диоксан и др. образуют водородные связи с аминогруппами или карбоксильными группами пептидного остова и с некоторыми группами радикалов аминокислот, подменяя собственные внутримолекулярные водородные связи в белке, вследствие чего четвертичная, третичная и вторичная структуры частично или полностью разрушаются.
Некоторые химические соединения (глицерин, глюкоза и др. сахара) оказывают на белки защитное действие. Считают, что это связано с их адсорбцией на глобулах белков и образованием крупных гидрофильных комплексов. Денатурация белков сопровождается изменением их физико-химических свойств. Снижается растворимость, гидрофильность белка, его вязкость, оптические свойства. Денатурированные белки дают более интенсивные цветные реакции на аминокислоты, т.к. они стали более доступными вследствие развертывания полипептидной цепочки. «Обнажаются» при этом и пептидные связи, поэтому протеолиз денатурированных белков ферментами протекает с большей скоростью, чем нативных.
При денатурации белок теряет свою биологическую активность, что выражается в неспособности ферментов, гормонов, вирусов выполнять свои функции т. е. они инактивируются.
В определенных условиях, при удалении денатурирующего агента или при медленном охлаждении раствора белка денатурированного нагреванием происходит ренативация – восстановление исходной (нативной) конформации и специфической биологической функции белка.
Процесс денатурации белков играет большую роль в технологии пищевых продуктов при их тепловой обработке. Так потеря всхожести и ухудшение хлебопекарных свойств, происходящее вследствие перегрева зерна при его неправильной сушке в зерносушилке, является следствием денатурации белков. Процесс глубокой денатурации происходит при выпечке хлеба. Копчение мяса при температуре (40-90єС) сопровождается частичной денатурацией белков и освобождением скрытых функциональных групп (–SH, –COOH, –NH2
, –OH–
, =С=О и др.), которые могут вступать во взаимодействие с летучими продуктами коптильных газов. В результате всех перечисленных процессов происходит необратимая дегидратация, коагуляция части белков, в связи с чем уменьшается влагоудерживающая способность ткани, продукт лучше обезвоживается и уплотняется.
Бланшировка плодов и овощей вызывает денатурацию окислительных ферментов – пероксидазы, о–дифенолоксидазы, аскарбинатоксидазы и т.д. что ведет к сохранению витамина С и препятствует образованию темноокрашенных соединений – меланинов. Тепловая обработка пищевых продуктов повышает их вкусовые качества и усвояемость, поскольку денатурированные белки обладают лучшей атакуемостью пищеварительными ферментами.
Белковая глобула обладает гидрофильностью. Водная оболочка белковой молекулы возникает в результате взаимодействия гидрофильных групп белковой молекулы с диполями воды с образованием водородных связей. Так, пептидная группировка притягивает одну молекулу воды, аминогруппа –NH2
– тоже одну, карбоксильная группа –СООН – четыре. Вблизи молекулы белка отмечается упорядоченное расположение молекул воды. По мере удаления от молекул белка расположение молекул воды носит все более беспорядочный характер. Водная оболочка является одним из факторов устойчивости белковых растворов – препятствует слипанию молекул белка и их осаждению.
При контакте сухого белка с водой он набухает, молекулы воды проникают в белковую массу, гидратируют молекулы белка, разъединяя их. Важную роль в этом процессе играют не только электростатические силы, но и силы осмоса. Дальнейшее поглощение воды приводит к растворению белка.
При определенных условиях белковые растворы образуют коллоидные системы – гели или студни, в которых растворитель и белок образуют одну внешне гомогенную массу. В гелях имеется гидратационная вода, окружающая толстым слоем коллоидные частицы белка, а так же вода, удерживаемая в капиллярных пространствах между ними. Высушенный гель, помещенный в воду, впитывает ее в очень больших количествах – набухает.
Гидрофильные свойства белков, т.е. их способность образовывать студни, набухать имеет большое значение в биологии и пищевых производствах. Очень подвижным студнем, является цитоплазма – полужидкое содержимое клетки, хрусталик глаза и т.д. Типичным белковым сильногидраторованным гелем является пшеничная клейковина, она содержит около 66% воды. Гидрофильные, водоудерживающие свойства продуктов необходимо так же учитывать при замораживании и размораживании мяса, овощей, фруктов, что бы сохранить, не ухудшить их качество. Гидрофильность белков зерна и муки играет большую роль при хранении и переработке зерна (кондиционирование, прорастание зерна) и хлебопечении. Студни используются в процессе приготовления различных заливных блюд.
Если у белковых глобул отнять гидратную оболочку, то частицы белка начнут слипаться, образуя ассоциации частиц белка, и осядут. В изоэлектрической точке белки обладают наименьшей способностью связывать воду, поэтому их легче осаждать. Агрегация белковых молекул происходит и при их обезвоживании с помощью некоторых органических растворителей – например спиртом и ацетоном. Молекулы этих веществ, являясь более гидрофильными, чем молекулы белка, образуют собственные гидраты с водой, оттягивая воду от белка, лишая их водной оболочки способствуют агрегации белка.
Обезвоживание белков можно проводить путем добавления нейтральных растворов солей высокой концентрации. Этот процесс называют высаливанием. Осаждающая способность соли зависит от размеров катиона и аниона, а так же от величины их заряда. Катионы и анионы по высаливающей способности можно расположить в ряды, в которых эта способность убывает.
Катионы: Cs+
, Rb+
, K+
, Na+
, Li+
, Ba2+
, Sr2+
, Co2+
, Mg2+
.
Анионы: SO4
-2
, CL-
, Br, NO3
-
, J-
, CNS-
, Mg 2+
.
