Міністерство освіти і науки України Національний технічний університет України «Київський політехнічний інститут»
Факультет біотехнології і біотехніки
Реферат по курсу: Біологія клітини
на тему: Хромосоми як матеріальна основа спадковості. Зміни хромосом та механізми їх реорганізації
Виконала:
студентка I курсу групи БТ-91
Терещук Ганна
Перевірила:
Жолнер Л.Г.
Київ 2009
Вступ
Успадковані ознаки закладені в матеріальних одиницях, генах, які розташовуються в хромосомах клітинного ядра. Хімічна природа генів відома з 1944 г.: йдеться про дезоксирибонуклеїнову кислоту (ДНК). Фізична структура була з'ясована в 1953 р. Подвійна спіраль цієї макромолекули пояснює механізм спадкової передачі ознак.
Придивляючись до навколишнього нас світу, ми відзначаємо велику різноманітність живих істот – від рослин до тварин. Різноманітністю, що під цим здається, насправді ховається дивна єдність живих клітин – елементів, з яких зібраний будь-який організм і взаємодією яких визначається його гармонійне існування. З позиції вигляду схожість між окремими особинами велика, і все-таки не існує двох абсолютно ідентичних організмів (не рахуючи однояйцевих близнят).Наприкінці XIX століття в роботах Грегора Менделя були сформульовані основні закони, що визначили спадкову передачу ознак з покоління в покоління. На початку ХХ століття в дослідах Т.Моргана було показано, що елементарні успадковані ознаки обумовлені матеріальними одиницями (генами), локалізованими в хромосомах, де вони розташовуються послідовно один за одним.
У 1944 р. роботи Евері, Мак-Леода і Мак-Карті визначили хімічну природу генів: вони складаються з дезоксирибонуклеїнової кислоти (ДНК). Через 10 років Дж. Уотсон і Ф. Крік запропонували модель фізичної структури молекули ДНК. Довга молекула утворена подвійною спіраллю, а компліментарна взаємодія між двома нитками цієї спіралі дозволяє зрозуміти, яким чином генетична інформація точно копіюється (репліцируєтся) і передається подальшим поколінням.
Одночасно з цими відкриттями вчені намагалися проаналізувати і «продукти» генів, тобто ті молекули, які синтезуються в клітках під їх контролем. Роботи Ефрусси, Бідла і Татума напередодні другої світової війни висунули ідею про те, що гени «продукують» білки. Отже, ген зберігає інформацію для синтезу білка (ферменту), необхідного для успішного здійснення в клітці певної реакції. Але довелося почекати до 60-х років, перш ніж був розгаданий складний механізм розшифровки інформації, увязненій в ДНК, і її переведення у форму білка. Врешті-решт, багато в чому завдяки працям Ніренберга (США), був відкритий закон відповідності між ДНК і білками – генетичний код.
1.
Структура ДНК
Ще в 1869 році швейцарський біохімік Фрідріх Мішер виявив в ядрі кліток з'єднання з кислотними властивостями і з ще більшою молекулярною масою, ніж білки. Альтман назвав їх нуклеїновими кислотами, від латинського слова «нуклеус» - ядро. Так само, як і білки, нуклеїнові кислоти є полімерами. Мономерами їх служать нуклеотиди, у зв'язку з чим нуклеїнові кислоти можна ще назвати полінуклеотидами.
Нуклеїнові кислоти були знайдені в клітинах всіх організмів, починаючи від простих і закінчуючи вищими. Найдивніше, що хімічний склад, структура і основні властивості цих речовин виявилися схожими у всіляких живих організмів. Але якщо в побудові білків беруть участь близько 20 видів амінокислот, то різних нуклеотидів, що входять до складу нуклеїнових кислот, всього чотири.
У живих клітинах міститься два типи нуклеїнових кислот – дезоксирибонуклеїнова (ДНК) і рібонуклеїнова (РНК). Як ДНК, так і РНК несуть в собі нуклеотиди, що складаються з трьох компонентів: азотистої підстави, вуглеводу, залишку фосфорної кислоти. Проте комбінація цих компонентів в ДНК і РНК декілька різні.
Фосфорна кислота в молекулах ДНК і РНК однакова. Вуглевод же є у двох варіантах: у нуклеотидів ДНК – дезоксирибоза, а в нуклеотидів РНК – рібоза. І рібоза, і дезоксирибоза – пятічленниє, п'ятивуглецеві з'єднання – пентози. В дезоксирибози, на відміну від рібози, лише на один атом кисню менше, що і визначає її назву, оскільки дезоксирибоза в перекладі з латинського означає позбавлена кисню рібоза. Строга локалізація дезоксирибози в ДНК, а рібози в РНК, якраз і визначає назву цих двох видів нуклеїнових кислот.
