Федеральное агентство по образованию
Пензенский государственный педагогический университет
им. В.Г. Белинского
|
Факультет
Естественно-географический
|
Кафедра
Биохимии
|
Дипломная работа
Разработка условий оптимизации процесса амплификации ДНК
|
Студент _____________________________________________ Венедиктов А.А.
подпись
|
Руководитель _________________________________________ Белецкий И.П.
подпись
К защите допустить. Протокол № от «____» ___________200_г.
|
Зав. кафедрой _________________________________________ Генгин М.Т.
подпись
|
Пенза, 2008 г.
|
Содержание
Введение
Глава 1.
Обзор литературы
1.1 RealTimeПЦР
1.1.1 Виды RealTimeПЦР
1.1.2 Кинетические параметры
1.2 Биологические микрочипы
1.2.1. ДНК-микрочипы
1.2.2. Гибридизация-основа технологии
1.2.3. Применение ДНК-чипов
Глава 2.
Материал и методы
2.1 Изготовление микропланшетов
2.2 Определение спектра поглощения и эмиссии SYBRgreen
2.3 Протоколы ПЦР
2.3.1 Постановка реакции с использованием фирменного набора(системаSYBgreen)
2.3.2 Постановка реакции без использования фирменного набора(системаSYBgreen)
2.3.3 Протокол полимеразной цепной реакции с использованием TaqMan
2.4 Электрофоретическое разделение нуклеиновых кислот в образцах ПЦР-смеси после амплификации
2.5 Система детекции результатов анализа
Глава 3.
Результаты и обсуждение
3.1 Определение зависимости сигнала флуоресценции от концентрации ДНК
3.2 Проведение ПЦР в разных объемах смеси в пробирках
3.3 Проведение ПЦР в пластиковых микропланшетах
3.4 Постановка ПЦР в микрообъемах с отрицательными контролями
3.5 Электрофоретическое разделение нуклеиновых кислот в образцах ПЦР-смеси после амплификации
Выводы
Заключение
Список литературы
Введение.
С возникновением принципиально новых направлений в науке, в клинической диагностике успешно начали применяться методы ДНК-технологий среди которых, безусловно, первое место занимают полимеразная цепная реакция (ПЦР). Не менее мощным и уникальным по своим характеристикам методом является анализ с использованием биологических микрочипов. Этот метод появился сравнительно недавно, но уже интенсивно применяется в практике.
В основе всех ДНК-технологий, лежат процессы амплификации и регистрации молекул ДНК.
Оптимизация условий данных процессов позволит увеличить производительность, снизить стоимость анализа и повысить его качество. Одним из возможных путей решения этого вопроса является объединение в единую технологическую схему современных подходов в детекции ДНК.
В настоящее время возможные применения подобных схем находятся на стадии разработки в ряде зарубежных лабораторий. Наиболее вероятным направлением ближайшего их применения является молекулярная диагностика наследственных заболеваний и другие практические приложения, основанные на выявлении однонуклеотидных полиморфизмов последовательностей ДНК человека.
Цель работы – это попытка создать модель диагностической системы за счет объединения преимуществ двух методов – количественного вида ПЦР – RealTime и биочипового анализа, в связи с чем были поставлены следующие задачи : изготовление микропланшетов с лунками, общий объем которых не должен превышать 2мкл (в настоящее время используются микроплашки с лунками, рассчитанные на 200 мкл), перевод реакции в формат микрочипа и отработка условий ПЦР в микрообъеме (обычно ПЦР ставят в объеме 20-30 мкл,в формате биочипа общий объем реакционной смеси не должен превышать 1-2 мкл).
Работа выполнялась на базе лаборатории клеточной инженерии Института Теоретической и Экспериментальной Биофизики РАН, г. Пущино.
Глава 1. ОБЗОР ЛИТЕРАТУРЫ
1.1.
RealTime
ПЦР
Принцип метода полимеразной цепной реакции был разработан Кэри Мюллисом в 1983 г. Первые сообщения по практическому применению метода появились в 1985 г.[18] С тех пор количество публикаций, где ПЦР используется в качестве одного из методов исследования, занимает одно из первых мест наряду с работами, в которых применяются автоматическое определение нуклеотидной последовательности (сиквенирование ДНК) и гибридизационные технологии.[7]Существует большое количество способов получить данные о концентрации нуклеиновых кислот в пробе методом ПЦР, но все они требуют дополнительных трудоемких этапов работы, связанных с предварительной раститровкой выделенной из анализируемой пробы ДНК, или полученных в ходе ПЦР ампликонов, что приводит к увеличению времени, необходимого для постановки анализа и сложности интерпретации полученных результатов [12,15]. Также, наличие дополнительных этапов работы увеличивает вероятность ошибки и получения недостоверного результата.