Эти ряды называются лиотропными рядами. Высаливающее действие объясняется тем, что при высокой концентрации ионов в растворе белка, они оттягивают на себя от молекул белка поляризованные молекулы воды и тем самым лишают белок гидратной оболочки, которая препятствует осаждению белка. Метод высаливания используется для разделения и получения в очищенном виде белков и ферментов.
При добавлении растворов нейтральных солей (Na2
SO4
, (NH4
)2
SO4
, MgSO4
и др.) небольших концентраций, растворимость белков в воде возрастает. Растворению белков, как и других веществ, способствуют те факторы, которые уменьшают взаимодействие между молекулами растворяемого вещества. Нейтральные соли в малых концентрациях увеличивают степень диссоциации ионизированных групп белковых молекул и тем самым уменьшают белок-белковое взаимодействие. Известно, что степень диссоциации электролитов ( в том числе белков) прямо пропорциональна диэлектрической постоянной растворителя, которая в свою очередь пропорциональна степени поляризации молекул растворителя, их дипольному моменту. Нейтральные соли в малых концентрациях еще больше увеличивают диэлектрическую постоянную воды. В результате вода усиливает диссоциацию растворенного вещества, в частности белка. Входя между заряженными группами и ориентируясь вокруг них, диполи воды препятствуют их взаимодействию.
Растворимость белков зависит так же от рН растворителя, его состава, температуры. Минимальной растворимостью обладают белки в ИЭТ, что объясняется отсутствием электростатического отталкивания между молекулами белка.
Белки способны образовывать высококонцентрированные системы жидкось-газ-пены. В качестве традиционных пенообразователей используют белки сыворотки крови, молока, которые подвергают вначале гидролизу, а затем сушат на распылительных сушках. Устойчивость пены, в которой белок является пенообразователем зависит от его природы, концентрации и температуры.
Белки в качестве пенообразователей играют важную роль при образовании пены в пиве. В кондитерской промышленности это свойство белков используется при выработке пастилы, зефира, суфле. Структуру пены имеет хлеб и это влияет на его органолептические и структурно-механические свойства.
Благодаря гидрофильным и гидрофобным группировкам белки являются поверхностно-активными веществами и могут влиять на растворимость других веществ. Они выступают в роле эмульгаторов – веществ, стабилизирующих эмульсию, которую образуют взаимнонерастворимые жидкости (вода-жир). В организме человека в эмульгированном состоянии находятся жиры в крови и лимфе. Молоко представляет собой эмульгированные казеиногеном капельки жира в воде. Белки в качестве эмульгаторов находят применение в пищевой промышленности при производстве майонезов, кондитерских изделий, кремов, шоколада и т.п.
Выделение белков из биологических тканей проводят после тщательного измельчения исследуемого материала, вплоть до разрушения клеточной структуры. Разрушают клетки механическим путем, растирая ткань с песком в ступке или в гомогенизаторе. Существуют и другие методы, например поочередное замораживание и оттаивание, обработка ультразвуком.
На всех этапах выделения и очистки белков следует учитывать их большую способность к потере природных, нативных свойств, т.е. к денатурации.
В большинстве случаев процесс разрушения клеток сопровождается выделением тепла, поэтому с целью предотвращения тепловой денатурации все операции следует проводить при пониженных температурах (около +4°С) в термостатированных холодных комнатах.
Современные методы измельчения тканей обычно сочетают с одновременной экстракцией белков из гомогенатов тканей. В качестве растворителей используют 8-10% растворы солей, различные буферные растворы, органические растворители (спирт, ацетон и т.п.), а так же неионные детергенты –вещества, разрушающие гидрофобные взаимодействия между белками и липидами и между белковыми молекулами.
После достижения полной экстракции белков, приступают к разделению-фракционированию смеси белков на индивидуальные белки. Для этого применяют различные методы: высаливание, осаждение органическими растворителями, хроматографию, электрофорез.
При выделении и очистке белков используют четыре основных вида хроматографии: адсорбционную, распределительную, ионообменную и аффинную (хроматографию по сродству). В адсорбционной хроматографии разделение компонентов смеси основано на их различной сорбируемости на твердом адсорбенте. В качестве адсорбентов используют активированный древесный уголь, оксиды алюминия или кремния. Адсорбент в виде суспензии с растворителем (чаще всего буферным раствором) вносят в колонку и равномерно утрамбовывают. Исследуемый образец в небольшом объеме растворителя вносят в колонку. Компоненты разделяемой смеси адсорбируются на адсорбенте. Затем приступают к десорбции компонентов из колонки, используя подходящие элюенты. Сбор фракций осуществляют при помощи автоматического коллектора фракций.
При распределительной хроматографии твердая фаза служит только опорой (основой) для стационарной жидкой фазы. Разновидностью распределительной хроматографии является хроматография на бумаге. В качестве стационарной фазы при этом служит вода, адсорбированная целлюлозными цепями фильтровальной бумаги. Образец наносят в виде капли (пятна) на одном конце бумажной полосы, этим же концом бумагу погружают в подходящую смесь органических растворителей (например, бутанол, уксусная кислота, вода в определенных соотношениях). При движении растворителя по бумаге благодаря силе капиллярности происходит разделение компонентов смеси. Проявленную хроматограмму высушивают, а местоположение каждого из разделяемых веществ определяют химическими или физико-химическими методами.
В ионообменной хроматографии в зависимости от заряда разделяемых белков используют подходящую ионообменную смолу (катионит или анионит) с функциональными группами которой обмениваются и задерживаются на колонке часть белков, в то время как другие белки беспрепятственно элюируются из колонки. Связанные с ионообменной смолой белки, отделяют, применяя более концентрированные солевые растворы или изменяя рН элюента.