Третій компонент нуклеотидів ДНК і РНК – азотисті з'єднання, тобто речовини, що містять азот і що володіють лужними властивостями. У нуклеїнові кислоти входять дві групи азотистих основ. Одні з них відносяться до групи пірімідінів, основу будови яких складає шестичленне кільце, а інші до групи пуринів, в яких до пірімідінового кільця приєднано ще і пятічленне кільце.
До складу молекул ДНК і РНК входять два різні пурини і два різні піримідини. У ДНК є пурини – аденін, гуанін і пірімідіни – цитозін, тимін. У молекулах РНК ті ж самі пурини, але з пірімідінов – цитозін і замість тиміну – урацил. Залежно від вмісту тієї або іншої азотистої підстави нуклеотиди називаються аденіловими, тіміловимі, цитозіловимі, урациловимі, гуаніловимі.
Як же з'єднуються між собою нуклеотиди в довгі полінуклеотідниє ланцюги? Виявляється, що таке з'єднання здійснюється шляхом встановлення зв'язку між залишком молекули фосфорної кислоти одного нуклеотиду і вуглеводом іншого. Утворюється сахаро-фосфорній скелет молекули полінуклеотиду, до якого збоку один за іншим приєднуються азотисті основи.
Якщо врахувати, що в кожній нуклеїновій кислоті по чотири види азотистих основ, то можна уявити собі безліч способів розташування їх в ланцюзі, подібно до того, як можна в самій різній послідовності нанизати на нитку намистинки чотирьох кольорів – червоні, білі, жовті, зелені. Послідовність розташування нуклеотидів в ланцюгах молекул нуклеїнових кислот так само, як і амінокислот в молекулах білків, строго специфічна для кліток різних організмів, тобто носить видовий характер.
ДНК є подвійною спіраллю.
Полінуклєїдні ланцюги досягають гігантських розмірів. Цілком зрозуміло, що у зв'язку з цим вони так само, як і білки, певним чином упаковані в клітці.
Модель структури молекули ДНК вперше створили біохіміки з Кембріджського університету в Англії Джеймс Уотсон і Френсис Крик. Було показано, що молекула ДНК складається з двох полінуклеотідних ланцюгів, закручених одна довкола іншої, з утворенням подвійної спіралі. Причому контакти існують між обома полінуклеотіднимі ланцюгами, точніше, між пурином одного нуклеотиду і піримідином іншого. Отже зовні молекулу ДНК можна представити як свого роду перекручені мотузяні сходи.
Утворення зв'язків в молекулі ДНК – процес строго закономірний. Аденіловий нуклеотид може утворювати зв'язки лише з тіміловим, а гуаніловий лише з цитозіловим. Ця закономірність отримала назву принципу компліментарності, тобто додатковості. Насправді, така строга послідовність у виборі пари наводить на думку, що в подвійній молекулі ДНК аденін як би доповнює тимін і навпаки, а гуанін відповідно – цитозін, як дві половинки розбитого дзеркала.
Принцип компліментарності дозволяє зрозуміти механізм унікальної властивості молекул ДНК – їх здатність самовідтворюватися. ДНК – це єдина речовина в живих клітинах, що володіє подібною властивістю. Процес самовідтворення молекул ДНК відбувається при активній участі ферментів. Особливі розплітаючі білки послідовно як би проходять уздовж системи водневих зв'язків, що сполучають азотисті основи обох полінуклеотідних ланцюгів, і розривають їх. Одиночні полінуклеотідні ланцюги ДНК, що утворилися в результаті, добудовуються згідно з принципом компліментарності за допомогою ферменту за рахунок вільних нуклеотидів, що завжди знаходяться в цитоплазмі і ядрі. Навпроти гуанілового нуклеотиду стає вільний цитозіловий нуклеотид, а навпроти цитозілового, у свою чергу, гуаніловий і так далі. У ланцюзі, що знов утворився, виникають вуглеводно-фосфатні і водневі зв'язки. Таким чином, в ході самовідтворення ДНК з однієї молекули синтезуються дві нові.
ДНК в клітці локалізована в основному в ядрі, в його структурних компонентах – хромосомах.
2.
Хромосоми еукаріот
У 80-х роках минулого століття в ядрах еукаріотичних клітин були відкриті ниткоподібні структури (Ст Флеммінг, Е. Страсбургер, Е. Ван Бенеден), названі В. Вальдейером (1888 р.) хромосомами (від греч. chroma – колір, забарвлення, soma – тіло). Цим терміном була підкреслено сильна схожість хромосом в порівнянні з іншими клітинними органелами до основних фарбників. Протягом подальших 10 – 15 років більшістю біологів було підтверджено, що саме хромосоми служать матеріальним носієм спадковості.