На сегодня существует метод, лишенный вышеперечисленных недостатков - это метод ПЦР в реальном времени (Real-Time PCR) [14]. Сущность метода заключается в исследовании накопления продуктов амплификации с помощью специального прибора без последующего электрофореза. Так как кинетика накопления продуктов амплификации связана с исходным количеством матрицы, это дает возможность точно оценить её количество [13].
Отличительными чертами данного метода, в отличие от классической ПЦР, является возможность количественного определения ДНК/РНК инфекционных агентов в исследуемом материале, отсутствие стадии электрофореза, менее строгие требования к организации ПЦР-лаборатории и автоматическая регистрация и интерпретация полученных результатов.
Отсутствие стадии электрофореза позволяет минимизировать риск контаминации продуктами ПЦР и таким образом резко уменьшить число ложноположительных результатов. Поскольку регистрация результатов проводится непосредственно в процессе ПЦР, весь анализ можно проводить в одной-двух комнатах лаборатории и нет необходимости в отдельном помещении для детекции продуктов реакции.
Использование математических методов анализа позволяет проводить автоматическую интерпретацию полученных результатов и снимает проблему субъективной оценки электрофореграмм.
1.1.1. Виды
RealTime
ПЦР
Для выявления продуктов амплификации в режиме реального времени используют следующие наиболее распространенные подходы.
Выщепление 5' концевой метки (TaqMan Assay).
Данная методика основана на использовании 5'-экзонуклеазной активности полимеразы. В реакционную смесь добавляют ДНК-зонды, в состав которых входит флуоресцентная метка в 5'-положении и гаситель флуоресценции в 3'-положении, а также фосфатная группа в 3'-положении. Эти зонды имеют места посадки внутри амплифицируемой области. Гаситель поглощает испускаемое флуоресцентной меткой излучение, а фосфатная группа в 3'-положении блокирует полимеразу (Рис. 1) В ходе ПЦР во время стадии отжига праймеров происходит присоединение ДНК-зонда к комплементарной цепи ДНК, причем чем больше продуктов амплификации образуется в ходе ПЦР, тем больше молекул зондов свяжется с соответствующими ампликонами. Во время стадии элонгации полимераза синтезирует комплементарную цепь ДНК и при достижении зонда начинает его расщеплять благодаря наличию 5'-экзонуклеазной активности. Таким образом происходит разъединение флуоресцентной метки и гасителя, что приводит к увеличению детектируемого свечения [13]. Очевидно, что чем больше ампликонов было наработано в ходе ПЦР на данный момент времени, тем интенсивнее будет свечение[19].
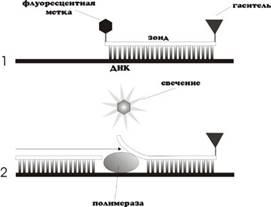
Рис. 1
. Схема ПЦР в реальном времени с использованием зонда TaqMan.[7]
С помощью серийных разведений образцов ДНК определили чувствительность системы ПЦР в реальном времени, которая в обсуждаемой модели позволяла обнаруживать 0,28 нг (приблизительно 20 копий ) ДНК в пробе[8]
Использование зондов с комплементарными концевыми последовательностями (molecular beacons).
Данная методика отличается от описанной выше тем, что концевые последовательности зонда представляют собой взаимно комплементарные области, поэтому при температуре отжига праймеров они схлопываются и образуют шпильки [16](Рис. 2). Внутренняя область зондов содержит нуклеотидную последовательность, комплементарную амплифицируемой области. При отжиге праймеров зонды, не присоединившиеся к ДНК матрице, остаются в "схлопнутом" состоянии, так что происходит тушение флуоресценции.
Те же зонды, которые отжигаются на матрицу, разворачиваются, и флуоресцентная метка и гаситель расходятся в разные стороны. Таким образом, увеличивается интенсивность свечения. Системы ПЦР в реальном времени, основанные на Использовании зондов с комплементарными концевыми последовательностями, обладают гораздо большей специфичностью чем система TaqMan, и позволяет обнаруживать одиночные замены нуклеотидов в ДНК.[8]
Применение 2-х зондов с резонансным переносом энергии (Fluorescentresonanceenergytransfer - FRET).
