Министерство образования Республики Беларусь
Учреждение образования
«Гомельский государственный университет
им. Ф. Скорины»
биологический факультет
Генетическая изменчивость, дифференциация и таксономические
взаимоотношения у лиственниц сибирской, Сукачёва и даурской
Курсовая работа
Исполнитель:
Студентка группы К-42 ____________
Лягушова А.Ю.
Научный руководитель:
Загрушевская Т.Е.
Гомель 2005
Содержание
Введение
1 Материалы и методы исследования
2 Результаты и их обсуждение
Заключение
литература
Введение
Лиственницы являются одними из главных структурных компонентов светлохвойной тайги. Площадь их лесов на постсоветском пространстве составляет 45% в структуре хвойных насаждений. Благодаря быстрому росту, высокой продуктивности (свыше 1000 м3
с га) (Пугач, 1985) лиственницы способны существенно повышать продуктивность лесов и поэтому широко внедряются в лесные культуры, в том числе и на территории республики Беларусь. Качество и продуктивность создаваемых лиственничных насаждений напрямую зависит от генофонда используемого семенного и посадочного материала. Поэтому, вопрос о том, генофонд какого вида наиболее успешно можно использовать для создания лиственничных культур, приобретает особую актуальность.
В настоящее время род Larix
объединяет более двадцати различных видов (Козубов, Муратова, 1986). Считается, что наибольше видовое разнообразие лиственниц сосредоточено в сибирско-дальневосточном регионе Палеарктики (Дылис, 1961; Бобров, 1978). Несмотря на то, что в последние годы в различных лабораториях были проведены интенсивные генетические исследования лиственниц Палеарктики с использованием изоферментов и фрагментов ДНК (Mejnartowicz, Bergmann, 1975; Paule, Gömöry 1995; Тимерьянов и др., 1986; Потенко, Разумов, 1996; Гончаренко, Силин, 1997; Lewandowski, 1997; Semerikovetal., 2003; Гончаренко,Шевцова, 2004; Ларионова и др. 2004; Козыренко и др. 2004), ряд вопросов касающихся уровня генетической изменчивости, дифференциации и генетико-таксономических взаимоотношений для видов этого региона не решены окончательно.
Целью нашей работы было на основании 20 изоферментных генов определить уровень генетической изменчивости и дифференциации трех видов – лиственницы сибирской (Larix sibirica
Ledeb.), лиственницы Сукачева (L.
s
ukaczewii
Dyl.)и лиственницы даурской (L
.
dahurica
Turcz.=L
.
gmelinii
Rupr.) – и уточнить их генетико-таксономический статус.
1. Материалы и методы исследования
При исследовании генетической изменчивости и таксономических взаимоотношений у трёх видов лиственниц Палеарктики нами был использован материал 160 деревьев из двух природных популяций лиственницы сибирской, одной популяций лиственницы даурской и одной популяции лиственницы Сукачева. Две исследованные природные популяции лиственницы сибирской расположены на территории Западной Сибири. Одна из них находится в смешанном горном лесном массиве Алтая, а другая в лиственничнике Западно-сибирской низменности, к северу от г. Томска. Проанализированная популяция так называемой лиственницы Сукачева представляет собой природное насаждение, расположенное на территории Центрального Урала. Семенной материал лиственницы даурской был собран с деревьев, произрастающих в лиственничнике недалеко от г. Хабаровска.
В качестве экспериментального материала при электрофоретическом фракционировании служили ткани гаплоидных эндоспермов и диплоидных зародышей. Для определения генотипа каждого дерева проводился анализ 10-20 эндоспермов, которые выбирались случайно из набора семян, полученного как минимум из пяти шишек, собранных из различных частей кроны дерева. Так как вероятность ошибочного отнесения гетерозиготных деревьев к гомозиготным рассчитывается из соотношения Р = 0.5 n–1
(где n – количество проанализированных эндоспермов), то даже при анализе 8 эндоспермов ошибка составляет менее 1%.
Электрофоретический анализ проводили в горизонтальных камерах в 13-14% крахмальном геле по методам, подробно описанным нами ранее (Гончаренко и др. 1989). Электрофорез проводили в трёх буферных системах: А – Трис - ЭДТА- боратная, рН 8,6; В – Трис-цитрат, рН 6,2 / Трис-НСl, рН 8,0; С – Трис-цитратная, рН 6,2. Название ферментов, кодовый номер согласно изданию “Номенклатура ферментов” (1979), предпочитаемая для анализа буферная система, а также количество используемых локусов приведены в табл. 1.
Таблица 1.
Ферменты, их кодовый номер, буферные системы и количество локусов, использованные для анализа популяций лиственницы сибирской, лиственницы Сукачева и лиственницы даурской.
