1. Биотехнологии
1.1. Основные понятия
Биотехнология - это производственное использование биологических агентов или их систем для получения ценных продуктов и осуществления целевых превращений.
Биологические агенты в данном случае - микроорганизмы, растительные или животные клетки, клеточные компоненты (мембраны клеток, рибосомы, митохондрии, хлоропласты), а также биологические макромолекулы (ДНК, РНК, белки - чаще всего ферменты). Биотехнология использует также вирусную ДНК или РНК для переноса чужеродных генов в клетки.
Человек использовал биотехнологию многие тысячи лет: люди пекли хлеб, варили пиво, делали сыр, используя различные микроорганизмы, при этом, даже не подозревая об их существовании.
Собственно сам термин появился в нашем языке не так давно, вместо него употреблялись слова «промышленная микробиология», «техническая биохимия» и др.
Вероятно, древнейшим биотехнологическим процессом было сбраживание с помощью микроорганизмов. В пользу этого свидетельствует описание процесса приготовления пива, обнаруженное в 1981г. при раскопках Вавилона на дощечке, которая датируется примерно 6-м тысячелетием до н. э.
Не менее древними биотехнологическими процессами являются виноделие, хлебопечение, и получение молочнокислых продуктов.
В традиционном, классическом, понимании биотехнология - это наука о методах и технологиях производства различных веществ и продуктов с использованием природных биологических объектов и процессов.
Термин «новая» биотехнология в противоположность «старой» биотехнологии применяют для разделения биопроцессов, использующих методы генной инженерии и более традиционные формы биопроцессов.
Так, обычное производство спирта в процессе брожения – «старая» биотехнология, но использование в этом процессе дрожжей, улучшенных методами генной инженерии с целью увеличения выхода спирта – «новая» биотехнология.
Биотехнология как наука является важнейшим разделом современной биологии, которая, как и физика, стала в конце XX в. одним из ведущих приоритетов в мировой науке и экономике.
Всплеск исследований по биотехнологии в мировой науке произошел в 80-х годах, но, несмотря на столь короткий срок своего существования, биотехнология привлекла пристальное внимание, как ученых, так и широкой общественности.
По прогнозам, уже в начале 21 века биотехнологические товары будут составлять четверть всей мировой продукции.
Что касается более современных биотехнологических процессов, то они основаны на методах рекомбинантных ДНК, а также на использовании иммобилизованных ферментов, клеток или клеточных органелл.
Современная биотехнология - это наука о генно-инженерных и клеточных методах создания и использования генетически трансформированных биологических объектов для улучшения производства или получения новых видов продуктов различного назначения.
1.2. Основные направления биотехнологий
Условно можно выделить следующие основные направления биотехнологии:
· биотехнология пищевых продуктов;
· биотехнология препаратов для сельского хозяйства;
· биотехнология препаратов и продуктов для промышленного и бытового использования;
· биотехнология лекарственных препаратов;
· биотехнология средств диагностики и реактивов.
Биотехнология также включает выщелачивание и концентрирование металлов, защиту окружающей среды от загрязнения, деградацию токсических отходов и увеличение добычи нефти.
1.3. Генная инженерия – основа биотехнологии
Генная инженерия — это область биотехнологий, включающая в себя действия по перестройке генотипов. Уже сегодня генная инженерия позволяет включать и выключать отдельные гены, контролируя, таким образом, деятельность организмов, а также — переносить генетические инструкции из одного организма в другой, в том числе – организмы другого вида. По мере того, как генетики всё больше узнают о работе генов и белков, всё более реальной становится возможность произвольным образом программировать генотип (прежде всего, человеческий), с лёгкостью достигая любых результатов: таких, как устойчивость к радиации, способность жить под водой, способность к регенерации повреждённых органов и даже бессмертие.
2. Генетическая инженерия
2.1. История генной инженерии
Генная инженерия появилась благодаря работам многих исследователей в разных отраслях биохимии и молекулярной генетики.
На протяжении многих лет главным классом макромолекул считали белки. Существовало даже предположение, что гены имеют белковую природу.
Лишь в 1944 году Эйвери, Мак Леод и Мак Карти показали, что носителем наследственной информации является ДНК.
С этого времени начинается интенсивное изучение нуклеиновых кислот. Спустя десятилетие, в 1953 году Дж. Уотсон и Ф. Крик создали двуспиральную модель ДНК. Именно этот год принято считать годом рождения молекулярной биологии.
На рубеже 50-60-х годов были выяснены свойства генетического кода, а к концу 60-х годов его универсальность была подтверждена экспериментально.
Шло интенсивное развитие молекулярной генетики, объектами которой стали кишечная палочка (E. Coli), ее вирусы и плазмиды.
Были разработаны методы выделения высокоочищенных препаратов неповрежденных молекул ДНК, плазмид и вирусов.
ДНК вирусов и плазмид вводили в клетки в биологически активной форме, обеспечивая ее репликацию и экспрессию соответствующих генов.
В 70-х годах был открыт ряд ферментов, катализирующих реакции превращения ДНК. Особая роль в развитии методов генной инженерии принадлежит рестриктазам и ДНК-лигазам.
Историю развития генетической инженерии можно условно разделить на три этапа:
Первый этап связан с доказательством принципиальной возможности получения рекомбинантных молекул ДНК in vitro. Эти работы касаются получения гибридов между различными плазмидами. Была доказана возможность создания рекомбинантных молекул с использованием исходных молекул ДНК из различных видов и штаммов бактерий, их жизнеспособность, стабильность и функционирование.
Второй этап связан с началом работ по получению рекомбинантных молекул ДНК между хромосомными генами прокариот и различными плазмидами, доказательством их стабильности и жизнеспособности.
Третий этап - начало работ по включению в векторные молекулы ДНК (ДНК, используемые для переноса генов и способные встраиваться в генетический аппарат клетки-реципиента) генов эукариот, главным образом, животных.
Формально датой рождения генетической инженерии следует считать 1972 год, когда в Стенфордском университете П. Берг и С. Коэн с сотрудниками создали первую рекомбинантную ДНК, содержавшую фрагменты ДНК вируса SV40, бактериофага и E. coli.
2.2. Цели и методы генной инженерии
Цель прикладной генетической инженерии заключается в конструировании таких рекомбинантных молекул ДНК, которые при внедрении в генетический аппарат придавали бы организму свойства, полезные для человека.
На технологии рекомбинантных ДНК основано получение высокоспецифичных ДНК-зондов, с помощью которых изучают экспрессию генов в тканях, локализацию генов в хромосомах, выявляют гены, обладающие родственными функциями (например, у человека и курицы). ДНК-зонды также используются в диагностике различных заболеваний.