Хромосоми особливо чітко видно під час ділень клітин, проте факт безперервності їх існування і в ядрах, що не діляться, сумнівів не викликає. Основна особливість функціональних перетворень хромосом полягає в циклі компактізациі – декомпактізациі. У компактізованном стані хромосомами є короткі товсті нитки, видимі в світловий мікроскоп. В результаті декомпактізациі хромосомна нитка стає невидимою в світловий мікроскоп, тому ядра багатьох живих клітин виглядають оптично пустими.
Перетворення хромосом строго залежать від фаз клітинного циклу, тому їх особливості можуть розглядатися лише стосовно тієї або іншої фази циклу. Проміжок часу між закінченням одного клітинного ділення – мітоза і закінченням подальшого називається мітотичним циклом. Таким чином, мітотичний цикл включає мітоз і проміжок між мітозами – інтерфазу. Інтерфаза складається з трьох періодів: центрального – фази синтезу ДНК (S), коли генетичний матеріал подвоюється, а також передсинтетичного (G1) і синтетичного (G2) для поста, після якого клітина вступає в мітоз (М). Після фази синтезу ДНК в G2-періоде і в мітозі, аж до анафази, в хромосомі виявляються дві нитки, звані сестринськими хроматидами.
Основний хімічний компонент хромосом – молекули ДНК. Вміст її в ядрах соматичних кліток в два рази більше, ніж в ядрах зрілих статевих кліток. Ці два типи клітин відрізняються один від одного і по числу хромосом. Число хромосом – п в соматичних клітках і кількість ДНК – з (від англ. content – вміст) в них позначають як диплоїдне (2п хромосом, 2с ДНК), а в зрілих статевих клітинах як гаплоїдне (п хромосом, з ДНК).
2.1
Мітоз
Мітоз, або непряме ділення, - основний спосіб розмноження еукаріотичних клітин, що обумовлює, зокрема, можливість збільшення їх біомаси, зростання і регенерацію. Мітоз складається з чотирьох фаз.

Перша – профаза – характеризується початком циклу компактизації хромосом, який продовжується протягом всієї цієї фази. Внаслідок цього хромосоми стають видимими під мікроскопом, причому вже в середній профазі мітоза вони представляються подвійними структурами – сестринськими хроматидами, закрученими одна довкола іншої. До кінця профази зникають ядерце і ядерна мембрана.
Друга –метафаза. Процес компактизації хромосом продовжується і веде до ще більшого укорочення їх довжини. Хромосоми вишиковуються по екватору клітки. Хроматиди сполучені між собою між собою в центромері, званою також первинною перетяжкою. З'являються нитки мітотичного веретена, які приєднуються до ценромерів. Кожна ценромера випробовує напругу, оскільки нитки веретена тягнуть її до протилежних полюсів.
Полюси клітки формуються спеціальними органелами – центросомами.
Третя – анафаза – починається з розриву ценромери, внаслідок чого сестринські хроматиди розходяться до різних полюсів клітки. З цієї миті кожна пара сестринських хроматид отримує назву дочірніх хромосом.
Четверта – телофаза. Хромосоми досягають полюсів клітки, з'являються ядерна мембрана, ядерце. Відбуваються декомпактізация хромосом і відновлення структури інтерфазного ядра. Закінчується мітоз діленням цитоплазми і в типових випадках – відновленням вихідної біомаси дочірніх клітин.
Біологічна роль мітоза полягає в забезпеченні ідентичною генетичною інформацією двох дочірніх кліток. Це досяжно лише завдяки циклу компактізациі – декомпактізациі, який і дозволяє розподілити спадкові молекули в мінімальному об'ємі мітотичних хромосом. Інакше, враховуючи розміри клітки (десятки або сотні кубічних мікрометрів) і довжину декомпактізованной хромосоми (сантиметри), кожне клітинне ділення супроводилося б хаотичним переплетенням хромосомного матеріалу.
У еволюції еукаріотичних клітин, мабуть, ця обставина і послужила причиною становлення настільки складного генетичного процесу, як мітоз.
2.2
Мейоз
Терміном «мейоз» позначають два наступних один за одним ділення, в результаті яких з диплоїдних кліток утворюються гаплоїдниє статеві клітини, – гамети. Якби запліднення відбувалося диплоїдними гаметамі, то плодоїдність нащадків в кожному наступному поколінні повинна була б зростати в геометричній прогресії. В той же час завдяки мейозу зрілі гамети завжди гаплоїдни, що дозволяє зберігати діплоїдность соматичних кліток виду. Можливість існування подібного до мейозу ділення при дозріванні гамет тварин і рослин була передбачена А. Вейсманом ще в 1887 р. Мейотичне ділення не еквівалентне мітозу. Обом мейотичним діленням передує лише одна фаза синтезу ДНК.