Аффинная хроматография основана на принципе избирательного взаимодействия белков (или других молекул) с закрепленными на носителе специфическими веществами – лигандами, которыми могут быть субстраты или коферменты (если выделяется какой-либо фермент) и т.д. Благодаря высокой специфичности белков к иммобилизованному (закрепленному) лиганду, к нему присоединяется только один какой-либо белок из смеси. Снятие с колонки этого белка осуществляется подобранными специальными элюентами.
В гель-хроматографии в качестве стационарной фазы используют гель в виде крошечных гранул, так что такую стационарную фазу можно рассматривать как молекулярные сита. Гранулы геля изготовлены из полимера со сшитой структурой, подобной ситу. В качестве такого полимерного материала используются или сшитая агароза, или декстран (полисахарид), или сшитый полиакриламид. В водной среде полимерный материал сорбирует воду и набухает, превращаясь в гелеподобные гранулы, сохраняющие пористую структуру, причем размер пор такого набухшего материала определяется степенью сшивки полимера. Нанесенные на колонку соединения (в виде раствора в подвижной фазе) начинают взаимодействовать с гранулами геля, проникая в объем гранул через поры, что замедляет движение растворенного вещества по колонке. Молекулы небольшого размера лучше задерживаться на колонке, поскольку легче проникают в объем гранул и распределяются там. Молекулы с размерами, большими, чем размеры пор, совсем не будут проникать внутрь гранул, они первыми вымываются из колонки.
Результаты хроматографического разделения представляются графически в виде зависимости измеряемого свойства (поглощение в УФ области и т.п.) от объема элюата, вышедшего из колонки. Пики на таком графике соответствуют выходу индивидуальных белков. Если при разделении получается только один пик, это указывает на чистоту исходного препарата, нанесенного на колонку.
Свойство белков приобретать определенной величины заряд при данном значении рН раствора нашло широкое применение для их разделения методом электрофореза. Электрофорез основан на передвижении заряженной частицы в электрическом поле. Движение ее происходит в жидкой среде, которая удерживается инертным твердым носителем, например, полоской бумаги, гелевой пленкой из крахмала, полиакриламида, декстрона и т.д.
При постоянном напряжении движение заряженной молекулы белка определяется отношением заряда к ее размеру:
m =¦ (Q/r)
где Q-суммарный заряд белковой молекулы;
r-радиус молекулы.
С увеличением этого отношения подвижность молекулы растет. Так как каждый белок имеет свою определенную величину Q/r, скорость перемещения различных белков в электрическом поле будет различной. Электрофорез используется для разделения белков и определения их молекулярных масс.
Применение в определенной последовательности выше перечисленных методов позволяет получить белок в очищенном состоянии, не лишенный, однако, некоторых примесей солей. Для полного очищения белков от низкомолекулярных примесей используются методы диализа, кристаллизации, гельхроматографии и ультрафильтрации.
Является ли полученный белковый препарат индивидуальным белком или смесью имеет важное значение. Всегда можно ожидать, что в составе изолированного белка есть примесь других белков; это может привести к неправильным выводам о свойствах исследуемого белка. Поэтому большое внимание уделяется оценке гомогенности – однородности белков. Критерием чистоты белков служат следующие показатели: получение белка в кристаллическом состоянии; дальнейшая неразделяемость при электрофорезе и ультрацентрифугировании; независимость растворимости от количества твердой фазы; постоянство аминокислотного состава; определенный молекулярный вес; для многих белков – постоянство специфических биологических свойств (ферментативная активность, гормональная активность и т.д.)
Белки в зависимости от химического строения делят на простые и сложные. Простые белки при гидролизе распадаются только на аминокислоты. При гидролизе сложных белков наряду с аминокислотами образуется вещество небелковой природы – простетическая группа. Классификация простых белков основана на их растворимости.
Альбумины – водорастворимые белки с высокой гидрофильностью, выпадают в осадок при 100%-ом насыщении сульфатом аммония. К этим белкам относятся белок куриного яйца, белки зародыша семян злаковых и бобовых культур. Альбумин пшеницы называют лейкозин, гороха – легумелин. Альбумины содержат все незаменимые аминокислоты.
Глобулины – растворяются в солевых растворах, чаще всего для извлечения глобулинов используют 2 –10%-ый раствор хлорида натрия. Они осаждаются 50%-ым раствором сульфата аммония. Белки семян бобовых и масличных культур в основном представлены глобулинами; легумин – гороха и чечевицы, фазеолин – фасоли; глицин – соевых бобов. Многие альбумины и глобулины обладают ферментативным действием.
Проламины. Эта группа белков характерна исключительно для семян злаков. Эти белки растворяются в 60-80%-ом растворе этилового спирта. Эти белки содержат значительные количества пролина и глютаминовой кислот. Лизина они не содержат или содержат его в следовых количествах. Хорошо изучены проламины пшеницы – глиадины, ячменя – гордеин, кукурузы – зеин. Проламины – это комплексы белков различающиеся по составу и молекулярной массе.
Глютелины находятся, как правило, с проламинами. Растворяются они в щелочах (чаще 0,2%-ым NaOH). Глютелины не однородные белки, а смеси разных белков со сходными свойствами. Наиболее исследованы глютелин пшеницы, орезенин риса.
Глютенин и глиадин пшеницы образуют комплекс, который называют клейковиной. Клейковина муки влияет на структурно-механические свойства теста, а, следовательно на качество хлеба.
Протамины – самые низкомолекулярные белки. Встречаются эти белки в молоках рыб. На 2/3 эти белки состоят из аргинина, поэтому имеют основной характер. Протамины не содержат серы.
Гистоны – содержаться в хромосомах клеточных ядер, они участвуют в стабилизации пространственной структуры ДНК. Гистоны на 20-30% состоят из основных аминокислот. Из растворов их осаждают аммиаком.