Данный способ детекции накопления продуктов амплификации отличается повышенной специфичностью, так как увеличение флуоресценции происходит при комплементарном связывании с ампликонами сразу 2-х ДНК зондов [11] (Рис 3). Принцип метода заключается в переносе энергии от одного флуорофора, находящегося на 3` конце первого зонда, ко второму флуорофору, находящемуся на 5` конце второго зонда, причем расстояние между флуорофорами составляет 1-3 нуклеотида.
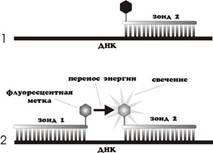
Рис. 3.
Схема ПЦР в реальном времени с использованием зондов FRET[7].
При одновременном связывании обоих зондов с ДНК матрицей испускаемое первым флуорофором излучение передается на второй флуорофор, а его излучение детектируется прибором. Таким образом, возрастает специфичность анализа.
Использование интеркалирующих агентов.
Этот способ детекции основан на том факте, что флуоресценция бромистого этидия и SYBR Green I значительно возрастает при их внедрении в двухцепочечные молекулы ДНК [14] (Рис. 4). Таким образом, можно наблюдать за накоплением продуктов амплификации. Очень важно отметить то, что увеличение флуоресценции может быть связано как с накоплением специфического продукта, так и неспецифического (праймеры-димеры, шмер). Для получения корректных результатов необходимо дополнительное изучение полученных ампликонов с помощью построения так называемых "кривых плавления".
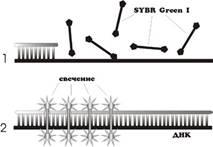
Рис. 4
. Схема ПЦР в реальном времени с использованием интеркалирующих красителей [7].
Подводя итоги, стоит отметить следующее: использование ДНК-зондов в том или ином варианте является наиболее предпочтительным в свете повышения специфичности анализа. Однако к недостаткам зондов относится высокая стоимость, что делает работу по подбору зондов, праймеров и условий амплификации дорогостоящей. Вместе с тем, использование интеркалирующих агентов является очень простым и дешевым. Отпадает необходимость подбора специальных праймеров, зондов, так как можно пользоваться уже используемыми праймерами, эффективность работы которых уже проверена. Эти обстоятельства делают применение интеркалирующих агеннтов весьма привлекательным.
1.1.2 Кинетические параметры
Кинетическая кривая.
Флуоресцентные красители обеспечивают флуоресценцию, прямо пропорциональную количеству ПЦР-продукта - репортерную флуоресценцию. Механизмы генерации репортерной флуоресценции различаются в зависимости от типа real-time PCR.
Кинетическая кривая PCR в координатах "Уровень репортерной флуоресценции — цикл амплификации" имеет сигмоидную форму (Рис. 5).
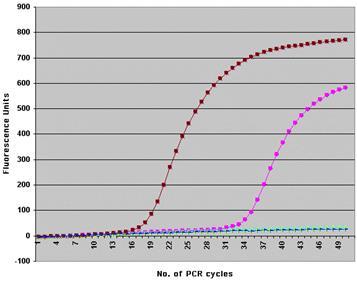
Рис 5
. Пример кинетических кривых RealTimePCR(по материалам www.molbiol.ru).
В ней можно выделить три стадии:
1. Стадию инициации
(когда PCR-продукты еще не детектируется флуоресцентной меткой).
2. Экспоненциальную стадию
(в которой наблюдается экспоненциальная зависимость количества флуоресценции от цикла PCR).
3. Плато
(стадию насыщения).
Экспоненциальная стадия PCR описывается уравнением:
Pn
= P0
* E
n
(1) (www.molbiol.ru),
где Pn
- количество молекул продукта/репортерной флуоресценции к циклу n
, P0
- исходное количество молекул, содержащих амплифицируемый фрагмент (template), E
- эффективность амплификации. В идеальных условиях E
= 2, т.е. на каждом цикле цепной реакции происходит удвоение количества продукта.