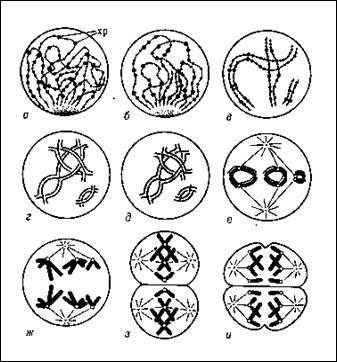
Тривалість її, як і профази I ділення мейозу, у багато разів перевершує відповідні показники мітотичного циклу будь-яких соматичних клітин даного виду. Головні події мейозу розгортаються в профазі I ділення. Вона складається з п'яти стадій.
У першій стадії – лептотені, слідуючою безпосередньо за закінченням предмейотичного синтезу ДНК, виявляються тонкі довгі хромосоми. Вони відрізняються в профазі мітоза двома особливостями: по-перше, в них не виявляється подвійність, тобто не видно сестринських хроматид, по-друге, лептотенні хромосоми мають виражену хромомерну будову. Хромомери – вузлики. Ділянки щільної компактізациі ДНК, розміри і розташування яких строго видоспецифічний. Хромомери зустрічаються як в мейотичних, так і в мітотичних хромосомах, проте в останніх без специфічної обробки вони не помітні.
Третя стадія профази I ділення – пахитена – у більшості видів найтриваліша. Під світловим мікроскопом видно кон’югіровавші хромосоми з більш менш чітко вираженою хромомерною будовою. Приблизно в середині пахитени між хроматидами гомологічних хромосом з'являється подовжня щілина, яка чітко показує, що бівалент – це, по суті, четверна хромосомна структура. У пахитене відбувається важлива генетична подія – кросинговер, або перехрещення хроматид гомологічних хромосом. В результаті цього в кожному гомологу змішуються батьківський і материнський спадковий матеріал.
Результати кросинговера стають помітними лише в четвертій і п'ятій стадіях профази I ділення – діплотені і діакенезі. Діплотена починається з моменту розбіжності гомологічних хромосом. В цей час в точках кросинговера видно перехрещені хроматиди. Область перехрещення хроматид називають хіазмою. Число хіазм в цілому відповідає кількості актів кросинговера в біваленті і пропорційно довжині гомологічних хромосом, його складових. Для діплотени і діакенезу характерне прогресуюче укорочення хромосом в результаті компактізаціі; тому хіазми поступово терміналізуются, тобто наближаються до кінців бівалента і спадають з нього. Таким чином, у міру наближення до метафази першого ділення число хіазм зменшується.
У метафазі I ділення мейозу район центромери кожної хромосоми сполучений (на відміну від метафази мітоза) ниткою веретена лише з одним полюсом клітки, причому центромери гомологів, що розійшлися, завжди пов'язані з протилежними полюсами.
Анафазе I ділення мейозу не передує розщеплювання центромери, як при мітозі, і тому до полюсів відходять не хроматиди, а цілі хромосоми, що складаються з двох хроматид. Проте, оскільки гомологічні хромосоми розходяться до різних полюсів, перше мейотичне ділення приводить до редукції числа хромосом. Іншими словами, по числу хромосом продукти I ділення мейозу стають гаплоїднимі. Проте у зв'язку з тим, що хромосоми в них зберігають подвійність, тобто містять дві хроматиди, кількість ДНК зменшується лише до 2с.
Друге ділення мейозу, наступне після короткого проміжку – інтеркінезу, приводить у відповідність число хромосом і вміст ДНК. Формально воно нагадує мітоз. На початку анафази відбувається розділення центромери, сестринські хроматиди стають дочірніми хромосомами і розходяться до полюсів. Таким чином, кожна з чотирьох клітин, що утворилися унаслідок двох мейотичних ділень однієї клітки, що пройшла предмейотичну s-фазу, міститиме п хромосом і з ДНК.
Отже, головна відмінність мейозу від мітоза – кон'югація гомологічних хромосом з подальшою розбіжністю їх в різні гамети. Точність розбіжності обумовлена точністю кон'югації, а остання – ідентичністю молекулярної структури ДНК гомологів.
2.3
Каріотип
Каріотипом називається хромосомний комплекс вигляду зі всіма його особливостями: числом хромосом, їх формою, наявністю видимих під світловим мікроскопом деталей будови окремих хромосом. Інколи термін «каріотип» вживають по відношенню до хромосомного набору одиничної клітини або групи тканинних клітин.