Технология рекомбинантных ДНК сделала возможным нетрадиционный подход «белок-ген», получивший название «обратная генетика». При таком подходе из клетки выделяют белок, клонируют ген этого белка, модифицируют его, создавая мутантный ген, кодирующий измененную форму белка. Полученный ген вводят в клетку. Таким способом можно исправлять дефектные гены и лечить наследственные заболевания.
Если гибридную ДНК ввести в оплодотворенное яйцеклетку, могут быть получены трансгенные организмы, передающие мутантный ген потомками.
Генетическая трансформация животных позволяет установить роль отдельных генов и их белковых продуктов как в регуляции активности других генов, так и при различных патологических процессах.
Технология рекомбинантных ДНК использует следующие методы:
· специфическое расщепление ДНК рестрицирующими нуклеазами, ускоряющее выделение и манипуляции с отдельными генами;
· быстрое секвенирование всех нуклеотидов в очищенном фрагменте ДНК, что позволяет определить границы гена и аминокислотную последовательность, кодируемую им;
· конструирование рекомбинантной ДНК;
· гибридизация нуклеиновых кислот, позволяющая выявлять специфические последовательности РНК или ДНК с большей точностью и чувствительностью;
· клонирование ДНК: амплификация in vitro с помощью цепной полимеразной реакции или введение фрагмента ДНК в бактериальную клетку, которая после такой трансформации воспроизводит этот фрагмент в миллионах копий;
· введение рекомбинантной ДНК в клетки или организмы.
2.3. Возможности генной инженерии.
Значительный прогресс достигнут в практической области создания новых продуктов для медицинской промышленности и лечения болезней человека
В настоящее время фармацевтическая промышленность завоевала лидирующие позиции в мире, что нашло отражение не только в объёмах промышленного производства, но и в финансовых средствах, вкладываемых в эту промышленность (по оценкам экономистов, она вошла в лидирующую группу по объёму купли-продажи акций на рынках ценных бумаг). Важной новинкой стало и то, что фармацевтические компании включили в свою сферу выведение новых сортов сельскохозяйственных растений и животных, и тратят на это десятки миллионов долларов в год, они же мобилизировали выпуск химических веществ для быта. Добавок к продукции строительной индустрии и так далее. Уже не десятки тысяч, а возможно, несколько сот тысяч высококвалифицированных специалистов заняты в исследовательских и промышленных секторах фарминдустрии, и именно в этих областях интерес к геномным и генно-инженерным исследованиям исключительно высок. Очевидно поэтому любой прогресс биотехнологий растений будет зависеть от разработки генетических систем и инструментов, которые позволят более эффективно управлять трансгенами. Для чистого вырезания трансгенного ДНК в растительный геном, всё больше применяют заимствованные из микробной генетики системы гомологичной рекомбинации, такие как системы Cre-lox и Flp-frt. Будущее, очевидно, будет за управляемым переносом генов от сорта к сорту, основанного на применении предварительно подготовленного растительного материала, который уже содержит в нужных хромосомах участки гомологии, необходимого для гомологичного встраивания трансгена. Помимо интегративных систем экспрессии, будут опробованы автономно реплицирующиеся векторы. Особый интерес представляют искусственные хромосомы растений, которые теоретически не накладывают никаких ограничений на объём вносимой теоретической информации.
Кроме этого учёные занимаются поиском генов, кодирующих новые полезные признаки. Ситуация в этой области меняется радикальным образом, прежде всего, существованию публичных баз данных, которые содержат информацию о большинстве генов, бактерий, дрожжей, человека и растений, а также вследствие разработки методов, позволяющих одновременно анализировать экспрессию большого количества генов с очень высокой пропускной способностью. Применяемые на практике методы можно разделить на две категории:
Методы, позволяющие вести экспрессионное профилирование: субстракционная гибридизация, электронное сравнение EST-библиотек, «генные чипы» и так далее. Они позволяют устанавливать корреляцию между тем или иным фенотипическим признаком и активностью конкретных генов.
Позиционное клонирование, заключается в создании за счет инсерционного мутагенеза мутантов с нарушениями в интересующем нас признаке или свойстве, с последующим клонированием соответствующего гена как такового, который заведомо содержит известную последовательность (инсерция).
Вышеназванные методы не предполагают никаких изначальных сведений о генах, контролирующих тот или иной признак. Отсутствие рационального компонента в данном случае является положительным обстоятельством, поскольку неограничен нашими сегодняшними представлениями о природе и генном контроле конкретного интересующего нас признака.
Кроме всего этого группа ученых, таких как Марк Адам (ведущий сотрудник института геномных исследований в штате Мэриленд – США, частной исследовательской компании, занимающейся исключительной работой в области картирования генов), Крэйк Вентер (директор этого института) и соавторами, разрабатывается проект «Геном человека». Цель этого проекта заключается в выяснении последовательности оснований во всех молекулах ДНК в клетках человека. Одновременно должна быть установлена локализация всех генов, что помогло бы выяснить причину многих наследственных заболеваний и этим открыть пути к их лечению. Что бы последовательно приближаться к решению проблемы картирование генов человека, было сформулировано пять основных целей:
- завершить составление детальной генетической карты, на которой были бы помечены гены, отстоящие друг от друга на расстоянии не превышающем в среднем 2 млн. оснований (1 млн. оснований принято называть мегобазой);
- составить физические карты каждой хромосомы (разрешение 0.1 Мб);
- получить карту всего генома в виде охарактеризованных клонов (5 тыс. оснований в клоне или 5 Кб);
- завершить к 2004 году полное секвенирование ДНК (разрешение одного основание);
- нанести на полностью завершенную секвенсовую карту все гены человека (к 2005 году).
Ожидалось, что, когда все указанные цели будут постигнуты, исследователи определят все функции генов и разработают методы биологического и медицинского применения полученных данных.
Рассмотрев темпы ускорения работы в рамках проекта «Геном человека», руководители этого проекта объявили 23 октября 1998г., что программа будет полностью завершена гораздо раньше, чем планировалось, и сформулировали «Новые задачи проекта «Геном человека»:
- полностью завершить в декабре 1998 года работу по секвенирование генома «Круглого червя» c. Elegans (это было сделано в срок);
- закончить предварительный анализ последовательности ДНК человека к 2001 году, а полную последовательность к 2003 году;
- картировать к 2002 году геном плодовой мухи;
- начать секвенирование генома мыши с использованием методов ДНК искусственных хромосом дрожжей (завершить этот проект к 2005 году).