| Фермент |
Аббревиатура |
Кодовый номер |
Буферная система |
Количество
локусов
|
| Аконитаза |
АСО |
4.2.1.3. |
C |
1 |
| Аспартатаминотрансфераза |
ААТ |
2.6.1.1. |
А |
3 |
| Глутаматдегидрогеназа |
GDH |
1.4.1.2. |
A |
1 |
| Глюкозофосфатизомераза |
GPI |
5.3.1.9. |
С |
1 |
| Изоцитратдегидрогеназа |
IDH |
1.1.1.42. |
B |
1 |
| Лейцинаминопептидаза |
LAP |
3.4.11.1. |
B |
2 |
| Малатдегидрогеназа |
MDH |
1.1.1.37. |
C |
4 |
| Фосфоглюкомутаза |
PGM |
2.7.5.1. |
A |
2 |
| Флюоресцентная эстераза |
FL-EST |
3.1.1.2. |
B |
1 |
| 6-фосфоглюконатдегидрогеназа |
PGD |
1.1.1.44. |
C |
2 |
| Сорбитолдегидрогеназа |
SDH |
1.1.1.14. |
A |
1 |
| Шикиматдегидрогеназа |
SKDH |
1.1.1.25. |
B |
1 |
Определение уровня генетической изменчивости проводили на основе ряда общепринятых показателей: среднее число аллелей на локус (А
), полиморфность (Р
) и ожидаемая гетерозиготность (Не
). Для расчета ожидаемой гетерозиготности Не
по каждому локусу использовали формулу
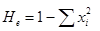 , ,
где хi
– частота i
-того аллеля. Показатель средней ожидаемой гетерозиготности вычислялся как
 , ,
где Hj
– гетерозиготность j-того локуса, К
– количество исследованных локусов. Показатель полиморфности (Р
) рассчитывали путем деления числа полиморфных локусов на общее количество исследованных локусов, а параметр среднего числа аллелей на локус (А
) путем деления количества выявленных аллелей на общее количество исследованных локусов. Полиморфность подсчитывалась по 99% критерию (частота наиболее общего аллеля не превышала 99%), а среднее число аллелей на локус по всем обнаруженным.
Для оценки генетической дифференциации среди таксонов лиственниц использовался коэффициент генетической дистанции Неи (D
N
), который учитывает различия в аллельных частотах всех проанализированных локусов:

DN
= - lnIN
,
где xij
и yij
— частоты i
-го аллеля j
-го локуса сравниваемых таксонов. Если D
N
равно 0, то таксоны идентичны. Чем больше значение D
N
, тем менее они родственны.
Считается, что коэффициент дистанции Неи самый точный, и поэтому он используется практически всеми исследователями. Дендрограмма наглядно демонстрирующая общую картину генетических взаимоотношений между исследованными таксонами на основании полученных коэффициентов D
N
строились методом невзвешенного парно-группового кластерного анализа (UPGMA) (Sneath, Sokal 1973).
2. Результаты и их обсуждение
В ходе электрофоретического анализа 12 ферментных систем у трех лиственниц нами было обнаружено 62 различных электрофоретических варианта. Наглядное изображение электрофоретических спектров малатдегидрогеназы и глюкозофосфатизомеразы исследованных видов с выявленными электрофоретическими вариантами приведено на рис. 1, 2. Проведенный генетический анализ показал, что все обнаруженные нами электрофоретические варианты кодируются 62 аллелями 20 локусов (Гончаренко, Шевцова, 2004). Все эти аллельные варианты и их относительная электрофоретическая подвижность наглядно изображены на рис. 3. Обозначение аллелей дано по общепринятой номенклатуре Пракаша (Prakashetal., 1969).

Наиболее часто встречающийся аллель локуса у лиственницы сибирской получил цифровой символ 1.00. Остальные аллели этого локуса, встреченные у проанализированных нами видов, включая L.sibirica,
обозначались цифровыми символами в соответствии с их электрофоретической подвижностью относительно аллеля 1.00]. Например, Gpi0.65
– это обозначение гена, кодирующего аллозим, подвижность которого на 35% медленнее Gpi1.00
. Нулевые аллельные варианты обозначены символом 0.
Следует подчеркнуть, что вопросы генетической детерминации ген-ферментных систем в определенной степени отражены в ряде работ, посвященных анализу некоторых лиственниц Палеарктики (Mejnartowicz, Bergmann, 1975;Ларионова, Милютин, 1981; Шурхал и др., 1989; Тимерьянов и др., 1994; Потенко, Разумов 1996; Гончаренко, Силин, 1997; Fins, Seeb, 1986; Lewandowski, Mejnartowicz, 1991, 1994; Гончаренко, Шевцова 2004). Таким образом, к настоящему времени разными лабораториями разработаны методы выявления достаточно большого набора изоферментных локусов, являющихся надёжными генетическими маркёрами. Всё это даёт возможность объективно проводить оценку уровня генетического полиморфизма и генетическогородства в различных таксонах лиственниц и, тем самым, позволяет решать как различные вопросы генетической изменчивости, так и сложные вопросы систематикии эволюционной филогении представителей рода Larix
.
В результате электрофоретического анализа ферментных систем у трех лиственниц нами было обнаружено 40 различных электрофоретических аллельных вариантов у L. sibirica
, 29у L. sukaczewii
и 48 уL. dahurica.

Частоты встречаемости всех 62 аллелей, отражающие генетические структуры трех видов лиственниц, представлены в табл. 2. Из таблицы хорошо видно, что L
.
sibirica
и L
.
sukachevii
имеют крайне сходные генетические структуры практически по всем локусам.
Четкие различия в аллельных частотах, превышающие 20%, наблюдались только по двум локусам: Aat-1и Mdh-3. Более существенные отличия в генетических структурах были выявлены между L
.
sibirica
и L. dahurica.
Здесь четкие различия между двумя видами найдены по аллелям четырех локусов: Aat-1, Mdh-3, Pgm-1 и 6-Pgd-2. Что касается пары L. dahurica
и L
.
sukachevii
,
то в этом случае существенные различия в аллельных частотах, превышающие 20%, наблюдались уже по пяти локусам. Причем по Aat-1 эти различия достигли 52%, а по Mdh-3 даже 64% (табл. 2).