Прологарифмируем обе части уравнения 1 и преобразуем его к виду:
n
= - (1/log E
) * log P0
+ log Pn
/log E
(2) (www.molbiol.ru)
Назовем пороговым циклом (threshold cycle, C(T)
) такой цикл n
, на котором достигается некий заданный уровень репортерной флуоресценции - пороговая флуоресценция PC(T)
=const
. Дляn=C(T)
уравнение 2 принимаетвид:
C(T)
= - (1/log E
) * log P0
+ log PC(T)
/log E
(3) (www.molbiol.ru),
т.е. значение С(T)
прямо пропорционально логарифму количества субстрата(по материалам www.molbiol.ru).. Таким образом, real-time PCR позволяет сравнивать количества субстрата при условии, что эффективность реакции и заданный уровень пороговой флуоресценции одинаковы для каждой из сравниваемых реакций [19].
Базовый уровень флуоресценции.
Это тот уровень флуоресценции, который наблюдается в реакционной смеси до появления репортерного сигнала. Как правило, управляющие программы real-time PCR позволяют задать базовый уровень несколькими способами. Необходимо выбрать такой способ, при котором:
1. Обеспечивается минимальный уровень флуоресценции до появления репортерного сигнала.
2. Начало экспоненциальной фазы детектируется с наибольшей точностью.
В общем случае, базовую линию следует выбирать как среднее значение флуоресценции в достаточно широком диапазоне циклов до появления репортерной флуоресценции (т.е. до начала детекции экспоненциальной фазы). При выборе диапазона желательно также избегать и циклы вне экспоненциальной фазы, в которых наблюдаются скачки флуоресценции.
Для SYBR Green, как правило, хорошо работает расчет базового уровня по среднему значению в диапазоне циклов с 3 по 7 (первые 2 цикла не влючают в диапазон, поскольку на них может наблюдаться более сильная флуоресценция из-за недостаточно стабилизировавшейся реакции).
Более осторожно нужно выбирать базовую линию при TaqMan
амплификации, поскольку к началу PCR в смеси может наблюдаться довольно высокий уровень флуоресценции от "сломанных проб" (в которых произошла диссоциация маркера от тушителя). Если "сломанные пробы" не превалируют, это не представляет большой опасности, и базовая флуоресценция постепенно сходит на нет (обычно к 3-7 циклу). Как уже было сказано выше, в таком случае начальный цикл диапазона нужно выбирать такой, в котором флуоресценция от "сломанных проб" уже достигла минимума
.
В некоторых случаях флуоресценция от "сломанных" проб наблюдается и на более поздних стадиях PCR, к 20-25 циклу. Такой эффект нежелателен, поскольку он создает трудности при выборе базовой линии, что может привести к "маскировке" начала экспоненциальной фазы. В таком случае рекомендуется собирать реакцию заранее
и инкубировать ее в холодильнике 1 час или более до добавления ДНК (если реакции инкубируются в темноте и при температуре около +4o
C, максимальное время инкубации практически неограничено) [20].
Свидетельствами того, что базовый уровень выбран некорректно, являются:
1. Значительное "провисание" кривой флуоресценции ниже нулевого уровня.
2. Положительное значение флуоресценции в точке перегиба (т.е. начала экспоненциальной фазы).
"Провисание" часто является следствием включения в расчетный диапазон циклов, при которых уже наблюдается начало экспоненциальной фазы или, наоборот, начальная флуоресценция. Однако "провисание" может быть вызвано также и выбором слишком узкого диапазона циклов, так что разброс базовой флуоресценции в нем оказывается существенно меньше, чем в среднем по всем циклам.
Положительное значение флуоресценции в точке перегиба также обычно "исправляется" расширением диапазона циклов. Такой эффект также может наблюдаться, если расчет ведется по минимальным, а не по средним значениям диапазона.
Напомним, что вытекающая из уравнения (1) зависимость, отражаемая уравнением (3)позволяет сравнивать результаты различных экспериментов лишь при условии, что PC(T)
= const
. Таким образом, уровень пороговой флуоресценции PC(T)
должен быть одинаков для всех сравниваемых образцов.
Пороговый уровень флуоресценции.
Репортерная флуоресценция наиболее точно отвечает зависимости 3 в самом начале детекции экспоненциальной фазы. На более поздних этапах вклад стохастических эффектов в кинетику реакции становится все более значительным. Поэтому пороговый уровень флуоресценции выбирают в самом начале экспоненциальной фазы, когда стохастические процессы не столь велик.