2.3.1. Что будет сделано после завершения анализа генома человека
Главная стратегическая задача будущего сформулирована следующим образом: изучить однонуклеотидные вариации ДНК в разных органах и клетках отдельных индивидуумов и выявить различия между индивидуумами. Анализ таких вариаций даст возможность не только подойти к созданию индивидуальных генных портретов людей, что в частности даст возможность лечить болезни, но и определить различия между популяциями. А также выявлять географические районы повышенного риска, что поможет давать чёткие рекомендации о необходимости очистки территории от загрязнения и выявить производства, на которых есть большая опасность поражение геномов персонала.
Эта грандиозная задача рождает не одни радужные ожидания всеобщего блага, но и вполне осознанную тревогу юристов и борцов за индивидуальные права человека. Так, в частности, высказываются возражения против распространения персональной информации без решения тех, кого она касается. Один пример помогает понять эти тревоги: уже сейчас страховые компании нацелились на добывание таких сведений правдами и неправдами, они намериваются использовать данные против тех, кого они страхуют. Например, если подающий на страховку несёт потенциально болезнетворный ген, компании не хотят страховать таких людей вовсе или же пытаются заломить бешенные суммы за их страховки. Исходя из этого, конгресс США уже принял ряд законов, направленный на строгий запрет распространения генетической информации относительно отдельных людей, юристы всего мира интенсивно работают в данном направлении.
3. Предпосылки формирования генной инженерии
3.1. Открытие двойной структуры ДНК и матричного синтеза
Начальные работы американских учёных Уотсона и Крика были произведены в 1953 году. Они дали возможность развиваться генной инженерии в качестве самостоятельного раздела науки. Эти открытия заключены в следующем:
Была открыта двойная структура ДНК и постулирован её матричный синтез. Двойная спираль ДНК при репликации разделится и вдоль нити ДНК, специальные ферменты-полимеры, собирают точные копии материнской ДНК, таким образом в клетке перед делением две совершенно одинаковые молекулы ДНК, одна из которых после деления клетки попадает в дочернюю клетку. Таким образом дочерняя клетка несет ту же самую информацию, что и материнская, следовательно выполняет те же самые функции. Итак, в клетках живого организма возможен особый тип реакции – матричный синтез. Одна молекула – матрица, а вторая строится по её программе. Репликация ДНК, синтез всех видов РНК и сборка молекул белка, в соответствии со структурой иРНК – это все варианты матричного синтеза, который происходит всегда при участии нуклеиновых кислот.
По тому же самому механизму осуществляется сборка РНК, только не двух спиралей, а одной. Этот процесс получил название – транскрипция. Поток информации в клетке обеспечивает реакции матричного синтеза: репликация ДНК (необходима для передачи наследственной информации дочерним клеткам), транскрипция (синтез иРНК в ядре клетки) и трансляция (сборка белковой цепи на иРНК при помощи рибосомы).
Казалось бы, что на рубеже 70-х годов молекулярная биология достигла определённой степени завершенности: были установлены структура и механизм репликации ДНК, провозглашена «центральная догма» экспрессии гена (транскрипция и трансляция), выявлены основные аспекты регуляции активности гена. В этот период главным объектом молекулярно-генетических исследований были микроорганизмы. Переход к эукариотам (включая человека) встретился с дополнительными проблемами и трудностями, и кроме того, существовавшие в то время методы не позволяли рассчитывать на получение принципиально новых результатов. Стремительный порыв в развитии молекулярной генетики в начале 70-х годов стал благодаря появлению нового экспериментального инструмента – рестриктационных эндонуклеаз. Был открыт путь для широкомасштабного получения генных продуктов (физически значимых белков) и для генетического манипулирования с различными организмами.
В прошлом генетика и медицинская генетика развивалась как относительно независимые отрасли науки, теперь многие из их разделов оказались вовлечённые в общее русло молекулярно-генетических исследований, и провести между ними грань – трудно.
Сейчас, множество ученых заняты различными работами связанные с проблемами генной инженерии – это и методы, основанные на использовании рестриктационных ферментов, анализ гена человека, методы гибридизации нуклеиновых кислот, секвенирование ДНК, сортировки хромосом при помощи цитофиурометрии и многое, многое другое.
3.2. Принципы технологий рекомбинантных ДНК
Было выделено много рестриктаз (более 150), расщепляющих ДНК в специфических сайтах. Например эндонуклеаза R1 регистрирует двухцепочную ДНК по двум сайтам таким образом, что образуются два липких конца:
¯
G-A-A-T-T-C
||| || || || || |||
C-T-T-A-A-G
Липкие концы различных молекул ДНК, расщеплённых этим ферментом, могут вступать по четырём –A-T-парам. Рестриктационные эндонуклеазы различаются по тем сайтам ДНК, которые они распознают и разрезают. Их можно использовать для различных целей. Однако наиболее распространенным этапом является их применение для амплификации специфического определения нуклеотидных последовательностей фрагментов ДНК, необходимых для ДНК
или для изучения механизмов экспрессии генов. Последняя проблема наиболее важна в практическом аспекте: гены контролирующие образование функционально активных белков теперь можно вводить в бактерии и размножать (амплифицировать). Эта процедура называется клонированием генов. Благодаря ей появилась возможность вырабатывать в больших количествах белки, которые раньше удавалось получить ничтожно мало. Эта технология основана на следующем принципе: помимо своей собственной кольцевой хромосомы, бактерии часто содержат дополнительные маленькие кольцевидные молекулы двух цепочной ДНК, называемые плазмидами.
Плазмиды реплицируются автономо и сами могут содержать гены, определяющие устойчивость бактерий к антибиотикам или контролирующие синтез веществ, например: колицинов, убивающих другие бактерии (рис.1).
Плазмидную ДНК можно выделить и расщепить подходящей рестриктазой только в одном сайте, превратив кольцевую молекулу в линейную с липкими концами.

Фрагменты любой чужеродной ДНК с такими же липкими концами (полученными после разрезания аналогичной рестриктазой) можно сшить с плазмидой ДНК с помощью лигазы.
Рис. 1. Клетка E-coli с хромосомой и плазмидой.
Рекомбинантную конструкцию вводят затем в бактерию, где она реплицируется ( рис.2 )Источник экзогенной ДНК не имеет значения. ДНК может быть получена, например, из клеток человека, но можно сшивать и искусственно синтезированные гены. Кроме бактериальных плазмид в качестве векторов (носителей) ДНК используют фаги λ (объект исследования Альберта). Часть генома этого фага не обязательна для его размножения в бактерии. Вместо него можно ввести чужеродную ДНК, которая будет размножаться вместе с фаговой после инфицирования бактерий.