Протеноиды – подгруппа нерастворимых фибриллярных белков животного происхождения. К ним относятся фиброин – шелка, кератин – волос, рогов, перьев, сухожилий и связок. Характерная особенность протеноидов – высокое содержание в них серы. Эти белки не гидролизуются пищеварительными ферментами.
Протеиды – сложные белки, в которых белок связан с веществом небелковой природы – простетической группой. В зависимости от химической природы простетической группы их делят на липопротеины, гликопротеины, хромопротеины, нуклеопротеины, фосфопротеины, металлопротеины.
В липопротеинах простетическая группа представлена каким-либо липидом. Эти белки входят в состав клеточных мембран, участвуют в структурной организации миелиновых оболочек, нервной ткани, хлоропластов и т.д., а так же присутствуют в свободном состоянии.
Гликопротеины – в качестве простетической группы содержат углеводный компонент. Это могут быть глюкоза, монноза, N-ацетилглюкозамин, L-фукоза и т.д. Эти белки выполняют специфические функции: обеспечивают клеточную адгезию, молекулярное и клеточное узнавание, антигенную активность опухолевых клеток, оказывают защитное, гормональное, и антивирусное действие.
Типичные представители гликопротеинов – белки, входящие в состав слюны, а так же некоторых растительных слизей. Растительные гликопротеины называют еще лектинами. Некоторые лектины оказывают антипитательное действие – они нарушают процессы всасывания питательных веществ. Антипитательное действие лектинов фасоли твердо установлено.
Хромомпротеины – состоят из простого белка и связанного с ним окрашеного небелкового компонента. Среди хромопротеинов различают гемопротеины, содержащие в качестве простетической группы железо- или магнийпорфирины; флавопротеины – содержащие производные изоаллоксазина. Хромопротеины участвуют в таких функциональных процессах жизнедеятельности, как транспорт кислорода и углекислого газа, фотосинтез, окислительно-восстановительные реакции, свето- и цветовосприятие и т.д. К группе хромопротеинов относятся гемоглобин и его производные, хлорофилсодержащие белки, такие ферменты как каталаза и пероксидаза, сукцинатдегидрогеназа, все белки цитохромы.
Нуклеопротеины – одна из самых важных групп белков, состоящая из простых белков связанных с нуклеиновыми кислотами. Эти белки играют первостепенную роль в хранении и передаче генетической информации и биосинтезе белка и содержаться в основном в ядрах клеток. Дезоксирибонуклеопротеины содержат дезоксирибонуклеиновую кислоту (ДНК). Рибонуклеопротеины содержат рибонуклеиновую кислоту (РНК)
Фосфопротеины – эти белки содержат органически связанный, лабильный фосфат, абсолютно необходимый для выполнения клеткой ряда биологических функций. Кроме того, они являются ценным источником энергетического и пластического материала в процессе роста и развития зародышей и молодого растущего организма. Наиболее изучены фосфопротеины – казеин молока, вителлин яичного желтка, ихтулин икры рыб. Металлопротеины наряду с белком содержат ионы какого-либо металла или нескольких металлов. Металлопротеины выполняют различные функции. Например, белок трансферрин (содержит железо) служит физиологическим переносчиком железа в организме. Другие металлопротеины являются биологическими катализаторами-ферментами – амилазы (содержат Са2+
) гидролизуют крахмал, карбоангидроза (Zn2+
) расщепляет угольную кислоту, аскорбинотоксидаза (Cu2+
) разрушает витамин С и т.д.
2. НУКЛЕИНОВЫЕ КИСЛОТЫ
Нуклеиновые кислоты были открыты в 1868г. швейцарским врачом Ф. Мишером. Биологическая функция этого вещества оставалась неизвестной еще в течение почти столетия, и только в 40-х годах прошлого века Эвери, Маклеод и Маккарти установили, что нуклеиновые кислоты, отвечают за хранение, репликацию (воспроизведение), транскрипцию (передачу) и трансляцию (воспроизведение на белок) генетической (наследственной) информации. Короче, именно нуклеиновые кислоты определяют вид, форму, химический состав и функции живой клетки и всего организма в целом.
В 1953 г. Уотсон и Крик сообщили о расшифровке молекулярной структуры ДНК. В каждом живом организме присутствуют два типа нуклеиновых кислот: рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК). В то же время вирусы содержат только один какой-нибудь тип нуклеиновых кислот: либо РНК, либо ДНК.
Нуклеиновые кислоты – это высокомолекулярные соединения, размер которых сильно варьирует. Молярная масса транспортной РНК составляет 25000, тогда как отдельные молекулы ДНК обладают массой от 1 000 000 до 1 000 000 000.
Количественное содержание ДНК в клетках одного и того же организма постоянно и исчисляется несколькими пикограммами, однако в клетках разных видов живых организмов имеются существенные количественные различия в содержании ДНК. ДНК преимущественно сосредоточено в ядре, митохондриях и хлоропластах. РНК большей частью содержится в цитоплазме клеток. Содержание РНК, как правило, в 5-10 раз больше, чем ДНК. Соотношение РНК/ДНК в клетках тем выше, чем интенсивнее в них синтез белка.
Нуклеиновые кислоты обладают сильно выраженными кислотными свойствами и при физиологических значениях рН несут высокий отрицательный заряд. В связи с этим в клетках организмов они легко взаимодействуют с различными катионами и прежде всего с основными белками, образуя нуклеопротеины.
Нуклеиновые кислоты при полном их гидролизе распадаются на три типа веществ – азотистые основания (пуриновые и пиримидиновые основания), сахара (пентозы) и фосфорную кислоту.