Локусы, различия по которым достигают 95% и более (Ayala, Powell 1972; Левонтин, 1978) предложили считать «диагностическими», т.е. такими, по которым таксоны различаются качественно. Наличие «диагностических» локусов обычно характеризует четкие виды с полностью сформировавшимся репродуктивным барьером. В нашем случае разницу в частотах аллелей по локусам Aat-1 и Mdh-3 между L
.
sukachevii
с одной стороны и L. dahurica
с другойможно рассматривать только как тенденцию к образованию диагностических.
Таким образом, только у пары L. dahurica -
L
.
sukachevii
из 20 проанализированных локусов существенная генетическая дифференциация обнаружилась в аллельных частотах двух генов (табл. 2).
Для более точной оценки степени генетической дифференциации мы использовали коэффициент генетической дистанции Неи (Nei, 1972), который учитывает отличия не только по диагностическим и существенно различающимся, но и по всем исследованным локусам.
Таблица 2
.
Аллельные частоты по 20 локусам у лиственницы сибирской, лиственницы Сукачёва и лиственницы даурской.
| Локус |
Аллели |
Виды |
Локус |
Аллели |
Виды |
| L. sib.
|
L. suk
.
|
L. dah.
|
L. sib.
|
L. suk
.
|
L. dah.
|
| Aat-1
|
n |
45 |
23 |
90 |
Gpi
|
n |
44 |
23 |
90 |
| 1.00 |
0.676 |
0.478 |
0.995 |
0.65 |
0.011 |
0.000 |
0.000 |
| 1.15 |
0.324 |
0.522 |
0.005 |
0.75 |
0.044 |
0.000 |
0.078 |
| Aat-2
|
n |
46 |
23 |
90 |
0.90 |
0.117 |
0.000 |
0.000 |
| 0.80 |
0.000 |
0.000 |
0.005 |
1.00 |
0.828 |
1.000 |
0.878 |
| 1.00 |
0.870 |
1.000 |
0.992 |
1.10 |
0.000 |
0.000 |
0.022 |
| 1.20 |
0.130 |
0.000 |
0.005 |
1.20 |
0.000 |
0.000 |
0.022 |
| Aat-3
|
n |
46 |
23 |
90 |
6-Pgd-1
|
n |
44 |
23 |
90 |
| 1.00 |
1.000 |
1.000 |
0.989 |
1.00 |
1.000 |
0.957 |
1.000 |
| 2.00 |
0.000 |
0.000 |
0.011 |
1.10 |
0.000 |
0.043 |
0.000 |
| Skdh
|
n |
44 |
23 |
90 |
6-Pgd-2
|
n |
43 |
23 |
90 |
| 0.75 |
0.000 |
0.043 |
0.000 |
0.80 |
0.152 |
0.174 |
0.000 |
| 0.95 |
0.000 |
0.000 |
0.022 |
1.00 |
0.722 |
0.826 |
1.000 |
| 1.00 |
1.000 |
0.957 |
0.934 |
1.10 |
0.126 |
0.000 |
0.000 |
| 1.10 |
0.000 |
0.000 |
0.044 |
Lap-1
|
n |
47 |
23 |
90 |
| Gdh
|
n |
45 |
23 |
90 |
0 |
0.000 |
0.000 |
0.017 |
| 1.00 |
1.000 |
1.000 |
1.000 |
0.95 |
0.000 |
0.000 |
0.017 |
| Idh
|
n |
45 |
23 |
90 |
1.00 |
1.000 |
1.000 |
0.966 |
| 1.00 |
1.000 |
1.000 |
1.000 |
Lap-2
|
n |
47 |
23 |
90 |
| Mdh-1
|
n |
47 |
23 |
90 |
0.95 |
0.000 |
0.000 |
0.105 |
| 0.80 |
0.000 |
0.000 |
0.022 |
1.00 |
0.989 |
1.000 |
0.856 |
| 1.00 |
1.000 |
1.000 |
0.973 |
1.05 |
0.011 |
0.000 |
0.039 |
| 1.20 |
0.000 |
0.000 |
0.005 |
Pgm-1
|
n |
45 |
23 |
90 |
| Mdh-2
|
n |
47 |
23 |
90 |
0 |
0.012 |
0.000 |
0.011 |
| 0.80 |
0.000 |
0.000 |
0.022 |
0.90 |
0.033 |
0.000 |
0.117 |
| 1.00 |
1.000 |
1.000 |
0.778 |
1.00 |
0.955 |
1.000 |
0.650 |
| 1.15 |
0.000 |
0.000 |
0.195 |
1.03 |
0.000 |
0.000 |
0.133 |
| 1.35 |
0.000 |
0.000 |
0.005 |
1.10 |
0.000 |
0.000 |
0.078 |
| Mdh-
3
|
n |
47 |
23 |
90 |
1.20 |
0.000 |
0.000 |
0.011 |
| 0 |
0.000 |
0.000 |
0.005 |
Pgm-2
|
n |
45 |
23 |
90 |
| 0.50 |
0.150 |
0.587 |
0.000 |
1.00 |
0.988 |
1.000 |
0.989 |
| 0.80 |
0.310 |
0.087 |
0.027 |
1.10 |
0.012 |
0.000 |
0.011 |
| 1.00 |
0.540 |
0.326 |
0.963 |
Fl-Est
|
n |
40 |
23 |
90 |
| 1.25 |
0.000 |
0.000 |
0.005 |
0.50 |
0.025 |
0.000 |
0.000 |
| Mdh-
4
|
n |
47 |
23 |
90 |
0.80 |
0.055 |
0.043 |
0.000 |
| 0 |
0.053 |
0.000 |
0.000 |
1.00 |
0.756 |
0.696 |
1.000 |
| 1.00 |
0.885 |
1.000 |
0.955 |
1.20 |
0.167 |
0.261 |
0.000 |
| 1.10 |
0.105 |
0.000 |
0.000 |
Aco
|
n |
43 |
23 |
90 |
| 1.40 |
0.052 |
0.000 |
0.005 |
0.95 |
0.126 |
0.000 |
0.000 |
| Sdh
|
n |
44 |
23 |
90 |
1.00 |
0.874 |
0.978 |
1.000 |
| 1.00 |
1.000 |
1.000 |
1.000 |
1.05 |
0.000 |
0.022 |
0.000 |
| L
.