Добиться репликации и амплификации в составе плазмидной (или фаговой) ДНК после трансформации бактериальной клетки ещё не значит решить все её проблемы. Прежде всего возникают два вопроса:
1. Как распознавать клоны, содержащие гибридную ДНК, среди потомства трансформированных клеток или живых бактериофагов ?
2. как идентифицировать необходимые фрагменты ДНК среди многих клонированных неизвестных фрагментов?
Например можно отбирать бактериальные клетки, если они несут плазмиду с фактором устойчивости к антибиотику, выращивая их на среде, на среде, содержащей антибиотик. Нетрансформированные клетки без плазмид(и, следовательно, без гена устойчивости к антибиотику) просто не будут расти на такой среде. В последнее время разработано много специальных методов вакцинации, которые позволяют отбирать только рекомбинантные клетки.
Для генной инженерии белков недостаточно отобрать и размножить определённые фрагменты ДНК, необходимо ещё индуцировать их экспрессию в клетке. Для этого необходимо «подключить» рекомбинантную молекулу ДНК , последующую трансляцию матричной РНК и процессинг как на транскрипционном , так и на трансляционных уровнях.
3.3. Идентификация и анализ генов
Ещё одна область применения рестриктаз – идентификация и определение числа генов. Эти задачи решаются с помощью метода разработанного Саузерном.
Тотальную ДНК из клеток человека гидролизуют эндонуклеазой примерно на 500000 фрагментов длиной от 102
до 105
нуклеотидных пар. Затем фрагменты разделяют по молекулярной массе с помощью гель- электрофореза в ага розе, после чего ДНК денатурирует с щелочью прямо в геле, чтобы получить одноцепочные фрагменты. Их переносят на нитроцеллюлозный фильтр и фиксируют высушиванием при 800
С. В результате получается отпечаток(реплика) картины разделения фрагментов ДНК по их размеру. Эти фрагменты можно идентифицировать методом гибридизации с радиоактивными ДНК-зондами, специфичными для определённых генов или хромосом. Любой фрагмент, содержащий всю последовательность зондируемого гена или его часть, будет выглядеть на радиоавтографе в виде тёмной полосы.
Зонды и генные библиотеки. Главное условие такого анализа - наличие подходящего геноспецифического радиоактивного ДНК-зонда, который можно использовать для гибридизации.
3
.4. Сортировка хромосом
Следующий метод – это метод сортировки хромосом при помощи цитофлюрометрии. Этот метод может быть использован в двух разных целях:
1) Для идентификации и количественного анализа большого числа хромосом в течение очень короткого времени.
2) Для препаративного разделения хромосом. Этот метод имеет два преимущества перед стандартными методами анализа хромосом:
- во-первых, он полностью автоматический, благодаря чему исключается элемент субъективности
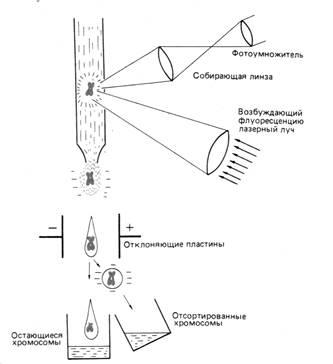
- во-вторых, он намного быстрее (рис. 3)
Однако важнее, что этот метод позволяет препаративно разделять хромосомы, и при наличии специфических зондов исследовать структуру и функцию отдельных генов становится относительно просто. В этом случае ген можно локализовать в хромосоме с помощью гибридизации in situ, размножить его ДНК путём клонирования и секвенирования.
Рис. 3 Принцип сортировки хромосом с помощью лазера. Хромосомы окрашены флуоресцирующим красителем. Флуоресценция возбуждается лазерным лучом и измеряется для каждой хромосомы отдельно. Данные измерений используют для сортировки хромосом.
3.5. Секвенирование ДНК
Последовательность нуклеотидов и генетический код.
Методы определённой последовательности аминокислот в полипептидной цепи были известны ещё в 50-х годах. Теоретически это относительно лёгкая проблема, поскольку все 20 аминокислот, встречающихся в природных белках, имеют разные свойства. С другой стороны, нуклеотидная последовательность ДНК относительно однородна по составу однородных звеньев, так как содержит только четыре типа азотистых оснований – гуанин, цитозин, аденин и тимин. Когда в 60-е годы был расшифрован генетический код, появилась возможность востанавливать (дедуцировать) нуклеотидную последовательность соответствующего белка. Однако генетический код является вырожденным, то есть одной и той же аминокислоте соответствуют несколько нуклеотидных триплетов. Следовательно сведения о нуклеотидной последовательности аминокислот в белке, не однозначны. Кроме того последовательности аминокислот не содержат никакой информации о последовательности некодирующих участков ДНК. Принцип состоит в следующем: длинную молекулу ДНК фрагментируют при помощи агентов, расщепляющихся в специфических сайтах. Затем определяют последовательность нуклеотидов в каждом из этих фрагментов. Очерёдность фрагментов в целой молекуле восстанавливают, используя перекрывающие концы: идентичные цепи разрезают повторно другой рестриктазой, а затем последовательности, перекрывающихся образующихся при обработке двумя рестриктазами разной специфичности, сравнивают. Так может быть реконструирована полная последовательность. В пределах отдельных фрагментов порядок нуклеотидов определяют с помощью специальных методов. Раньше секвенирование ДНК было весьма трудным делом, теперь же оно осуществляется очень легко и быстро.
3.6. Динамичность генома
Методы новой гентики расширили наши знания о структуре генетического материала. В 1963 году Тэйлор описал “индуцированные фагом мутации E. Coli”, вкоре после этого, Старлингер и Седлер описали IS-элементов у бактерий. Эти элементы получили название мобильных, теперь же они определяются как специфические последовательности ДНК, которые могут неоднократно внедряться в разные сайты генома. Перенос генов от одной бактерии к другой с помощью фага (трансдукция) известен давно, а теперь используется и в генетической инженерии эукариот (включая клетки млекопитающих). Возможно, такие процессы могут происходить и в природе. Более того, последовательности ДНК, гомологичные глобиновому гену человека, были обнаружены у бобовых растений. Функция такого гена у растений может состоять в том, чтобы “обеспечить кислородом клубеньковые бактерии в ткани”. Наличие этого гена может быть объяснено переносом его от насекомых или млекопитающих.
4. Области практического применения генной инженерии
4.1. Создание трансгенных растений
Еще 10 лет тому назад биотехнология растений заметно отставала в своем развитии, но за последние 3 года наблюдается быстрый выброс на рынок трансгенных растений с новыми полезными признаками. Трансгенные растения в США в 1996 году занимали площадь 3 млн. акров, в 1997 году площадь увеличилась до 15 млн. акров, в 1998 году – до 60 млн. акров, а в прошлом году до 80 млн. акров. Поскольку основные трансгенные формы кукурузы, сои, хлопчатника с устойчивостью к гербицидам и насекомым хорошо себя зарекомендовали, есть все основания ожидать, что площадь под генноиженерные растения в будущем (2001 году) увеличатся в 4-5 раз.