Число хромосом видоспецифічне. Хоча закономірності, що характеризують каріотип, інколи і відображають еволюцію певних видів, в цілому по структурі каріотипу прямо судити про систематичне положення виду не можна.
У більшості вищих тварин і рослин одна пара хромосом у особин однієї з родів гетероморфна. Ці несхожі хромосоми називаються статевими. Зокрема, у ссавців і в дрозофіли клітини чоловічих організмів мають Х- і y-хромосоми. В багатьох видів y-хромосома відсутня. Всі останні хромосоми називаються аутосомами.
Таким чином, завдяки дослідженням цитологів в кінці XIX – початку ХХ ст була обгрунтована роль ядра в спадковості, а спостереження за поведінкою хромосом в мітозе і мейозі привели до висновку, що саме з ними пов'язана передача спадкових ознак.
3.
Секрети генетичного коду
У організмі кожної людини – своя спадкова конституція, характерна лише для нього. Саме з цим пов'язана тканинна несумісність, що виявляється, зокрема, при пересадці органів і тканин від одного організму іншому. «Чужа» шкіра, наприклад, зі своїми особливими молекулами вступає в небажані реакції з організмом «господаря». Вона викликає появу білків – антитіл – і в результаті не «приживається». Аналогічне явище спостерігається і при пересадці окремих органів.
Інакше проходять ці процеси у однояйцевих близнят, які розвиваються з двох клітин, що утворилися з однієї заплідненої яйцеклітини – зіготи. Такі близнята завжди однополі і зовні вражаюче схожі один на одного. У однояйцевих близнят пересадка тканин і органів сповна можлива, жодного відторгнення їх не відбувається. Інакше і бути не може. Один і той же комплекс всіх спадкових чинників не провокує появи антитіл в їх організмах.
Ці і багато інших фактів показали, що програмування синтезу білків – головна властивість ДНК. Проте, перш ніж прийти до такого висновку, необхідно було довести, що саме ДНК – носій генетичної інформації. Перше підтвердження тому було отримано при вивченні явищ трансформації.
3.1 Історія доказу, що ДНК – носій генетичної інформації
Явище це було відкрите в дослідах з пневмококами, тобто з бактеріями, що викликають запалення легенів. Відомо дві форми пневмококів: А-форма з полісахарідной капсулою і б-форма без капсули. Обидві ці ознаки спадкові.
Пневмококи а-форми при зараженні ними мишей викликають запалення легенів, від якого миші гинуть. Б-форма для них нешкідлива.
У 1928 році англійський бактеріолог Ф.Гріффітс заражав мишей сумішшю, що складається з убитих нагріванням пневмококів а-форми і живих пневмококів Б- форми. Учений передбачав, що миші не захворіють. Але всупереч чеканням піддослідні тварини загинули. Ф. Гріффітсу удалося виділити з тканин загиблих мишей пневмококи. Всі вони виявилися капсулірованнимі, тобто А- форми. Отже, убита форма якимсь чином передавала свої властивості живим клітинам б-форми. Але як?
Від рішення цього питання залежало багато що, оскільки, встановивши речовину, що передає спадкову ознаку, – утворення капсули, можна було отримати потрібну відповідь. Проте зробити це не виходило досить довго. Лише через 16 років після дослідів Ф. Гріффітса, в 1944 році, американський учений А. Евері із співробітниками, поставивши ряд чітких експериментів, зумів з повним обгрунтуванням довести, що полісахарид і білок не мають жодного відношення до передачі спадкових властивостей пневмокока а-форми.
В процесі цих експериментів за допомогою спеціального ферменту розчинили полісахаридну капсулу убитих пневмококів а-форми і перевірили, чи продовжують залишки клітки форми А передавати спадкову інформацію клітинам форми Б. Виявилось, що продовжують. Зрозуміло, що полісахарид як джерело генетичної інформації відпадає.
Далі вчені за допомогою інших ферментів видалили із залишків пневмококів А білки і знову перевірили їх дію. Передача спадкової інформації від А до Б продовжувалася. Отже, і білок ні при чому.
Таким чином, методом виключення було встановлено, що спадкову інформацію в клітині зберігає і передає молекула ДНК. І дійсно, коли зруйнували ДНК, утворення капсульних форм А з беськапсульних Б припинилася.
Явище перетворення, тобто спадкової зміни властивостей однієї форми бактерій під впливом речовин іншої форми, було назване трансформацією. Речовина ж, що викликає трансформацію, отримала назву трансформуючого агента. Ним, як було встановлено, служить ДНК.