sib
.-
лиственница сибирская, L
.
suk
.-
лиственница Сукачева, L
.
dah
.-
лиственница даурская, n – количество проанализированных деревьев. |
Значения коэффициентов дистанции (DN
) по 20 генам для трех лиственниц Палеарктики представлены в табл. 3. Как и следовало ожидать, исходя из сравнения аллельных частот, наименьшее значение коэффициента дистанции Неи выявлено между лиственницей сибирской и Сукачева, средняя величина DN
для которых составила 0.015 (табл. 3). Иными словами, эти лиственницы, отличаются лишь по 1,5% своих локусов, что характерно только для географически связанных популяций одного вида, значение DN
между которыми находится в пределах от 0 до 0.030 (Yeh, O'Malley, 1980; Guries, Ledig, 1982; Wheeler, Guries, 1982; Dancik, Yeh, 1983; Plessas, 1986; Ross, Hawkins, 1986; Yehetal, 1986; Merkle, Adams, 1987; Cheliaketal., 1988; Millar et al., 1988; Gianninietal 1991; Goncharenkoetal, 1993a, b, 1994, Boscherinietal, 1994; Hawley, DeHayes, 1994; Kimetal, 1994; Teisseireetal, 1995; Silin, Goncharenko 1996; Rajaetal, 1997), тогда как дистанция даже между молодыми видами с неполным репродуктивным барьером обычно составляет 0.10 (Dancik, Yeh, 1983; Wheeler et al., 1983; Wheeler, Guries, 1987; Millar et al., 1988; Hawley, DeHayes, 1994; Goncharenko et al., 1992, 1995; Гончаренко 1999).
Исходя из всей совокупности полученных нами генетических данных, можно сделать однозначное заключение о том, что нет никаких оснований считать лиственницу сибирскуюи лиственницу Сукачева самостоятельными видами, поскольку между ними выявлено огромное сходство генетических структур практически по всем проанализированным локусам, а значение генетической дистанции Неи составило лишь 0.015. Все это еще раз подтверждает мнение тех исследователей, которые рассматривают две лиственницы только как морфологически обособленные популяции одного вида L
.
sibirica
Ledeb.(Бобров, 1972, Милютин, Муратова 1993).

Как следует из данных приведенных в табл. 3, генетическая дистанция DN
между уральской L.
sukaczevii
и дальневосточной L. dahurica
оказалась 0.052, что почти в 4 раза выше, чем дистанция между L
.
sibirica
и L
.
sukachevii
. Это значение DN
является немногим выше, чем дистанция Неи, обычно наблюдаемая между подвидами и географическими расами у хвойных, но несколько ниже, чем DN
, характерная для близкородственных видов, установленная в основном для сосен и елей (Wheeler, Guries 1982, Schiller at al. 1986, Conkle at al. 1988, Wang atal. 1990, Goncharenkoatal. 1995, Гончаренко 1999). Интересно отметить, что генетическая дистанция между географически более близкими западносибирской L
.
sibirica
иL. dahurica
меньше, чем между географически более удаленными L
.
sukachevii
и L. dahurica
и составляет 0.029 (табл.3).
С использованием невзвешенного парно-группового метода кластерного анализа (UPGMA) на базе данных табл. 3 нами построена дендрограмма, иллюстрирующая степень генетической дифференциации у трех исследованных лиственниц Палеарктики (рис. 4). Из дендрограммы хорошо видно что, как и следовало ожидать, L
.
sibirica
и L
.
sukachevii
объединились в один тесный кластер с дистанцией Неи между ними, составившей 0.015 и лишь затем к единому таксону урало-сибирских лиственниц подсоединяется дальневосточная L. dahurica
с дистанцией, превышающей 0.041.

Генетическая дистанция DN
между урало-сибирской L
.
sibirica
и дальневосточной L. dahurica
оказалась несколько ниже, чем DN
, характерная для близкородственных видов хвойных. Тем не менее, полученные нами генетические данные не позволяют опровергнуть самостоятельность видового статуса L. dahurica
поскольку она отличается от L
.
sibirica
в целом боле чем 4% своего генома и имеет 2 существенно дифференцированных "полудиагностических" локуса Aat-1 и Mdh-3 (см. табл. 2).