Существует три основных способа выбора уровня пороговой флуоресценции:
а) на глаз;
б) по превышению над стандартным отклонением кинетической кривой;
в) по второй производной кинетической кривой.
При выборе "на глаз
" за пороговый уровень выбирают минимальный уровень флуоресценции, который во всех проведенных с данной парой праймеров реакциях соответствует началу экспоненциальной фазы. Хотя пороговый уровень должен быть как можно ниже, при его достижении реакция должна уже "устаканиться" - на первом цикле экспоненциальной фазы погрешность определения репортерной флуоресценции обычно достаточно высока (в том числе и из-за того, что выбор baseline также имеет определенную погрешность). Обычно оптимальными являются пороговые уровни порядка 0.05-0.1 единиц флуоресценции, в зависимости от способа задания базовой линии и от того, как ведет себя реакция. Выбор "на глаз" является наиболее корректным с точки зрения уравнения 3, при условии, что базовая линия выбрана одинаково и измерения проводятся одновременно. Таким образом, с нашей точки зрения, им имеет смысл пользоваться, когда необходимо сравнить данные в одной плашке (или, в крайнем случае, нескольких экспериментов, выполненных с использованием одинаковых реактивов, субстратов, на одной и той же машине с небольшим промежутком времени) - например, при отладке реакции. При сравнении более разнородных данных определение "на глаз" будет некорректным. Принцип двух других методов состоит в том, чтобы стандартизировать выбор порога не по абсолютному значению, а по поведению кривой. Выбор порога по превышению над стандартным отклонением кинетической кривой
- это установка порога таким образом, чтобы превышение порогового уровня над базовым уровнем (threshold над baseline) было достаточно малым, но уже статистически достоверным. Обычно выбирают значения в диапазоне 8-10 стандартных отклонений. Недостатком такого метода является то, что поскольку стандартное отклонение - это усредненная величина по всей кинетической кривой, в каких-то конкретных случаях порог может оказаться слишком низким (когда реакция еще не "устаканилась") или, наоборот, слишком высоким (когда уже наблюдаются какие-то отклонения). Этих недостатков лишен следующий метод выбора порога - по второй производной кинетической кривой
. В таком случае, из всех скачков уровня флуоресценции за пороговый уровень выбирается наиболее резкий скачок - абсолютный максимум второй производной. Этот способ наименее чувствителен к способу выбора базовой линии и различиям условий реакции (по материалам www.molbiol.ru).
Анализ кривой плавления
. Оценку качества реакции проводят по специфичности амплификации, определяемой из кривой плавления (для SYBR Green).Кривая должна быть построена в диапазоне температур от температуры отжига (при более низких температурах это делать не имеет смысла) до температуры полной денатурации - примерно 95O
C. Для этого после окончания ПЦР реакционную смесь нагревают и непрерывно измеряют флуоресценцию. По достижении температуры плавления продукта амплификации флуоресценция резко снижается.

Рис. 6
. Пример кривой плавления [7].
Каждое резкое уменьшение флуоресценции на графике соответствует числу полосок, получаемых на электрофорезе, то есть числу разных типов ампликонов. Для облегчения работы с полученной информацией проводят дифференциальный анализ кривой плавления. Такой способ визуализации полученных данных гораздо удобнее для понимания и анализа.
Применение кривых плавления не ограничивается только детекцией продуктов амплификации с помощью бромистого этидия и SYBR Green I. При использовании кривых плавления в системах с ДНК-зондами (Taq-man assay, beacons) возможно различать точечные мутации, расположенные внутри областей связывания ДНК-матрицы и зонда. Наличие таких мутаций способно привести к изменению температуры плавления зонда и к изменениям в графике кривой плавления [11]. Использование кривых плавления не требует от оператора амплификатора никаких дополнительных манипуляций с пробирками, а интерпретация полученных данных автоматизирована и формализована.
В идеальном случае, в реакциях с матрицей мы должны наблюдать один пик (соответствующий плавлению специфического продукта), а контроль "праймеры без ДНК" - слабые, затухающие стохастические колебания во всем диапазоне температур.
Обработка данных.
Методы обработки данных real-time PCR основаны на применении уравнения (3).Основной принцип обработки данных Real-time PCR – это -
метод калибровочного графика.
Этот метод предполагает построение калибровочного графика в координатах C(T)
- log P0
с серией разведений ДНК-стандарта, из которого находят концентрацию субстрата (P0
) в экспериментальных образцах.