В апреле 1998 года доля в процентах трансгенных форм растений в сельском хозяйстве составило:
- кукуруза – 6
- соя – 12
- хлопчатник – 15
- томаты – <1
Так как число жителей за последнее столетие увеличилось с 1.5 до 5.5 млрд. человек, а к 2020 году предполагается вырост до 8 млрд., таким образом возникает огромная проблема, стоящая перед человечеством. Эта проблема заключается в огромном увеличение производства продуктов питания, несмотря на то, что за последние 40 лет производство увеличилось в 2.5 раза, все равно этого не достаточно. И в мире в связи с этим наблюдается социальный застой, который становится все более настоятельным. Другая проблема возникла с медицинским лечением. Несмотря на огромные достижение современной медицины, производимые сегодня лекарственные препараты столь дороги, что ¾ населения земли сейчас полностью полагаются на традиционные донаучные методы лечения, прежде всего на неочищенные препараты растительного происхождения.
В развитых странах лекарственные средства на 25% состоят из природных веществ, выделенных из растений. Открытия последних лет (противоопухолевые препараты: таксол, подофиллотоксин) свидетельствуют о том, что растения еще долго будут оставаться источником полезных биологически-активных веществ (БТА), и что способности растительной клетки к синтезу сложных БТА все еще значительно превосходят синтетические способности инженера-химика. Вот почему ученые взялись за проблему создания трансгенных растений.
Отсчёт истории генетической инженерии растений принято вести с 1982 года, когда впервые были получены генетически трансформированные растения. Метод трансформации основывается на природной способности бактерий
Agrobacteriumtumefaciens генетически модифицировать растения. Реконструированные штаммы Agrobactrium, содержащие неонкогенные варианты Ti-плазмид и обладающие повышенной вирулентностью, стали основой одного из наболее популярных методов трансформации. Первоначально трансформация применялась для генно-инженерных двудольных растений, однако работы последних лет свидетельствуют, что этот метод эффективен и в отношении кукурузы, риса, пшеницы. Другим широко распространённым методом трансформации, является технология, основанная на обстреле ткани микрочастицами золота (или других тяжелых металлов), покрытыми раствором ДНК. Все выращиваемые ныне коммерческие сорта получены с помощью названных выше двух методов.
Современный арсенал методов трансформации, однако, довольно обширен и включает такие подходы, как введение ДНК в голые клетки (протопласты), электропорация клеток, микроинъекций ДНК в клетки, прокалывание клеток путём встряхивания их в суспензии микроигл, опосредованная вирусами инфекции и так далее.
Генетические изменённые растения с устойчивостью к различным классам гербицидов в настоящее время являются наиболее успешным биотехнологическим продуктом. Дело в том, что биотехнология позволила совершить такой прыжок, так как оказалось возможным генетически изменять устойчивость растений к тем или иным гербицидам либо путем введения генов, кодирующих белки, нечувствительные к данному классу гербицидов, либо за счет введения генов, обеспечивающих ускоренный метаболизм гербицидов растений. К настоящему времени клонированы гены, кодирующие нечувствительные к действию гербицидов ферменты-мишени, что дало возможность получать трансгенные растения, устойчивые к таким гербицидам, как глифостат и хлорсульфуроновым, и имидазолиноновым гербицидом. Изолированы также гены, которые кодируют ферменты деградации некоторых гербицидов, что позволило получить трансгенные растения устойчивые к фосфинотрицину и далапону. В 1997 году устойчивая к Roundup соя, распространяемая компанией "AsGrow", была признана в США сельскохозяйственным продуктом года.
Ученые пошли далее. Так как множество растений подвержены нападению и поеданию со стороны насекомых, то ученые генной инженерии провели эксперимент с давно известной бактерией Bacillus-Thiringiensis, которая продуцирует белок, оказалось она является очень токсичной для многих видов насекомых, но в то же время безопасна для млекопитающих., белок (дельта-эндотаксин, CRY-белок) продуцируется различными штамами Bacillus-Thiringiensis. Это прототаксин который расщепляется в кишечнике насекомых, образуя активированный токсин. Активизированный белок специфично связывается с рецепторами средней кешки насекомых, что приводит к образованию пор и лизису клеток кишечного эпителия. Взаимодействие токсинов с рецепторами строго специфично, что усложняет подбор комбинации токсин-насекомое. В природе найдено большое количество штаммов Bacillus-Thiringiensis, чьи токсины действуют только на определенные виды насекомых. Препараты Bacillus-Thiringiensis в течение десятилетий использовались для контроля насекомых на полях.
Встраивание гена этого белка в геном растений дает возможность получить трансгенные растения, не поедаемые насекомые. Но этот метод потребовал большой работы со стороны генной инженерии, в плане подборов необходимых штаммов и созданию генно-инженерных конструкций, которые дают наибольший эффект для конкретных классов насекомых. Кроме видоспецифичности по действию на насекомых встраивание прокариотических генов дельта-токсинов в геном растений даже под контролем сильных эукариотических промоторов не привело к высокому уровню экспрессии. Предположительно такое явление возникло в связи с тем, что эти бактериальные гены содержат значительно больше адениновых и тиминовых нуклеатидных оснований, чем растительная ДНК. Эта проблдема была решена путем создания модефицированных генов, где один из природного гена вырезали и добавили те или иные фрагменты с сохранением доменов, кодирующих активные части дельта-токсинов. Так, например, с помощью таких подходов был получен картофель, устойчивый к колорадскому жуку. В настоящее время так называемый Bt – растения хлопка и кукурузы занимают основную долю в общем объеме генетически модифицированных растений этих культур, которые выращивают на полях США.