Низкая генетическая дифференциация по структурным генам, выявленная нами между двумя общепризнанными видами, скорее всего, связана с отсутствием географической изоляции и репродуктивного барьера между ними. Как указывал в своей фундаментальной работе ведущий исследователь хвойных Палеарктики Е.Г. Бобров, на протяжении всей западной границы L. dahurica
легкосмешиваетсяв процессе интрогрессив-ной гибридизации с L
.
sibirica
, образуя гибридную лиственницу, получившую название лиственницы Чекановского – L
.
x
czekanovskii
Szaf. Причем полоса интрогрессии этих двух видов распространяется на несколько сотен километров (Бобров, 1972, 1978). По-видимому, именно вследствие интенсивной гибридизации между двумя видами происходит обмен генетическим материалом, который сглаживает и уменьшает генетическую дифференциацию.
Важной эволюционно сложившейся характеристикой является уровень генетической изменчивости, который определяется при помощи основных показателей полиморфизма. Считается, что наиболее точные оценки данных показателей достигаются при анализе 18-20 и более случайно выбранных из генома локусов (Левонтин 1978, Айала 1984). Значение основных показателей генетического полиморфизма для представителей рода Larix
проанализированных с использованием 18 и более генов приведены в таблице 4.
Как видно из табл. 4, у проанализированных нами видов наименьшим уровнем генетической изменчивости характеризуется лиственница Сукачева, поскольку у неё доля полиморфных локусов, Р
составила 0.350, среднее число аллелей на локус, А
1.45, а средняя ожидаемая гетерозиготность, Не
оказалась 0.099. У лиственницы сибирской эти показатели оказались существенно выше: гетерозиготность Не
превысила 15%, а число аллелей на локус составило 2.05. Что касается лиственницы даурской, то здесь картина оказалась сложнее, т.к. показатель А
превысил 2.4, в то время как гетерозиготность едва достигла 9% (табл. 4).
Таблица 4.
Основные параметры генетического полиморфизма у лиственниц, полученные при анализе 18 и более локусов.
| Вид |
Доля полиморфных локусов, Р99
|
Среднее число аллелей
на локус, А
|
Средняя ожидаемая гетерозиготность, Не |
Ссылка |
| L. dahurica
|
0.500 |
2.40 |
0.088 |
Наши данные |
| L. dahurica
|
0.601 |
2.46 |
0.129 |
Потенко, Разумов 1996 |
| L. dahurica
|
0.780 |
2.00 |
0.125 |
Ларионова и др. 2004 |
| 0.119* |
| L
.
decidua
|
0.576 |
1.90 |
0.157 |
Lewandowski,Mejnartowicz 1991 |
| L
.
kurilensis
|
0.500 |
1.70 |
0.166 |
Гончаренко, Силин 1997 |
| L
.
kaempferi
|
0.500 |
1.60 |
0.119 |
Гончаренко, Силин 1997 |
| L
.
laricina
|
0.500 |
1.80 |
0.220 |
Cheliak et al. 1988 |
| L.laricina
|
0.470 |
1.70 |
0.151 |
Ying, Morgenstern 1991 |
| 0.211* |
| L.occidentalis
|
0.304 |
1.50 |
0.082 |
Fins, Seeb 1986 |
| L. sibirica
|
0.600 |
2.05 |
0.153 |
Нашиданные |
| L. sibirica
|
0.570 |
1.70 |
0.129 |
Шурхал и др. 1989 |
| 0.141* |
| L
.
sukaczevii
|
0.350 |
1.45 |
0.099 |
Наши данные |
| L
.
sukaczevii
|
0.670 |
1.15 |
0.043 |
Тимерьянов 1996 |
| L
.
sukaczevii
|
0.319 |
1.55 |
0.122 |
Путенихин, Старова 1991 |
| 0.071* |
| Среднее
|
0.133
|
| * В случае, когда по виду было проведено несколько исследований, то рассчитывалась средне взвешенная величина ожидаемой гетерозиготности, зависящая от количества проанализированных локусов и деревьев. |
Интересно отметить, что совпадающие с нашими показатели изменчивости у лиственницы Сукачева получены и в одной работе уральских исследователей (Путенихин 1991), в тоже время в другой (Тимерьянов и др., 1996) – гетерозиготность Не
была аномально низкой и составила только 4%, а показатель А
едва превысил 1.1. В исследовании Шурхала и др. (1989), посвященном L
.
sibirica
,
полиморфизм оказался достаточно высок, хотя и чуть ниже выявленного нами (табл. 4). Что касается L. dahurica,
то в работах Потенко, Разумова (1996) и Ларионовой и др. (2004) гетерозиготность в популяциях этого вида превысила 12%, хотя количество аллелей на локус колебалось в разных популяциях от 2.0 до 2.4. (табл. 4).
Анализируя данные таблицы 4, по роду Larix
в целом следует сказать, что наиболее высокий уровень гетерозиготности (Не
=22%) среди всех проанализированных в настоящее время лиственниц отмечен в одной из работ для L
.
laricina
(Cheliaketal. 1980), которая имеет широкое распространение на североамериканском континенте, однако в другой работе (YingL., Morgenstern, 1991), посвященной генетическому исследованию этого же вида, показатели гетерозиготности были существенно нижеи хорошо соотносятся с таковыми, полученными нами по западносибирской L
.
sibirica
.