3.2 Розшифровка генетичної інформації
дезоксирибонуклеїновий хромосома генетичний спадковість
Полімерні ланцюги білків складаються з мономірних ланок – амінокислот і послідовність розташування їх в білковій молекулі строго специфічна. У зв'язку з цим очевидно, Що в ДНК повинна зберігатися інформація не лише про якісний і кількісний склад амінокислот в молекулі даного білка, але і про послідовність їх розташування. Відповідно якимсь чином мають бути закодовані в полінуклеотидному ланцюзі ДНК кожна амінокислота і білок в цілому.
Знаючи, що амінокислот всього 20, а нуклеотидів – 4, легко уявити собі, що 4 нуклеотида явно недостатньо для кодування 20 амінокислот. Недостатньо також і коди з двох нуклеотидів на кожну кислоту (4 = 16). Для кодування 20 амінокислот необхідно групи щонайменше з трьох нуклеотидів (4 = 64). Подібна група, що несе інформацію про одну амінокислоту в молекулі білка, називається кодоном. Вся ж ділянка ДНК, відповідальна за синтез однієї молекули білка, в цілому якраз і є ген. Значить, в гені стільки кодонів, скільки амінокислот входить до складу даного білка, що синтезується.
Синтез білків відбувається на рибосомах. ДНК же локалізована в ядрі, в його хромосомах. Виникає питання: яким чином генетична інформація з ядра переноситься в цитоплазму на рибосому? Передбачити, що ДНК сама поступає через пори ядерної мембрани, не можна: Адже ДНК ядер володіє величезною молекулярною масою і у зв'язку з цим просто не може проникнути через крихітні пори ядерної мембрани. Тому мають бути якісь дрібніші молекули – посередники, що передають генетичну інформацію від ДНК до білків. А.Н. Белозерський і А.Г. Спірін висунули міркування, що цю роль грають молекули РНК.
Але одразу ж виникає інше питання: як копіюється інформація з ДНК на коротші молекули РНК? Щоб відповісти на нього, треба пригадати, що в будові нуклеотиду ДНК і РНК багато загального. Зокрема, через схожість азотистих підстав інформація з ДНК на РНК може переноситися за принципом компліментарності, згідно з яким утворювати пари можуть не лише нуклеотиди в системі ДНК-ДНК, але і нуклеотиди в системі ДНК-РНК.
Оскільки РНК так само, як і ДНК, містить пурінові і пірімідінові основи, на ділянках одного з ланцюгів ДНК за допомогою ферменту РНК – полімерази будуються компліментарні короткі ланцюги РНК. Цей процес синтезу РНК на матриці ДНК, що відбувається за допомогою ферментів, носить назву транскрипції. В результаті процесу транскрипції закодована в ДНК послідовність нуклеотидів, яка і є певною генетичною інформацією, передається на РНК. Транскрипція відбувається на окремих ділянках ДНК – генах, кожен з яких містить набір кодонів, що програмують послідовності амінокислот в даній молекулі білка.
Рибонуклеїнова кислота, на якій зроблена копія ДНК, складається з одного ланцюга нуклеотидів, в яких дезоксирибоза замінена на рібозу, а тимін (Т) замінений на урацил (У) .
Таким чином, в кожному кодоні ДНК транскрибірується в компліментарний кодон РНК. В результаті виходить як би негатив РНК з позитиву – ДНК. Ця РНК, що знімає інформацію з ДНК, називається інформаційній РНК (І-РНК).
До теперішнього часу ученим удалося розшифрувати кодони для всіх амінокислот. Виявилось, що одній амінокислоті частенько відповідає декілька кодонів. Такий код називається виродженим. Поряд з цим виявилося, що деякі кодони не кодують жодну амінокислоту. Їх називають безглуздими. Безглузді кодони мають дуже важливе значення, оскільки визначають кордони початку і кінця транскрипції, тобто кордони генів в даній молекулі ДНК.
Якщо в прокаріот гени по своєму запису безперервні, то в еукаріот це далеко не так. Інформація необхідна для синтезу білка, виявляється записаною з пропусками, переривчасто: гени складені з кодуючих ділянок (екзонів), розділених некодуючими послідовностями (інтронами). При транскрипції таких генів інтрони копіюються разом з екзонамі в загальну молекулу пре-мРНК. Остання піддається в ядрі серії реакцій, в ході яких інтрони вирізуються, а екзони з'єднуються один з одним своїми краями. Молекула М-КОДУ-РНК, що вийшла, покидає ядро і виявляється вже у владі системи трансляції, що дешифрує нуклеотидну послідовність. З'єднання амінокислот з утворенням білка відбувається в цитоплазмі на особливих частках-рибосомах. Все це можна порівняти з фабрикою (клітина), в якій креслення (гени) зберігаються в бібліотеці (ядро), а для випуску продукції (білки) використовуються не самі креслення (ДНК), а їх фотокопія (мРНК). Копіювальна машина (РНК-полімераза) випускає або по одній сторіночці фотокопії (ген), або відразу цілу главу (оперон). Виготовлені копії видаються через спеціальні віконця (пори ядерної мембрани). Їх потім використовують на монтажних лініях (рибосоми) з дешифратором (генетичний код) для здобуття із заготовок (амінокислот) остаточної продукції (білки).