Точность метода зависит от того, насколько условия PCR (и, прежде всего, эффективность амплификации) серии стандартов близки к условиям PCR экспериментальных образцов. Поэтому при выборе стандартов необходимо руководствоваться следующими правилами:
а) сдержание примесей в стандартном препарате должно быть сходно с таковым в экспериментальном препарате;
б) для амплифицируемого фрагмента в стандартной ДНК должна быть близка к таковой в экспериментальной ДНК;
в) серия разведений стандартной ДНК должна охватывать весь диапазон возможных концентраций субстрата в экспериментальной ДНК.
Если достаточно определить лишь относительную концентрацию субстрата, для построения калибровочного графика удобно использовать серию разведений одного из экспериментальных образцов. В тех случаях, когда образцы могут сильно отличаться по примесям и доле амплифицируемого фрагмента в реакционной смеси, имеет смысл при наладке эксперимента приготовить серию разведений каждого из них и сравнить эффективности.
В тех случаях, когда требуется оценить "абсолютное" количество субстрата, выбор стандартов для градуировочного графика представляет собой более сложную задачу, поскольку с одной стороны, необходимо знать точное число. Для определения количества матрицы в RT-PCR предложены следующие варианты стандартов:
1. очищенный RT-PCR-продукт;
2. рекомбинантная ДНК;
3. рекомбинантная РНК с последующей обратной транскрипцией;
4. синтетический олигонуклеотид, содержащий амплифицируемую последовательность.
При использовании очищенных препаратов в реакцию всегда необходимо добавлять неспецифические ДНК, чтобы приблизить условия реакции к экспериментальным. Для RT-PCR это могут быть, например, обратные транскрипты бактериальной РНК, polyA-РНК, тРНК или рРНК. Однако насколько бы условия реакции со стандартными образцами не были приближены к экспериментальным, "абсолютное" определение по-прежнему остается расчётом относительно стандарта.
1.2. БИОЛОГИЧЕСКИЕ МИКРОЧИПЫ
В последнее время активно развиваются ДНК-технологии, которые позволяют не только определять признак, но и одновременно проводить дифференциальный сиквенс, т.е. определение точечных мутаций или полиморфизма в известных участках генома. Данные технологии имеют значительные преимущества перед традиционными молекулярно-биологическими методами, т.к. они позволяют миниатюризировать исследуемый образец и анализатор, что значительно снижает стоимость анализа и время его проведения, а также одновременно определять различные параметры исследуемого образца, причем без потери чувствительности амплификационных методов [6].Главным преимуществом методов основанных на использовании чипов с олигонуклеотидами всех возможных нуклеотидных последовательностей данной длины, является их универсальность. Наличие на чипе олигонуклеотида любой последовательности делает возможным анализ любой исследуемой последовательности [4]. В основе применения микрочипов лежит принцип быстрого определения взаимодействий тех или иных лигандов со множеством различных зондов одновременно. Собственно биологические микрочипы представляют собой ту или иную твердую подложку, на которой нанесены или определенные фрагменты нуклеиновых кислот, или белки, или углеводы, или какие-либо иные молекулы-зонды, способные быть узнанными или проявлять биологическую активность. Количество различных зондов на подложке может достигать сотен тысяч, причем чипы каждого типа строго идентичны и при существующих технологиях могут быть реплицированы в сотнях тысяч и миллионах копий нанесенных на подложку [1].
1.2.1 ДНК-микрочипы
Существуют белковые, ДНК, углеводные, тканеые чипы. Особого внимания заслуживают ДНК-чипы.Они представляют собой уникальный аналитический инструмент, позволяющий определять наличие в анализируемом образце (как правило, биологического происхождения) заданных последовательностей ДНК (т.н. гибридизационный анализ). Проведение анализа с помощью ДНК–чипов обходится в несколько раз дешевле, чем при использовании альтернативных технологий (электрофорез, ПЦР в реальном времени) и допускает, при наличии детектора несложной конструкции, работу вне лаборатории.
Впервые ДНК–чипы были использованы в исследованиях в конце 80-х годов прошлого века [10]. В основе этого теперь уже широко распространенного метода, позволяющего одновременно анализировать экспрессию множества генов, лежит принцип узнавания мРНК-овых или кДНК-овых мишеней посредством их гибридизации с иммобилизованными на микрочипе одноцепочечными фрагментами ДНК.