Пентозы нуклеиновых кислот представлены D-рибозой или 2-D-дезоксирибозой. Оба эти сахара содержатся в составе нуклеиновых кислот в фуранозной форме и имеют b-конфигурацию:
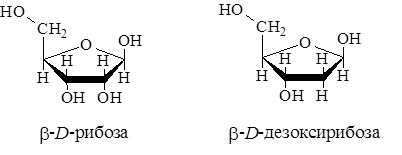
Нуклеиновая кислота называется рибонуклеиновой (РНК), если в ее состав входит рибоза, или дезоксирибонуклеиновой (ДНК), если в ее состав входит дезоксирибоза. Недавно установлено, что рибоза и дезоксирибоза не являются единственными углеводами, входящими в состав нуклеиновых кислот: в ряде фаговых ДНК и РНК некоторых видов раковых клеток найдена глюкоза.
Азотистые основания, которые обычно встречаются в нуклеиновых кислотах – это производные пурина аденин ( А) и гуанин (G)-и производные пиримидина – цитозин (С), тимин (Т) и урацил (U). Сами пурин и пиримидин в состав нуклеиновых кислот не входят.

Строение основных азотистых оснований-компонентов нуклеиновых кислот:
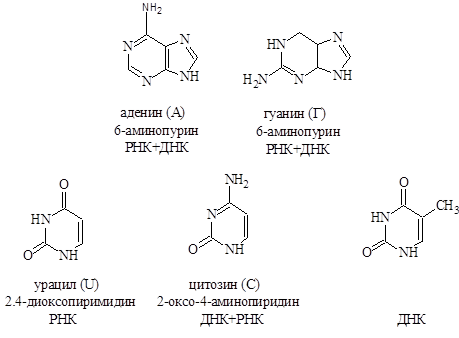
Цитозин, аденин, гуанин содержатся в нуклеиновых кислотах обоих типов, урацил входит только в состав РНК, а тимин в ДНК.
Для гуанина, цитозина, тимина и урацила известна кето-енольная таутомерия, однако кетоструктуры гораздо более стабильны и доминируют при физиологических условиях.
Таутомерия

В нуклеиновых кислотах все оксосодержащие азотистые основания присутствуют в кетоформе.
В составе ДНК и РНК встречаются так называемые необычные или «минорные» азотистые основания. К ним относятся, например, 5-метилцитозин, 4-тиоурацил, дигидроурацил и др.
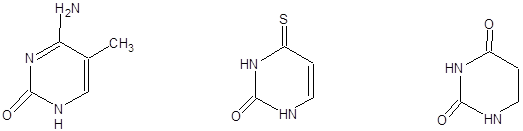
5- метилцитозин - тиоурацил дигидроурацил
( в ДНК) ( в тРНК) (в тРНК)
Рассмотренные пуриновые и пиримидиновые основания, а так же некоторые другие производные пурина и пиримидина, которые не входят в состав нуклеиновых кислот, часто содержатся в растениях в значительном количестве в свободном состоянии. Наиболее часто в свободном состоянии в растениях встречаются гипоксантин (6-гидроксиоксипурин), найденный в семенах горчицы и люпина. Ксантин (2,6-дигидроксиоксипурин) и аллонтоин очень широко распространены в растениях. В форме этих оснований, как и в форме амидов аминокислот, происходит запасание и транспорт азота в растениях.
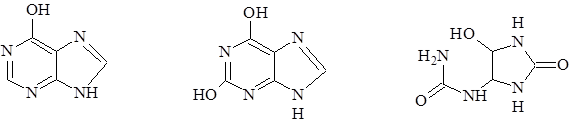
гипоксантин ксантин аллантоин
Пурины и пиримидины поглощают электромагнитную энергию в ультрафиолетовом (УФ) диапазоне, причем каждое соединение имеет характеристический спектр поглощения, однако для всех этих соединений максимум поглощения наблюдается вблизи 260 нм. Нуклеиновые кислоты так же поглощают в УФ-области. На этом свойстве основаны методы количественного определения нуклеиновых кислот.
В процессе обмена веществ у животных и растений пуриновые основания образуют такие продукты, как мочевая кислота, кофеин, теобромин, последние используются как лекарства.
Азотистое основание с присоединенным к нему углеводным остатком, называют нуклеозидом. В нуклеозидах ковалентная связь образована С1
-атом сахара и N1
- атомом пиримидина или N9
- атомом пурина, такая связь называется гликозидной. Что бы избежать путаницы в нумерации, атомы углеводной части отличают штрихом. Для наиболее распространенных нуклеозидов приняты тривиальные названия: аденозин, гуанозин, уридин и цитидин. Дезоксирибонуклеозиды называются дезоксиаденозин, дезоксигуанозин, дезоксицитидин и тимидин.
Например:
Пиримидиновый Пуриновый
рибонуклеозид дезоксирибонуклеозид

Нуклеозиды являются фрагментом структуры нуклеотидов; однако многие нуклеозиды встречаются в свободном состоянии. Некоторые из них обладают лечебными свойствами. Различные микроорганизмы выделяют арабинозилцитозин и арабинозиладенин, в состав которых входит b-D-арабиноза вместо рибозы. Эти вещества используются в качестве мощных антивирусных и антигрибковых агентов и против некоторых видов рака. Механизм действия ara-А и ara-С основан на ингибирование биосинтеза ДНК.
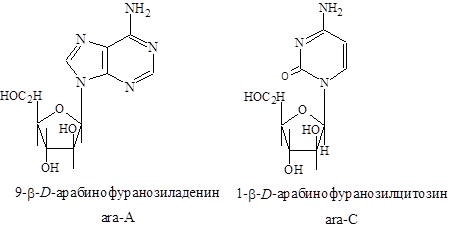
2.3Нуклеотиды
Нуклеотиды – это фосфорные эфиры нуклеозидов. В образовании связи участвует 51
-углеродный атом пентозы. В зависимости от строения пентозы все нуклеотиды можно разделить на рибонуклеотиды и дезоксирибонуклеотиды.