Як же відбувається сам процес синтезу білка?
Перший його етап пов'язаний з функціонуванням транспортної РНК (т- РНК). Число різновидів цих молекул РНК дорівнює числу основних амінокислот, тобто їх 20 видів. Кожній амінокислоті відповідає певна Т-РНК і певний фермент.
У цитоплазмі клітини завжди в достатній кількості є різні амінокислоти. З них молекула Т-РНК відбирає відповідну амінокислоту. Кожна амінокислота, перш ніж вступити в білковий ланцюг, за допомогою спеціального ферменту з'єднується з АТФ і запасається енергією.
«Зарядившись» таким чином амінокислота зв'язується з Т-РНК, яка переносить її до рибосом. Характерною межею молекул Т-РНК є наявність в їх структурах антикодонів. Ця особливість забезпечується розташуванням відповідних амінокислот в тій послідовності кодонів, яка зашифрована в молекулі І-РНК. Між рядом розташованими амінокислотами виникають пептидні зв'язки і синтезується молекула білка.
Таким чином, генетична інформація, ув’язнена в ДНК, реалізується різними видами РНК в молекулах відповідних білків. Процес передачі програми, принесеної з собою молекулами І-РНК, отримав назву трансляції.
4.
Як же працюють гени?
Біосинтез білків, що протікає під генетичним контролем, - це лише початок складних, багатоступінчастих біохімічних процесів клітки.
При вивченні рослин, що вегетативно розмножуються, отримані докази того, що окремі частини організму, такі як бульба, аркуш, цибулина, держак і так далі, дають початок нормальній рослині. А це означає, що всі клітини даного організму несуть повну генетичну інформацію, так само як і вихідна запліднена яйцеклітина, з якої розвивається тварина. В той же час в будь-якому організмі містяться диференційовані клітини з певною формою і функцією. Наприклад, у людини є клітини нервові, м’язові, статеві і так далі Але, не дивлячись на те, що кожна клітина нашого тіла несе повну генетичну інформацію, тобто повний набір генів, отриманих від батьків, функціонують лише певні гени, останні знаходяться в неактивному стані. Яким же чином в клітині регулюється діяльність тих або інших генів?
У всіх процесах життєдіяльності клітини роль біологічних каталізаторів грають ферменти. Без їх участі не протікає практично жодна хімічна реакція синтезу або розкладу речовин. У кожній клітині (з її характерними функціями) повинні знаходитися регуляторні механізми, контролюючі не лише якісний склад ферментів, але і їх кількість. Інакше макромолекули білків, що безперервно синтезуються, накопичувалися б в клітині непотрібним баластом, захаращуючи її.
І дійсно, подібний регуляторний механізм був виявлений в клітинах бактерій в 1961 році французькими ученими Франсуа Жакобом і Жаком Моно.
Що ж це за механізм?
Ф. Жакоб і Ж. Моно довели, що не всі гени бактерій однакові по своєму призначенню. Одна група – структурні гени, що видають інформацію про синтез певних поліпептидних ланцюгів, інша – регуляторні гени, що відають активністю структурних генів шляхом їх «включення» і «виключення».
Регуляторні гени представлені геном-оператором, безпосередньо зчепленим з групою структурних генів, і геном-регулювальником, який може знаходитися в деякому віддаленні від них.
Ген-оператор з групою регульованих їм структурних генів був названий опероном. Оперон служить одиницею транскрипції, тобто з нього списується одна молекула І-РНК. Ген регулювальник діє не шляхом безпосереднього контакту із структурними генами, а за допомогою білка репрессора.
За наявності молекул речовини, що синтезується, що досить накопичилися, белок-репрессор, з'єднуючись з цими молекулами, активізується і зв'язується з геном-оператором. В результаті синтез даної речовини припиняється. Свою назву белок-репрессор отримав через те, що пригнічує діяльність гена-оператора, тобто ставить його в положення «вимкнено».
При малій кількості молекул, що синтезуються, білок-репрессор залишається неактивним. У таких умовах дія оперону – гена-оператора і структурних генів – не пригнічується, і синтез продовжуватиметься безперешкодно.
5.