ДНК–чип – это твердая подложка, на которой иммобилизованы (как правило, ковалентно) однонитевые фрагменты ДНК разной длины: короткие – 15-25 нуклеотидов, длинные – 25-60 нуклеотидов и кДНК фрагменты – от 100 до 3000 нуклеотидов. В качестве материала подложки используют стекло, кремний, различные полимеры, гидрогели (например, на основе полиакриламида) [17] и даже золото [3].
1.2.2. Гибридизация-основа технологии
Основа всех современных ДНК-технологий – гибридизация. В результате гибридизации молекулы нуклеиновых кислот способны формировать устойчивые двухцепочечные структуры благодаря связям между элементами молекул - нуклеотидами. Нуклеотид аденин (А) комплиментарен тимину (Т), гуанин (Г) - цитозину (Ц). В результате одноцепочечная последовательность нуклеотидов АТГЦ будет образовывать стабильную ассоциацию, двухцепочечную структуру, с одноцепочечной молекулой ДНК состава ТАЦГ.
…..АТГЦ….
| | | |
…..ТАЦГ….
Подобная комплементарность приводит к «слипанию» двух молекул ДНК, одна из которых может быть неподвижно закреплена на подложке, и являться элементом ДНК-чипа. Чем больше содержится в образце молекул комплиментарных элементам чипа, тем больше их будет связываться с чипом, и тем выше будет интенсивность сигнала, воспринимаемого от данного элемента. На рис. 9 показан принцип действия ячейки ДНК или олигонуклеотидного биочипа, основанный на комплементарных взаимодействиях основания аденина (А
) с тимином (Т
) и гуанина (G
) с цитозином (С
) в двух нитях ДНК. Если последовательность оснований в одной нити ДНК (или олигонуклеотида) полностью комплементарна последовательности другой нити, то образуется стабильная совершенная двухнитчатая спираль - дуплекс. Однако присутствие в дуплексе даже одной неправильной пары, например G-G
, предотвращает образование дуплекса. Если иммобилизовать в одном из элементов микрочипа специфическую одноцепочечную ДНК или, положим, 20-мерный олигонуклеотид (пробу), то при добавлении к микрочипу меченных флюоресцентными красителями фрагментов ДНК, например генома человека, будет происходить их высокоспецифичное взаимодействие. Заданный олигонуклеотидный элемент биочипа специфически свяжет только одну комплементарную последовательность из 420
=1.09х1012
всех возможных последовательностей этой длины в ДНК. В результате флюоресцентное свечение наблюдается только на этом комплементарном элементе биочипа. Таким образом, один элемент биочипа производит одну выборку примерно из триллиона возможных вариантов, в отличие от элемента электронного чипа, где происходит двоичная выборка: ДА
или НЕТ
[5].
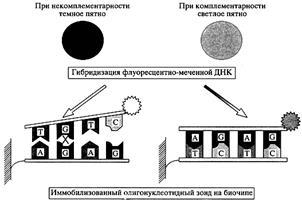
Рис. 9.
Схема образования двойной спирали ДНК на биочипе.Олигонуклеотид фиксирован на одном из элементов биочипа и избирательно связывает из многих флуоресцентно меченых фрагментов ДНК только комплементарный. В результате только этот элемент начинает светиться. Это происходит благодаря высоко-специфичным взаимодействиям комплементарных пар нуклеотидов А
с Т
и G
с С
. Присутствие некомплементарной пары, например G-G
, предотвращает взаимодействие и оставляет элемент микрочипа темным [5].
Используемые для определения параметров гибридизации устройства позволяют регистрировать не только конечный результат, но и кинетику ассоциации и диссоциации комплементарных цепей. Эти технологии, делая возможным многопараметрический анализ образцов, могут предоставлять большое количество информации. Результаты гибридизации зависят от длины ДНК - пробы, химического состава меченой ДНК - мишени, температуры, при которой проводится гибридизация, состава гибридизационной смеси, типа флуоресцентной метки. Здесь необходимо отметить, что в ДНК-чипах в основном используется пассивная гибридизация, т.е. взаимодействие ДНК-мишени с иммобилизованной пробой является вероятностным процессом и зависит от различных условий.