В зависимости от числа имеющихся остатков фосфорной кислоты различают нуклеозидмонофосфаты, нуклеозиддифосфаты и нуклеозидтрифосфаты. Все эти три вида нуклеотидов постоянно присутствуют в клетках.
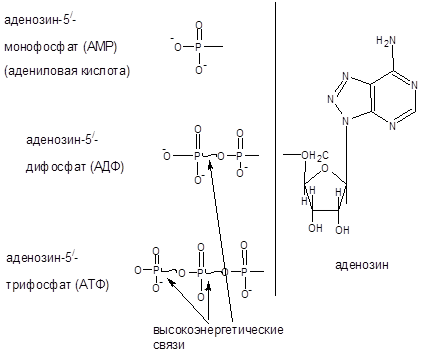
Рисунок 3 – моно-, ди- и трифосфонуклеотиды (51
) аденозина.
Названия отдельных нуклеотидов часто обозначают сокращенно большими первыми буквами названий соответствующих оснований. Ниже приведены нуклеотиды, входящие в состав нуклеиновых кислот, и даны их условные сокращенные обозначения.
Таблица 2 – Сокращенные названия отдельных нуклеотидов
| Основание |
Нуклеотиды в
составе РНК
|
Нуклеотиды в
составе ДНК
|
Сокращенное
обозначение
|
Аденин
Гуанин
Цитозин
Урацил
Тимин
|
Адениловая кислота
Гуаниловая кислота
Цитидиловая кислота
Урациловая кислота
Тиминовая кислота
|
Дезоксиадениловая кислота
Дезоксигуаниловая кислота
Дезоксицитидиловая кислота
Дезоксиуридиловая кислота
Дезокситимидиловая кислота
|
А
G
С
U
Т
|
Нуклеотиды – это сильные кислоты, так как остаток фосфорной кислоты, входящий в их состав, сильно ионизирован.
Главная функция нуклеотидов в клетке состоит в том, что они являются составными частями нуклеиновых кислот.
Все нуклеозиддифосфаты и нуклеозидтрифосфаты содержат высокоэнергетические связи (обозначены значком «»»). При гидролизе этой связи освобождается от 30 до 50 кДж/моль энергии, в то время как при гидролизе обычной сложноэфирной фосфатной связи освобождается энергия равная 8-12 кДж/моль.
Под влиянием соответствующих ферментов фосфатные группы содержащие высокоэнергетические связи, могут быть перенесены на другие вещества. Таким образом энергия, накопившаяся в высокоэнергетических соединениях, может быть использована далее в обмене веществ. Например: АДФ и АТФ принимают участие в биосинтезе белка. Уридинтрифосфат (UТФ) и уридиндифосфат (UДФ) необходимы для действия ферментов, катализирующих превращения и синтез сахаров (СДФ и СТФ) цитидиндифосфат и цитидинтрифосфат принимают участие в биосинтезе фосфолипидов.
Циклические нуклеотиды были выделены в 1959г. Сазерлендом ( лауреат Нобелевской премии 1971г.) при изучении механизма действия некоторых гормонов при регулировании метаболизма углеводов. В циклических нуклеотидах фосфорная кислота связывает два атома кислорода пентозного остатка в одном и том же нуклеотиде. Известны три циклических нуклеотида – циклический аденозинмонофосфат (с АМФ), циклический гуанозинмонофосфат (с GМФ) и циклический цитозинмонофосфат ( с СМФ).
Эти нуклеотиды образуются из соответствующих нуклеозидтри-фосфатов под действием ферментов аденилатциклазы и гуанилатциклазы. В биологических процессах они выступают в качестве промежуточного посредственника регуляторного действия гормонов.
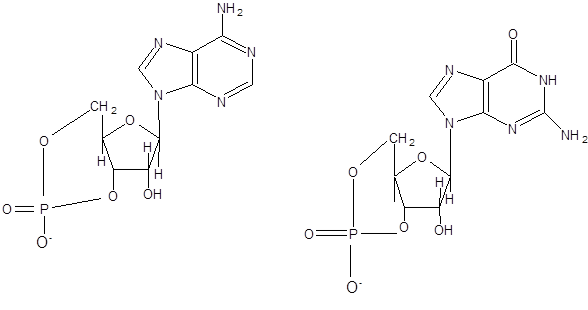
аденозин-3/
,5/
-цикломонофосфат гуанозин-3/
,5/
-цикломонофосфат
(3/
,5/
- сАМФ) (3/
,5/
-сGМФ)
Фосфатные остатки могут образовывать друг с другом кислотные ангидриды. Поэтому у нуклеотидов имеется возможность связываться друг с другом через фосфатные остатки. При этом возникают динуклеотиды с фосфоангидридной структурой.
К этой группе относятся некоторые кофакторы ферментов – НАД+
и НАДФ+
( никотинамидадениндинуклеотид ), ФАД ( флавинадениндинуклео-тидфосфат) и т. д.
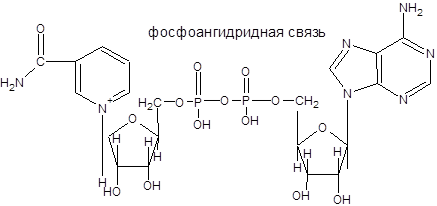
НАД+
Нуклеиновые кислоты это полимеры, состоящие из нуклеозидмонофосфатов, соединенных фосфодиэфирными связями. Поскольку фосфатная группа участвует в образовании двух эфирных связей с участием 3/
-и 5/
-углеродных атомов сахарных остатков двух соседних нуклеотидов эту связь называют 3/
- и 5/
-фосфодиэфирной связью.
Ниже изображены короткие структурные фрагменты цепей РНК и ДНК, позволяющие представить соединение отдельных нуклеотидов в цепи.