4.2. Генные вакцины
4.2.1. Актуальность разработки новых вакцин
Вакцины — одно из самых значительных достижений медицины, их использование к тому же чрезвычайно эффективно с экономической точки зрения. В последние годы разработке вакцин стали уделять особое внимание. Это обусловлено тем, что до настоящего времени не удалось получить высокоэффективные вакцины для предупреждения многих распространенных или опасных инфекционных заболеваний. По данным созданной в прошлом году международной организации «Всемирный союз по вакцинам и иммунизации» (в числе ее участников — ВОЗ, ЮНИСЕФ, Международная федерация ассоциаций производителей фармацевтической продукции, Программа Билла и Мелинды Гейтс по вакцинации детей, Рокфеллеровский фонд и др.), в настоящее время отсутствуют эффективные вакцины, способные предупредить развитие СПИДа, туберкулеза и малярии, от которых в 1998 г. умерло около 5 млн человек. Кроме того, увеличилась заболеваемость, обусловленная теми инфекциями, с которыми человечество ранее успешно боролось. Этому способствовало появление лекарственно-устойчивых форм микроорганизмов, увеличение числа ВИЧ-инфицированных пациентов с иммунной недостаточностью, ослабление систем здравоохранения в странах с переходной экономикой, увеличение миграции населения, региональные конфликты и др. При этом распространение микроорганизмов, устойчивых к воздействию антибактериальных препаратов, приобрело характер экологической катастрофы и поставило под угрозу эффективность лечения многих тяжелых заболеваний. Повышенный интерес к вакцинам возник после того, как была установлена роль патогенных микроорганизмов в развитии тех заболеваний, которые ранее не считали инфекционными. Например, гастриты, пептическая язва желудка и двенадцатиперстной кишки, ассоциированная с H. pylori, злокачественные новообразования печени (вирусы гепатита В и С).
Поэтому в последние 10–15 лет правительства многих стран стали принимать меры, направленные на интенсивную разработку и производство принципиально новых вакцин. Например, в США в 1986 г. был принят закон («National Vaccine Injury Compensation Act»), защищающий производителей вакцин от юридической ответственности при подаче судебных исков, связанных с развитием побочных реакций при вакцинации, если они не были обусловлены ошибками при производстве вакцины. С изменением ситуации увеличился и мировой рынок вакцин, объем продаж которого в 1998 г. составил 4 млрд долларов США в стоимостном выражении. Однако многие считают, что в ближайшие годы этот сектор фармацевтической промышленности будет развиваться гораздо быстрее. Так, согласно публикациям в американском журнале «Signals Magazine» (январь 1999 г.), который освещает ситуацию в современной биотехнологической промышленности, объем продаж вакцин на мировом рынке через 10 лет составит 20 млрд долларов США. Этот прогноз принадлежит М. Греко, исполнительному директору компании «Merieux MSD», совместного предприятия крупнейших производителей вакцин — компаний «Pasteur Merieux Connaught» (теперь «Aventis Pasteur») и «Merck & Co.».
4.2.2. Разработка ДНК – вакцин
Используемые сегодня вакцины можно разделить в зависимости от методов их получения на следующие типы:
• живые аттенуированные вакцины;
• инактивированные вакцины;
• вакцины, содержащие очищенные компоненты микроорганизмов (протеины или полисахариды);
• рекомбинантные вакцины, содержащие компоненты микроорганизмов, полученные методом генной инженерии.
Технологию рекомбинантной ДНК применяют также для создания живых ослабленных вакцин нового типа, достигая аттенуации путем направленных мутаций генов, кодирующих вирулентные протеины возбудителя заболевания. Эту же технологию используют и для получения живых рекомбинантных вакцин, встраивая гены, кодирующие иммуногенные протеины, в живые непатогенные вирусы или бактерии (векторы), которые и вводят человеку.

Рис. 4. Одноразовый генный пистолет компании «Powderject»
а — внешний вид; б — в разрезе
В 1990 г. в некоторых исследовательских лабораториях приступили к разработке новых вакцин, которые основаны на введении «голой» молекулы ДНК. Уже в 1992–1993 гг. несколько независимых групп исследователей в результате эксперимента доказали, что введение чужеродной ДНК в организм животного способствует формированию иммунитета.
Принцип применения ДНК-вакцин заключается в том, что в организм пациента вводят молекулу ДНК, содержащую гены, кодирующие иммуногенные белки патогенного микроорганизма. ДНК-вакцины называют еще генными, генетическими, полинуклеотидными вакцинами, вакцинами из нуклеиновых кислот. На совещании специалистов по генным вакцинам, проведенном в 1994 г. под эгидой ВОЗ, было решено отдать предпочтение термину «вакцины из нуклеиновых кислот» с их подразделением соответственно на ДНК- и РНК-вакцины. Такое решение основывалось на том, что употребление термина «ДНК-вакцина» не сформирует ошибочное мнение о том, что новые вакцины вносят изменения в генетические структуры организма вакцинируемого человека. Тем не менее, многие специалисты считают более точным термин «генные вакцины» (поскольку иммунная реакция направлена не против ДНК, а против антигенного белка, кодируемого геном), который также часто применяют.
Для получения ДНК-вакцин ген, кодирующий продукцию иммуногенного протеина какого-либо микроорганизма, встраивают в бактериальную плазмиду. Плазмида представляет собой небольшую стабильную молекулу кольцевой двухцепочечной ДНК, которая способна к репликации (воспроизведению) в бактериальной клетке. Кроме гена, кодирующего вакцинирующий протеин, в плазмиду встраивают генетические элементы, которые необходимы для экспрессии («включения») этого гена в клетках эукариотов, в том числе человека, для обеспечения синтеза белка. Такую плазмиду вводят в культуру бактериальных клеток, чтобы получить большое количество копий. Затем плазмидную ДНК выделяют из бактерий, очищают от других молекул ДНК и примесей. Очищенная молекула ДНК и служит вакциной. Введение ДНК-вакцины обеспечивает синтез чужеродных протеинов клетками вакцинируемого организма, что приводит к последующей выработке иммунитета против соответствующего возбудителя. При этом плазмиды, содержащие соответствующий ген, не встраиваются в ДНК хромосом человека.