Наименьший среди исследованных лиственниц уровень изменчивости (Не
=8%)выявлен у L.
occidentalis
(Fins, Seeb 1986), имеющей небольшой ареал в Северной Америке (табл. 4). Интересно отметить, что в целом у 8 проанализированных видов рода Larix
51.8% локусов находится в полиморфном состоянии, а каждая особь гетерозиготна в среднем по 13.3% своих генов. В этом отношении лиственницы по уровню полиморфизма уступают соснам, у которых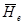 ,рассчитанная для 30 видов, составила 15.8%, елям, гдедля 12 видов составила 18%, и пихтам, гдедля 8 видов составила 15.4% (см. сводки в Гончаренко, Силин 1997; Гончаренко, Савицкий 2000; Гончаренко, Падутов 2001). ,рассчитанная для 30 видов, составила 15.8%, елям, гдедля 12 видов составила 18%, и пихтам, гдедля 8 видов составила 15.4% (см. сводки в Гончаренко, Силин 1997; Гончаренко, Савицкий 2000; Гончаренко, Падутов 2001).
Заключение
Таким образом, в результате проведенного нами генетического анализа природных популяций трех лиственниц, произрастающих в сибирско-дальневосточном регионе Палеарктики, с использованием 20 аллозимных локусов установлено, что выявленная близкая генетическая структура у L.
sibirica
и L
.
sukaczevii
с дистанцией Неи (DN
), равной всего 0.015 дает основание рассматривать их только как морфологически обособленные популяции одного видаLarix
sibirica
. Низкая генетическая дифференциация между L
.
sibirica
иL. dahurica
с DN
, составившей 0.052, свидетельствует об интенсивной гибридизации и обмене генетическим материалом между двумя географически связанными видами, однако не позволяет опровергнуть самостоятельность видового статуса L. dahurica.
Обнаружены высокие значения показателей изменчивости лиственницы сибирской и лиственницы даурской, величины которых соотносятся с таковыми, полученными для видов с широкими ареалами, в то время как у лиственницы Сукачева установлен наименьший уровень генетической изменчивости среди исследованных видов.
л
итература
1. Бобров Е. Г. Лесообразующие хвойные СССР. Л.: Наука, 1978. 190 с.
2. Гончаренко Г.Г., Падутов В.Е., Потенко В.В. Руководство по исследованию хвойных видов методом электрофоретического анализа изоферментов. Гомель, 1989.
3. Гончаренко Г.Г., Силин А.Е. К вопросу о генетической изменчивости и дифференциации лиственницы курильской (Larix kurilensis
Mayr.) и лиственницы японской (Larix kaempferi
Sarg.)// Докл. АН России. - 1997. Т. 355. № 3. C. 835-838.
4. Гончаренко Г.Г. Геносистематика и эволюционная филогения лесообразующих хвойных Палеарктики. Минск: Тэхналогiя, 1999. 188 с.
5. Гончаренко Г.Г.,Шевцова Л.А. К вопросу о генетико-таксономических взаимоотношениях между лиственницей сибирской (Larix
sibirica
Ledeb.) и лиственницей Сукачёва (Larix
sukachevii
Dyl.) // Известия Гомельского государственного университета им. Ф. Скорины. – Гомель, 2004. - №3. - С. 43-50.
6. Дылис Н.В.Лиственница Восточной Сибири и Дальнего Востока. М.: Изд-во АН СССР. – 1961. – 210 с.
7. Козубов Г.М., Муратова Е.Н. Современные голосеменные. – Л.: Наука, 1986. – 192 с.
8. Козыренко М.М., Артюкова Е.В., Реунова Г.Д., Лёвина Е.А., Журавлев Ю.Н. Генетическая изменчивость и взаимоотношения лиственниц Сибири и Дальнего востока по данным RAPD-анализа // Генетика, 2004. – Т. 40, №. 4. – С. 506-515.
9. Ларионова А.Я., Милютин Л.И. Исследование внутривидовой дифференциации сибирской лиственницы с помощью метода изоэнзимных спектров // Лесоведение, 1981. № 2. –С. 3-11.
10. Ларионова А.Я, Яхнева Н.В., Абаимов А.П.Генетическое разнообразие и дифференциация популяций лиственницы Гмелина в Евенкии (Средняя Сибирь) // Генетика, 2004. – Т. 40, №. 10. – С. 1370-1377.
11. Левонтин Р. Генетические основы эволюции: Пер. с англ.- М.: Мир,1978.- 352 с.
12. Номенклатура ферментов. – М.: ВИНИТИ, 1979.- 320 с.
13. Потенко В.В., Разумов П.Н. Генетическая изменчивость и популяционная структура лиственницы даурской на территории Хабаровского края // Лесоведение, 1996.– №5.– С.11-18.
14. Пугач Е.А. Лиственница // Лесная энциклопедия. Т. 2. Гл. ред. Воробьев Г. И. М.: Сов. Энциклопедия, 1985. – С. 13-15.
15. Тимерьянов А.Ш., Шигапов З.Х., Янбаев Ю.А. Генетическая изменчивость лиственницы Сукачева (Larix
sukaczewii
dyl.) на южном Урале. Механизм генного контроля изоферментных систем // Генетика, 1994. –Т. 30, № 9. –С. 1243-1247.
16. Тимерьянов А.Ш., Старова Н.В., Бахтиярова Р.М. Генетическая изменчивость лиственницы Сукачева (Larix
sukaczewii
dyl.) на южном Урале. II Уровни изоферментной изменчивости в природных популяциях // Генетика, 1996. –Т. 32, № 2. –С. 267-271.