Рис. фрагмент РНК фрагмент ДНК
У полинуклеотида имеется 5/
-конец со свободной фосфатной группой и 3/
-конец со свободной ОН-группой. Фосфатные группы в этих цепях обладают сильнокислотными свойствами. При рН~7 фосфатная группа ионизирована полностью, поэтому в естественных условиях нуклеиновые кислоты существуют в виде полианинов (несут множество отрицательных зарядов).
Нуклеиновые кислоты отличаются друг от друга числом мононуклеотидных остатков в молекуле, нуклеотидным составом и порядком чередования нуклеитидных остатков, фактически оснований, поскольку пентозофосфатные части у всех мономеров одинаковы. Для краткого изображения первичной структуры нуклеиновых кислот пользуются однобуквенными символами нуклеозидов.
Поэтому первичная структура фрагмента РНК может быть представлена такой записью UAГААСС××××××××Запись структуры ДНК отличается приставкой « g» ( дезокси-);
g (ТСАГТГ –)-эти две записи, помимо символа «g» различаются еще тем, что в первой (РНК) не встречается символ Т ( тимин), а во второй (ДНК) не встречается символ U (урацил).
Таким образом, полинуклеотид записывается как последовательный набор конкретных нуклеотидных остатков от 5/
-конца к 3/
-концу.
Нуклеотидный состав ДНК (независимо от источников ее выделения) имеет общие закономерности, которые известны как правила Чаргаффа (по имени ученого, сформулировавшего эти правила).
1. Число пуриновых оснований (А+G) равно числу пиримидиновых оснований ( Т+С), т. е. отношение пуринов к пиримидинам равно единице.
2. Число остатков аденина равно числу остатков тимина, т.е. отношение аденина к тимину равно единице (А/Т = 1,0)
Эти количественные соотношения были подтверждены исследованиями других ученых и стали важной предпосылкой при установлении трехмерной структуры ДНК и помогли понять каким образом генетическая информация кодируется в ДНК и передается от одного поколения к другому.
Базируясь на данных рентгеноструктурного анализа и правилах Чаргаффа Дж. Уотсон и Ф. Крик в 1953г. предложили следующую модель строения ДНК. Согласно этой модели, молекула ДНК состоит из двух полинуклеотидных антипараллельных цепей (5/
®3/
)(3/
®5/
) спирально право-закрученных одна относительно другой таким образом, что углеводнофосфатная цепь находится снаружи, а пуриновые и пиримидиновые основания внутри перпендикулярно центральной оси схема ДНК. Эти две цепи соединяются между собой водородными связями, возникающими между пуриновыми и пиримидиновыми основаниями отдельных нуклеотидов, образуя специфические пары.
Тимин связан тремя водородными связями с аденином ТºА, цитозин двумя водородными связями с гуанином G= С. Эти пары оснований называются комплементарными парами оснований. Благодаря этому нуклеотидная последовательность одной цепи полностью комплементарна последовательности другой.
Парные основания могут охватывать миллионы оснований в ДНК. Это возможно только тогда, когда полярность обоих нитей различна, т.е, когда нити имеют различное направление (различную ориентацию). Кроме того, обе нити должны быть скручены друг вокруг друга в виде двойной спирали. РНК не может образовывать из-за стерических помех, благодаря 2/
- ОН групп рибозных остатков, подобную двойную спираль. Поэтому в РНК попарное соединение азотистых оснований находят только в пределах коротких участков одной и той же нити, и структура в целом менее регулярна, чем для ДНК.

Рисунок 4 – Схема образования водородных связей между комплементарными азотистыми основаниями

Рисунок 5 – Схематическое изображение двойной спирали ДНК
Водородные связи между парами оснований – не единственный вид взаимодействий, стабилизирующих двухцепочечную структуру. Молекула ДНК – полианион, и на ее поверхности локализовано множество отрицательных зарядов, что обеспечивает стабилизацию путем электростатических взаимодействий с неорганическими противоионами, например с Mg+2
,или белками, содержащими большое количество положительно заряженных боковых цепей аминокислот – гистонами. Третий стабилизирующий фактор возникает благодаря гидрофобным взаимодействиям между азотистыми основаниями, которые уложены стопкой внутри спирали. Между нитями по всей длине ДНК лежат углубления – маленькая и большая бороздки.
Так как обе нити удерживаются вместе благодаря нековалентным взаимодействиям, то двойную спираль можно разделить нагреванием (денатурацией) на одиночные нити (рисунок 5). При медленном охлаждении структура двойной спирали снова восстанавливается. Денатурация ДНК играет важную роль в генной инженерии. В зависимости от рН среды, ионной силы раствора, концентрации воды и т.п. конфигурация двойной спирали может меняться. Методами рентгеноструктурного анализа доказано существование более десяти форм ДНК, которые различаются количеством пар оснований приходящихся на один виток, углом наклона оснований к вертикальной оси. Наиболее изучены А-, В-, С- и Т-формы ДНК. Предполагают, что каждая форма ДНК приспособлена для выполнения определенной биологической функции. А-форма ДНК с передачей информации от ДНК к РНК, В-форма – с биосинтезом ДНК и С-форма с хранением, упаковкой ДНК.

Рисунок 6 – разделение двойной спирали ДНК на одиночные нити
В последние годы появились данные о возможности существования левозакрученной биспиральной молекулы ДНК-Z-формы и SBS формы ДНК, у которой полидезоксирибонуклеотидные цепи располагаются бок о бок (лесенкой, без закручивания). Такая форма ДНК обеспечивает легкое распаривание и расхождение цепей ДНК, что очень важно при биосинтезе ДНК.