Передача генетичної інформації від батьків до нащадків
Як відомо, особливості, що характеризують нащадків, передаються їм від батьків через статеві клітини: чоловічу – сперматозоїд і жіночу – яйцеклітину. Злиття їх при заплідненні приводить до утворення єдиної клітини зіготи, з якої розвивається зародок людини. Очевидно, що саме в цих двох статевих клітинах і в зіготі, що утворилася при їх злитті, зберігається спадкова інформація про фізичні, біохімічні і фізіологічні властивості, з якими з'являється нова людина.
Матеріальною основою спадковості служать нуклеїнові кислоти, а саме ДНК. Але яким же чином генетична інформація передається від батьків до потомства? Як відомо, нові клітини з'являються в результаті ділення вихідних материнських.
Для більшості клітин характерне фізіологічно повноцінне клітинне ділення, що складається з ряду фаз, під час яких ядро зазнає закономірних змін, внаслідок чого утворюються два ядра, абсолютно ідентичні початковому. Цитоплазма при цьому ділиться на дві половини. Таке складне ділення отримала назва мітоза, і характерний воно для клітин тіла, тобто соматичних клітин.
Проте, в організмах рослин, тварин і людини, окрім соматичних, є і статеві клітини. Їх утворення відбувається в результаті особливого ділення. Перетворення ж, яке викликається цим діленням, отримало назву мейозу.
Під час і мітозу і мейозу ядро втрачає округлі контури і у ньому виразно вимальовувалися його структурні компоненти, звані хромосомами. Хромосоми мають різні форми: паличок, коротких стрижнів, капіж і так далі.
Висновок
Вивчення генетики людини, незважаючи на всю складність, важливе не лише з точки зору науки. Важко переоцінити і прикладне значення досліджень, що проводяться. Досягнення в цій області роблять помітний вплив на інші галузі наук про людину – медицину, психіатрію, психологію, педагогіку.
Зокрема, велика роль генетики людини, що розвивається, у вирішенні проблем спадкових хвороб. Сучасні дані свідчать, що людиною успадковуються багато хвороб, таких, як не згортання крові, колірна сліпота, ряд психічних захворювань. Крім того, генетика людини покликана вирішувати і інші питання.
Значення розвитку генетики людини очевидно. Можна з повною упевненістю сказати, що, наприклад, в молекулах ДНК кліток людини запрограмована генетична інформація, контролююча кожну мить нашого життя. Це стосується здоров'я, нормального розвитку, тривалості життя, спадкових хвороб, серцево-судинних захворювань, злоякісних пухлин, схильності до тих або інших інфе6кционним захворювань, старості і навіть смерті.
Якщо виділити з ядра однієї клітини людини всі генетичні молекули ДНК і розташувати їх в лінію одна за одною, то загальна довжина цієї лінії складе сім з половиною сантиметрів. Така біохімічна робоча поверхня хромосом. Це сконцентрована в молекулярному записі спадщина століть минулої еволюції.
Правильно і образно сказав про це свого часу в романі «Лезо бритви» письменник Іван Ефремов: «Спадкова пам'ять людського організму – результат життєвого досвіду незліченних поколінь, від риб'ячих наших предків до людини, від палеозойської ери до наших днів. Ця інстинктивна пам'ять клітин і організму в цілому є той автопілот, який автоматично веде нас через всі прояви життя, борючись з хворобами, заставляючи діяти складні автоматичні системи нервовою, хімічною, електричною і невідомо якого ще регулювання. Чим більше ми взнаємо біологію людини, тим більш складні системи ми в ній відкриваємо».
Дослідження останніх років довели, що будь-яка жива клітина, у тому числі і клітина людського організму, є цілісною системою, всі складові елементи якої виявляють тісну взаємодію між собою і довкіллям, що робить на гени величезний вплив. Тому розрізняють два поняття: генотип – комплекс всіх спадкових фактів – генів, що отримуються нащадками від батьків, і фенотип – сукупність ознак, що виникають при взаємодії генотипу і зовнішнього середовища.
Отже, у формуванні фенотипа важливі як генотип, так і зовнішнє середовище, в якому відбувається розвиток особини. Без цієї взаємодії неможливе було б життя, пов'язане з реалізацією генетичної інформації, ув’язненою в нуклеїнових кислотах.
Закономірності генетики в більшості випадків носять універсальний характер. Вони однаково важливі для рослин, для тварин. Велике їх значення і для людини.
Список літератури
1. Генетика і спадковість. Збірка статей. Г.34. Пер. з франц. М.: Світ. 1987 р.
2. Загальна генетика. Аліхонян С. І. І ін. М.: Вища школа.1985 р.
3. Генетика вчора, сьогодні і завтра. Дубінін Н.П. М.: Радянська Росія. 1981р.
4. Біологічний словник енцеклопедічеський.
5. Ботаніка: Морфологія і анатомія рослин. Васильєв А.Е. М.: Освіта. 1988 р.
|