17. Тимерьянов А.Ш. Генетическая изменчивость лиственницы Сукачева (Larix
sukaczewii
dyl.) на южном Урале. Пространственная дифференциация популяций // Генетика, 1996. Т. 32, № 5. С. 663-667.
18. Шурхал А.В., Подогас А.В., Семериков В.Л., Животовский Л.А. Аллозимный полиморфизм лиственницы сибирской Larix
sibirica
//
Генетика, 1989. – Т. 25, № 10. – С. 1899-1901.
19. Ayala F.J., Powell J.R. Allozymes as diagnostic characters of sibling species of Drosophila // Proc. Nat. Acad. Sci.- 1972.- V. 69.- P. 1094-1096.
20. Boscherini G., M. Morgante, P. Rossi, G.G. Vendramin. Allozyme and chloroplast DNA variation in Italian and Greek populations of Pinus leucodermis.
Heredity –V.73.–P.284-290. – 1994.
21. Cheliak W.M., Murray G., Pitel J.A. Genetic effects of phenotypic selection in white spruce// For. Ecol. Manage.- 1988. - V. 24. - P. 139-149.
22. Dancik B.P., Yeh F.C. Allozyme variability and evolution of lodgepole pine Pinus contorta var. latifolia
) and jack pine (P.banksiana
) in Alberta // Can. J. Genet. Cytol. - 1983.- V. 25.- P. 57-64.
23. FinsL., SeebL. Geneticvariationinallozymesofwesternlarch // Can. J. For. Res, 1986. –V. 16. – P. 1013-1018.
24. Giannini R.M., Morgante and G.G. Vendramin. Allozyme variation in Italian populations of Picea abies
(L.) Karst. Silvae Genetica. – 1991a. – V.40. – P.160-166.
25. Goncharenko G.G., Padutov V.E., Silin A.E. Allozyme variation in natural population of Eurasian pines. I. Population structure, genetic variation, and differentiation in Pinus pumila
(Pall.) Regel from Chukotsk and Sakhalin // Silvae Genet. – 1993a. – Vol.42 – P. 237-246.
26. Goncharenko G. G., Padutov V. E., Silin A. E. Allozyme variation in natural population of Eurasian pines. 2 Genetic variation, diversity, differentiation and gene flow in Pinus sibirica Du Tour in some lowland and mountain populations// Silvae Genet. 1993 b. V. 42. P. 246-253.
27. Goncharenko G.G., Silin A.E., Padutov V.E. Allozyme variation in natural populations of Eurasian pines. III. Population structure, diversity, differentiation and gene flow in central and isolated population of Pinus sylvestris
L. in Eastern Europe and Siberia // Silvae Genet. – 1994. – Vol.43. –P.119-132.
28. Guries R.P., Ledig F.T. Genetic diversity and population structure in Pitch pine (Pinus rigida Mill.)// Evolution.- 1982.- V. 36.- P. 387-402.
29. Hawley G.J., DeHayes D.H. Genetic diversity and population structure of red spruce (Picea rubens)
// Can. J. Bot.-1994 – Vol.72. – 1778-1786.
30. Kim Z.-S., Lee S.-W., Lim J.-H. e.a. Genetic diversity and structure of Pinus koraiensis
(Sieb. & Zucc.) in Korea // Forest Genetics. – 1994. – Vol.1. –P.41-49.
31. Lewandowski A., Mejnartowicz L. Inheritance of allozymes in Larix decidua
Mill. // Silvae Genet. 1990. Vol. 39. № 5-6. P. 184-188.
32. Lewandowski A., Mejnartowicz L. Levels and patterns of allozyme variation in some European larch (Larix decidua
) populations // Hereditas. 1991. 115. P. 221-226.
33. LewandowskiA. GeneticrelationshipsbetweenEuropeanandSiberianlarch, Larix
spp
. (Pinaceae
), studied by allozymes. Is the Polish larch a hybrid between these two species? // Pl. Syst. Evol. 1997. Vol. 204. P. 66–73.
34. Merkle S.A., Adams W.T. Patterns of allozyme variation within and among Douglas-fir breeding zones in southwest Oregon// Can. J. For. Res.- 1987.- V. 17.- P. 402-407.
35. Mejnartowicz L., Bergmann F. Genetic studies on European larch (Larix decidua
Mill.) employing isoenzyme polymorphisms // Genetica polonica, 1975. Vol. 16. № 1. P. 29-35.Paule L., Gömöry D. // Population genetics and genetic conservation of forest trees. 1995. P. 321–328.
36. Millar C.I., Strauss S.H., Conkle M.T., Westfall R.P. Allozyme differentiation and biosystematics of the Californian closed cone pines (Pinus subsect. Oocarpae) // Systematic Botany.- 1988.- V. 13 - P. 351-370.
37. Nei M. Genetic distance between populations // Amer. Nat.- 1972.- V. 106.- P. 283-292.
38. Plessas M.E., Strauss S.H. Allozyme differentiation among populations stands and cohorts on Monterey pine // Can. J. For. Res.- 1986.- V. 16.- P. 1155-1164.
39. Prakash S., Lewontin R.C., Hubby J.L. A molecular approach to the study of genic heterozygosity in natural populations. IV. Patterns of genic variation in central, marginal and isolated populations of Drosophila pseudoobscura// Genetics.- 1969.- V. 61.- P. 841-858.