 ДНК-вакцины можно вводить в солевом растворе обычным парентеральным способом (внутримышечно, внутрикожно). При этом бoльшая часть ДНК поступает в межклеточное пространство и только после этого включается в клетки. Применяют и другой метод введения, используя так называемый генный пистолет (рис. 5, 6). Для этого ДНК фиксируют на микроскопических золотых гранулах (около 1–2 мкм), затем с помощью устройства, приводимого в действие сжатым гелием, гранулы «выстреливают» непосредственно внутрь клеток. Следует отметить, что аналогичный принцип введения лекарства с помощью струи сжатого гелия используют и для разработки новых способов доставки лекарственных средств (с этой целью оптимизируют размеры частиц лекарственного вещества и их плотность для достижения необходимой глубины проникновения в соответствующую ткань организма). Этот метод требует очень небольшого количества ДНК для иммунизации. Если при иммунизации классическими субъединичными вакцинами вводят микрограммы протеина, то при использовании ДНК-вакцины — нанограммы и даже меньше. Говоря о минимальном количестве ДНК, достаточном для индукции иммунного ответа, С.А. Джонстон, директор Центра биомедицинских изобретений Техасского университета, в журнале «The Scientist» (1998) отмечает, что с помощью генного пистолета можно однократно ввести мыши «фактически 27 тыс. различных плазмид и получить иммунный ответ на индивидуальную плазмиду». ДНК-вакцины можно вводить в солевом растворе обычным парентеральным способом (внутримышечно, внутрикожно). При этом бoльшая часть ДНК поступает в межклеточное пространство и только после этого включается в клетки. Применяют и другой метод введения, используя так называемый генный пистолет (рис. 5, 6). Для этого ДНК фиксируют на микроскопических золотых гранулах (около 1–2 мкм), затем с помощью устройства, приводимого в действие сжатым гелием, гранулы «выстреливают» непосредственно внутрь клеток. Следует отметить, что аналогичный принцип введения лекарства с помощью струи сжатого гелия используют и для разработки новых способов доставки лекарственных средств (с этой целью оптимизируют размеры частиц лекарственного вещества и их плотность для достижения необходимой глубины проникновения в соответствующую ткань организма). Этот метод требует очень небольшого количества ДНК для иммунизации. Если при иммунизации классическими субъединичными вакцинами вводят микрограммы протеина, то при использовании ДНК-вакцины — нанограммы и даже меньше. Говоря о минимальном количестве ДНК, достаточном для индукции иммунного ответа, С.А. Джонстон, директор Центра биомедицинских изобретений Техасского университета, в журнале «The Scientist» (1998) отмечает, что с помощью генного пистолета можно однократно ввести мыши «фактически 27 тыс. различных плазмид и получить иммунный ответ на индивидуальную плазмиду».
Рис. 5. Многоразовый генный пистолет
компании «Powderject»
а — сменный картридж
б — прибор в полной сборке
Последующие эксперименты подтвердили способность ДНК-вакцин формировать иммунитет в отношении разнообразных возбудителей.
4.3. Генотерапия
Технологии генодиагностики и генотерапии базируются на мировых достижениях в расшифровке генома человека. Технологии генодиагностики включают разработку приемов точной локализации генов в геноме человека, ответственных за наследственные и соматические заболевания, а также методологии пренатальной и доклинической диагностики. Их важной составляющей является сравнительный анализ структуры генома в норме и патологии.
Среди технологий генотерапии в настоящее время актуальны следующие: генотерапия соматических клеток, генотерапия репродуктивных (половых) клеток, генотерапия с использованием рибозимов и антисенс-ДНК.
Генотерапия и генодиагностика - это перспективные технологии фундаментальной и прикладной биомедицины, направленные на лечение и профилактику наследственных (генетических) и приобретенных заболеваний, в том числе онкологических.
В основе генотерапии, развивающейся на базе и в комплексе с генодиагностикой, лежит контролируемое изменение генетического материала клеток, приводящее к "исправлению" не только наследственных, но и, как стало ясно в последнее время, приобретенных генетических дефектов живого организма.
Важнейшей технологической задачей генотерапии является разработка системы переноса или адресной доставки корректирующего генетического материала к клеткам-мишеням в организме больного, несущего в своем геноме дефектный ген. Предлагаемые технологии характеризуются точностью выявления гена, ответственного за генетический дефект и выбора системы переноса корректирующих генов, адресностью доставки в организм больного генетического материала, исправляющего генетический дефект.
Технологии генодиагностики и генотерапии применяются в следующих отраслях:
здравоохранение (развитие методологии генодиагностики и, в частности, системы пренатальной генодиагностики, будет способствовать своевременному выявлению генетических болезней и принятию соответствующих профилактических мер; генотерапия может быть использована для лечения болезней, связанных с мутациями генома (в том числе серповидно-клеточной анемии, эмфиземы, гемофилии и др.), инфекционных заболеваний; для коррекции дефектов центральной нервной системы и для стимуляции иммунного ответа организма при онкозаболеваниях);
сельское хозяйство (технологии генодиагностики и генотерапии могут быть применены в ветеринарии и фитопатологии).
Технологии генодиагностики и генотерапии являются инструментом реализации новой медико-биологической стратегии, конечная цель которой - избавление человечества от генетических и приобретенных болезней. Актуальность генотерапии для человека связана с тем, что более 5000 наследственных и приобретенных заболеваний связано с генетическими дефектами. Генотерапия может использоваться не только для лечения, но и для профилактики наследственных и приобретенных заболеваний. Таким образом, данная технология имеет большое социальное и народнохозяйственное значение.
За рубежом генодиагностика и генотерапия рассматриваются как один из приоритетов развития биомедицины. К настоящему времени одобрено более 7 протоколов по генотерапии, в которых предложены способы лечения наследственных заболеваний. Такие протоколы разрабатываются в Китае, Франции, Великобритании, Италии, Нидерландах и ряде других стран. В США Национальным Комитетом по рекомбинантным ДНК (RAC) одобрено 18 клинических испытаний с использованием генотерапии, начато лечение одного из видов рака кожи - меланомы.
В Российской Федерации также освоены основные технологии генотерапии - секвенирование, физическое и генетическое картирование генома человека и животных, осуществляется расшифровка молекулярных механизмов наследственных и онкозаболеваний, решаются проблемы генетической безопасности человека, сохранения его генофонда в условиях разрушающего антропогенного воздействия среды. Вместе с тем, для достижения зарубежного уровня в этой области России необходимо принять срочные меры по увеличению финансирования НИОКР и по усилению приборного обеспечения. Необходимым условием развития предлагаемых технологий в стране является организация международной кооперации.
Генную терапию на современном этапе можно определить как лечение наследственных, мультифакториальных и наследственных (инфекционных) заболеваний. Путем введения в клетки пациентов с целью направленного изменения генных дефектов или придания клеткам новых функций. Первые клинические испытания методов генетической терапии были предприняты 22 мая 1989 года с целью генетического маркирования опухаль-инфильтрующих лимфоцитов в случае прогрессирующей меланомы первым моногенным наследственным заболеванием, в отношении которого были применены методы генетической терапии, оказался наследственный иммунодефицит, обусловленный мутацией в гене аденозиндезоминазы (АДА). 14 сентября 1990 года в Бетесде (США) 4-летней девочке, страдающей этим достаточно редким заболеванием (1:100000), были пересажены ее собственные лимфоциты, предварительно трансформированные в не организма (exvivo) геном АДА (ген АДА + ген neo + ретровирусный вектор). Лечебный эффект наблюдается в течение нескольких месяцев, после чего процедура была повторена с интервалом 3-5 месяцев. За 3 года терапии проведены 23 внутривенные инъекции. В результате лечения, состояния пациентки настолько улучшилось , что она смогла вести нормальный образ жизни и не бояться случайных инфекций. Сейчас эти испытания проводятся в Италии, Франции, Великобритании и Японии.