ДНК обладает специфической третичной структурой. Двухцепочечная спираль ДНК на отдльных участках может подвергаться дальнейшей укладке в суперспираль. Может приобретать кольцевую форму, или свертываться в клубок. Суперскрученная структура обеспечивает экономную упаковку огромной молекулы ДНК в хромосоме. Суперспирали соединяются с белками (гистонами), упакованными в бороздах, обеспечивая тем самым стабильность третичной структуры ДНК.
В клетках любых биологических объектов содержаться три основных вида РНК: рибосомальная РНК(рРНК), транспортная РНК(тРНК) и информационная или матричная РНК(мРНК). Они являются одноцепочечными молекулами различной длины, различаются по локализации, свойствам, строении., функциям. В большинстве клеток содержание РНК в 5-10 раз превышает содержание ДНК. Основная часть РНК клетки–70-80% приходится на долю рРНК, которая содержится в рибосомах-внутриклеточных частицах, принимающих участие в биосинтезе белка рРНК образует каркас, к которому прикрепляются белки, образуя плотноупакованный рибонуклеопротеин. Нуклеотидный состав рРНК из разных источников сходен.
Существование матричной или информативной РНК (РНК-посредника передачи информации от ДНК в белоксинтезирующий аппарат клетки) было предсказано в 1957г., а выделена мРНК в 1962г. Содержание матричной РНК в клетке от 3% до 7% от общей суммы содержания РНК. Строение матричной РНК несколько специфично. В ее составе есть информативные участки, т.е. работающие как матрицы в процессе биосинтеза белка и неинформативные зоны. Предполагается, что неинформативные участки являются акцепторными при взаимодействии матричной РНК с рибосомой или отдельными белковыми факторами.
На 5/
-конце молекулы РНК имеется участок, содержащий минорные нуклеотиды. Это часть так же неинформативна и называется «шапочка» или «кэп». Предполагают, что «кэп» защищает мРНК от разрушительного действия ферментов экзонуклеаз. На 3/
-конце мРНК находится участок, содержащий от 50 до 400 остатков аденозинмонофосфата. Предполагают, что полиадениловый участок определяет время жизни мРНК, а так же участвует в процессе созревания и переноса м РНК из ядра в цитоплазму.
Название матричной РНК связано с функцией, которую она выполняет. Она служит матрицей, на которой синтезируется полипептидная цепь в рибосоме. Так же ее называют информационной так как она содержит информацию о том, какие аминокислоты и в какой последовательности, располагаются в белке. Эта информация представляет тройной нуклеотид, который называется кодоном. Каждый последовательно присоединенный набор из трех нуклеотидов (кодон) обеспечивает информацию для последовательного (упорядоченного) присоединения аминокислот при биосинтезе полипептида. Например, АИА-изолейцина, GAU-аспарагиновой кислоты и т.д. Последовательность UUUAUAGAU (читается по три нуклеотида UUU-AUA-GAU) определяет фрагмент трипептида фен-им-асп. В клетках синтезируются тысячи различных белков, поэтому существуют тысячи матричных РНК по структуре своей комплементарных отдельным участкам ДНК.
Содержание транспортной РНК 10% от общего содержания РНК, это самые малые по размеру молекулы РНК. Транспортная РНК не связана с клеточными структурами и находится в клетке в растворенном виде. Ее функция состоит в переносе, транспорте аминокислот к месту белкового синтеза – в рибосомы. Каждая тРНК переносит определенную аминокислоту. тРНК богаты минорными нуклеотидами.
Молекулы РНК, в отличие от молекул ДНК построены из одной нуклеотидной цепи, однако в этой цепи имеются комплементарные друг другу участки, которые могут взаимодействовать, образуя двойные спирали. При этом соединяются следующие нуклеотидные пары: Аденин-Урацил, Гуанин-Цитозин. Такие спирализованные участки (шпильки) обычно содержат небольшое число нуклеотидных пар и чередуются с неспаренными участками. Характерную вторичную структуру имеет транспортная РНК. Она имеет 4 спирализованные участка, и на плоскости эта структура напоминает фигуру клеверного листа. Кроме этого, имеется участок, содержащий нуклеотид комплементарный кодону матричной РНК, он называется антикодоном. С его помощью транспортная РНК прикрепляется к кодону матричной РНК. Имеется конец, который содержит остаток аденозинмонофосфата, к которому присоединяется соответствующая аминокислота. Третичная структура всех транспортных РНК схожа. Это позволяет всем им взаимодействовать с рибосомой. Незначительные отличия в пространственной структуре позволяет им взаимодействовать со специфическими ферментами тРНК синтезами, участвующими в биосинтезе белка. Третичная структура других видов РНК пока точно не установлена.
1. Березов Т.Т., Коровкин Б.Ф. Биологическая химия.-М.: Медицина, 1998.-704 с.
2. Филиппович Ю.Б. Основы биохимии.-М.: ИзD-во «Ангар», 1999–512 с.
3. Жеребцов Н.А., Попова Т.Н., Артюхов В.Г. Биохимия-Воронеж; Издательство ВГУ, 2002-696 с.
4. Биохимия растительного сырья. /Под. ред. В.Г. Щербакова.-М.: Колос, 1999.-376 с.
5. Ленинджер А. Биохимия.-М.: Мир, 1999 Т 1-3.
6. Основы биохимии / Под ред. А.А. Анисимова.-М.: Высшая школа, 1986.-551 с.
7. Кретович В.Л. Биохимия растений М.: Высш. школа, 1986.-503 с.
8. Химия и биохимия аминокислот и полипептидов. Методические указания для студентов технологических специальностей 49 01 01, 49 01 02, 91 01 01,48 01 02 по дисциплинам «Органическая химия», «Биологческая химия» и «Химия природных волокнообразующих полимеров». МГУП, Могилев, 2003 – 43 с.
|