40. Raja R.G., Tauer C.G., Wittwer R.F., Huang Y. Isoenzyme variation and genetic structure in natural populations of shortleaf pine (Pinus echinata
) // Can. J. For. Res. – 1997. – Vol.27. – P.740-749.
41. Ross H.A., Hawkins J.L. Genetic variation among local populations of Jack pine (Pinus banksiana) // Can. J. Genet. Cytol. - 1986. - V. 28. - P. 453-458.
42. Semerikov V.L., Zhang H., Sun M., Lascoux M. Conflicting phylogenies of Larix (Pinaceae)
based on cytoplasmic and nuclear DNA //
Mol. Phyl. Evol., 2003. Vol. 27. № 2. Р. 173-184.
43. Silin A.E., Goncharenko G.G., Allozyme variation in natural populations of Eurasian pines. IV. Population structure and isolated populations of Pinus nigra
Arnold on the Crimean peninsula. // Silvae Genet. V.45. – P.67-75. 1996.
44. Sneath P. H. A., Sokal R. R. Numerical Taxonomy: the Principles and Practice of Numerical Classification. San Francisco: W. H. Freeman, 1973. 573 p.
45. Teisseire H., Fady B., Pichot C. Allozyme variation in five French populations of Aleppo pine ( Pinus halepensis Miller)
// Forest Genetics. 1995. V. 2. P. 225-236.
46. Wheeler N.C., Guries R.P. Population structure, genic diversity, and morphological variation in Pinus contorta
Dougl.// Can. J. For. Res. - 1982. - V. 12. - P. 595-606.
47. Yeh F.C., Khalil M.A.K., El-Kassaby Y.A., Trust D.C. Allozyme variation in Picea mariana from Newfoundland: genetic diversity, population structure, and analysis of differentiation// Can. J. For. Res. - 1986. - V. 16. - P. 713-720.
48. Yeh F.C., O'Malley D. Enzyme variations in natural populations of Douglas-fir, Pseudotsuga menziesii
(Mirb.) Franco, from British Columbia. I. Genetic variation patterns in coastal populations// Silvae Genet. - 1980. - V. 29. - P. 83-92.
49. Wheeler N.C., Guries R.P., O'Malley D.M. Biosystematics of the Genus Pinus, Subsection Contortae // Biochem. Syst. Ecol.- 1983.- V. 11.- P. 333-340.
50. Wheeler N.C., Guries R.P. A quantitative measure of introgression between lodgepole and jack pines // Can. J. Bot.- 1987.- V.65.- P. 1876-1885.
51. Goncharenko G.G., Padutov V.E., Silin A.E. Population structure, gene diversity, and differentiation in natural populations of Cedar pines (Pinus subsect.
Cembrae, Pinaceae) in the USSR// Pl. Syst. Evol. - 1992. - V. 182. - P. 121-134.
52. Goncharenko G. G., Silin A. E., Padutov V. E. Intra-and interspecific genetic differentiation in closely related pines from Pinus subsection Sylvestres (Pinaceae ) in the former Soviet Union// PL. Syst. Evol. 1995.V. 194. P. 39-54.
53. Милютин Л.И., Муратова Е.Н., Ларионова А.Я. Генетико-таксономический анализ популяций лиственниц сибирской и Сукачева // Лесоведение, 1993. – №5. – С. 55-63.
54. Schiller G., Conkle M.T., Grunwald C. Local differentiation among Mediterranean of Aleppo pine in their isoenzymes// Silvae Genet. -1986. -V. 35.No.1.- P. 11-19.
55. Conkle M.T., Schiller G., Grunwald C. Electrophoretic analysis of diversity and phylogeny of Pinus brutia
Ten. and closely related taxa // Systematic Botany.- 1988.- V. 13.- P. 411-424.
56. Wang X.-R., Szmidt A.E., Lewandowski A., Wang Z.-R. Evolutionary analysis of Pinus densata
Masters,a putative Tertiary hybrid. 1. Allozyme variation. // Theor. Appl.Genet.80: 635-640 (1990).
57. Бобров Е.Г. История и систематика лиственниц. Л.:Наука, 1972. – 96 с.
58. Айала Ф. Введение в молекулярную и эволюционную генетику: Пер. с англ. - М.: Мир, 1984. - 230 с.
59. Путенихин В.П., Фарукшина Г.Г., Шигапов З.Х. Лиственница Сукачева на Урале: изменчивость и популяционно-генетическая структура.– М.: Наука, 2004. – 276 с.
60. Cheliak W.M., Wang J., Pitel J.A. Population structures and genic diversity in tamarack, Larix laricina
(Du Roi) K. Koch.// Can. J. For. Res. - 1988.- V. 18.- P.1318-1324.
61. Ying L., Morgenstern E.K. The population structure of Larix laricina
in New Brunswick, Canada// Silvae Genet. - 1991. - V.40. - No.5/6. - P.180-184.
62. Гончаренко Г.Г., Силин А.Е. Популяционная и эволюционная генетика сосен Восточной Европы и Сибири. - Минск: Тэхналогiя, 1997. 192с.
63. Гончаренко Г.Г., Савицкий Б.П. Популяционно-генетические ресурсы пихты белой в Беларуси. - Гомель: ИЛ НАН Беларуси, 2000. 122 с.
64. Гончаренко Г.Г. , Падутов В.Е. Популяционная и эволюционная генетика елей Палеарктики. Гомель: ИЛ НАН Беларуси, 2001. 188 с.
|