4.4. Перспективы клонирования животных.
Идея клонирования животных, т.е. получение генетически идентичных копий, родилась благодаря успешным экспериментам по пересадке ядер дифференцированных клеток в энуклеированные яйцеклетки или ооциты, выполненным на амфибиях. Цель этих экспериментов была сугубо теоретическая - выяснить вопрос, способно ли ядро (геном) дифференцированной клетки к репрограммированию и восстановлению тотипотентности, т.е., будучи помещенным в цитоплазму яйца, способно ли оно обеспечить полное развитие подобно оплодотворенной яйцеклетке. Фактически речь шла о возможности обратимости эмбриональной дифференцировки и выяснению вопроса: претерпевает ли геном в процессе развития необратимые изменения или модификации? Успешные опыты J.Gurdon и его сотрудников, показавшие возможность развития взрослых амфибий из реконструированных яйцеклеток после трансплантации в них ядер из клеток эпителия кишечника плавающей личинки (головастика), были интерпретированы как убедительное доказательство, что геном дифференцированных клеток способен к репрограммированию в цитоплазме яйцеклетки и восстановлению тотипотентности, подобно оплодотворенному яйцу. Из этих результатов логично вытекало, что используя технику трансплантации ядер из соматических клеток взрослых особей в энуклеированные яйца или ооциты, можно получать генетические копии животного, служившего донором ядер дифференцированных клеток. Безусловно, клонирование животных открывало бы заманчивые перспективы для генетического копирования животных, прежде всего сельскохозяйственных, тех, которые имеют те или иные выдающиеся показатели продуктивности.
Однако первые попытки применить описанный выше подход для клонирования млекопитающих были безуспешными и даже скандальными. Сенсационные результаты Illmensee по рождению мышей, развившихся после пересадки кариопластов из разных частей предимплантационных эмбрионов мыши в энуклеированные яйца, не были подтверждены другими исследователями. Эти результаты вызвали еще большие сомнения после заявления лаборанта Illmensee, что результаты опытов Illmensee были фальсифицированы. В начале 80-х годов эксперименты по трансплантации ядер дифференцированных клеток в энуклеированные яйца или ооциты показали, что у мышей тотипотентность утрачивается после 2-го деления-дробления. Другой экспериментальный подход для изучения тотипотентности эмбрионов был основан на разделении бластомеров на ранних стадиях развития (до 16-клеточной стадии) и независимой их трансплантации приемным матерям. Результаты этих экспериментов показали, что у мышей тотипотентность утрачивается после 4-клеточ-ной стадии, хотя у овец такая потеря происходит на более поздней стадии развития (после 16-клеточной стадии). Открытие импринтинга и его существенной роли в развитии млекопитающих сделало еще более проблематичной возможность клонирования млекопитающих, поскольку выяснилось, что материнский и отцовский геномы имеют разный вклад в нормальное развитие эмбриона, причем эти функциональные различия родительских геномов формируются в процессе овогенеза и сперматогенеза, импринтируются и реализуются в течение всего онтогенеза.
Тем не менее, исследования тотипотентности и плюрипотентности в эмбриональном развитии продолжались с использованием новых экспериментальных подходов. Уже в конце 80-х годов стало очевидным, что ооцит на стадии М2 (второе меойтическое деление) обладает факторами, способными репрограммировать геном клеток, полученных из внутренней клеточной массы бластоцисты после их трансплантации в энуклеированный ооцит М2. Здесь следует отметить значительный вклад в разработку этой техники шотландской группы исследователей под руководством Ian Wilmut и американских исследователей Keefer, Mathews и First. В 1996 г. вышли две публикации по успешному рождению ягнят и развитию эмбрионов коров до 80-85 дней беременности в экспериментах по трансплантации кариопластов, полученных из клеток культуры эмбриональных стволовых клеток, в энуклеированные ооциты М2 (Campbell et al., 1996; Keefer et al., 1996). По-видимому, эти успехи подтолкнули Ian Wilmut и его коллег попытаться использовать в качестве доноров ядра (кариопласты) дифференцированных клеток, взятых от эмбрионов или взрослых животных. Результатом этих экспериментов явилось рождение Dolly, овцы, развившейся из ооцита М2, у которого было заменено собственное ядро на ядро клетки из культуры эпителиальных клеток, полученной из молочной железы взрослой лактирующей овцы. Вне всяких сомнений, это выдаюшийся успех в клонировании животных. Впервые предложен, хотя и сложный в техническом отношении, способ получения генетических копий млекопитающих. Несомненно также и то, что клонирование станет сильнейшим стимулом для развития нового направления в биотехнологии животных и откроет широкие возможности в селекции сельскохозяйственных животных.
В теоретическом плане работа группы Ian Wilmut показала, что в процессе развития геном не претерпевает каких-либо необратимых изменений, по крайней мере, в части высокодифференцированных соматических клеток. Более того, возможно репрограммирование генома соматических клеток путем трансплантации его в цитоплазму ооцита М2. Фактически это исследование заложило теоретическую базу клонирования животных, хотя здесь имеется много неясностей и нужны дополнительные экспериментальные исследования.
Уместно в этом контексте упомянуть недавние результаты, полученные в лаборатории генетических основ онтогенеза Института цитологии и генетики СО РАН. Здесь были впервые получены гибридные клетки путем слияния высокоплюрипотентных стволовых эмбриональных клеток с клетками селезенки взрослой мыши. Некоторые клоны гибридных клеток имели нормальный диплоидный набор хромосом и дополнительную Х-хромосому, происходящую из высокодифференцированной клетки. В экспериментах по микроинъекциям гибридных клеток в полость бластоцисты была получена серия химерных животных, что показало сохранение высокой плюрипотентности в гибридных клетках. Однако самым впечатляющим результатом этих опытов было обнаружение функциональной "соматической" Х-хромосомы (происходящей от дифференцированной клетки) в разных тканях и органах химер. Показана возможность репрограммирования индивидуальной хромосомы, происходящей от дифференцированной клетки, при введении ее в геном плюрипотентных эмбриональных клеток. Иными словами, получены сходные данные по обратимости дифференцировки на уровне индивидуальных хромосом генома дифференцированной клетки, подобно реактивации целого генома в опытах Ian Wilmut. Другим следствием этого исследования является возможность переноса индивидуальных хромосом от одного животного в геном другого этого же или близкого вида. Таким образом, открывается перспектива использовать плюрипотентные гибридные клетки в качестве носителей-векторов для переноса индивидуальных хромосом между животными и создавать уникальные генотипы, не существующие в природе, поскольку их невозможно получить обычным половым путем.
|