1.2.3. Применение ДНК-чипов
Оценка состояния и идентификация всех генов исследуемого организма является одной из важнейших задач, поставленных перед разработчиками ДНК-чипов. Решение этой задачи может быть реализовано в иммобилизации всех генов организма на биологический чип, что позволит комплексно оценить состояние генов и генома в целом. Биогенетические базы данных, содержащие всю информацию (систематизированную) по генам и геномам различных организмов, представляют исследователям огромные возможности в дизайне ДНК-чипов.
К основным причинам широкого распространения биочиповых исследовианий относят высокую чувствительность, специфичность и воспроизводимость, простоту процедуры выполнения, возможность одновременного анализа множества параметров и относительно невысокую стоимость работ. Эти же причины заставляют рассматривать биочипы как перспективный инструмент в различных областях народного хозяйства [9].
Подводя некоторые итоги нужно отметить, что микрочипы являются эффективным подходом для одновременной идентификации от десятков до тысяч генов и их структурного анализа, для выявления специфичных нуклеотидных последовательностей и нуклеотидных вариаций в их структуре. Однако, когда гены присутствуют в геноме в количестве одной или нескольких копий, с чем постоянно и приходится сталкиваться в клинической практике, требуется их предварительная амплификация. Наиболее эффективным методом амплификации ДНК является полимеразная цепная реакция, в процессе которой происходит экспоненциальное увеличение количества молекул ДНК от нескольких до миллионов и более копий, а основное достоинство такого вида ПЦР как RealTime, позволяют давать даже количественную оценку исследуемой матрицы. Это имеет важное значение для решения задач в области развития фундаментальных и интегральных наук, а так же оптимизации условий методов диагностики.
Итак, два метода, ставшие уже традиционными для некоторых областей науки и прикладных технологий,на ряду со своими недостатками имеют совершенно уникальные достоинства.
RealTime
ПЦР:
· дает возможность оценивать количество исходной матрицы;
· не требует дополнительных трудоемких этапов работы;
· отсутствие стадии электрофореза позволяет минимизировать риск контаминации и таким образом уменьшить число ложноположительных результатов;
· использование математических методов анализа позволяет проводить автоматическую интерпретацию полученных результатов и снимает проблему субъективной оценки электрофореграмм;
· обеспечивает менее строгие требования к организации ПЦР-лаборатории и автоматической регистрации и интерпретации результатов;
· позволяет экономить время.
Биологические микрочипы:
· позволяют миниатюризировать образец и анализатор;
· экономит время и стоимость анализа;
· позволяет одновременно определять несколько параметров исследуемого образца;
· убладает высокой чувствительностью амплификационных методов,специфичностью и воспроизводимостью;
· обеспечивает простоту процедуры выполнения работы.
Возможно , что объединие этих методов за счет перевода полимеразной цепной реакции в формат микрочипов позволит создать диагностическую систему нового поколения, которую будут характеризовать следующие качества : более высокая чувствительность и, главным образом, специфичность определения нуклеиновых кислот, высокая производительность при низкой себестоимости выполнения анализа, общее сокращение количества манипуляций в пределах каждого этапа анализа.
Глава 2.
МАТЕРИАЛЫ И МЕТОДЫ
Поставив перед собой цель объединить преимущества методов, мы столкнулись с рядом препятствий, касающихся инструментального обеспечения исследования. Проведение полимеразной цепной реакции в микрообъеме требует определенного технического оснащения. Современный рынок предлагает микропланшеты, объем лунок которых составляет не менее 200 мкл. Проведение ПЦР в микрообъеме в подобных планшетах противоречит условиям нашего эксперимента. Кроме того, современные амплификаторы не позволяют манипулировать формами термоблоков. В связи с этим перед началом исследований необходимо было создание экспериментальной аппаратной базы для амплификации и регистрации сигналов, изготовление микропланшетов с лунками, общий объем которых не должен превышать 2 мкл.
2.1. Изготовление микропланшетов
Для эксперимента были изготовлены пластины размером 50х50 мм с рабочей зоной 20х20 мм с различным количеством лунок (до 91 шт). При изготовлении планшетов мы руководствовались размером термокамер в экспериментальном образце амплификотора, поэтому такие их длина и ширина являются оптимальными. Схема амплификатора представлена на рис. 10.
Материал, из которого должны быть изготовлены плашки должен обладать следующими свойствами:
- термостойкость (материал должен выдерживать минимум 95 С-самая высокая температура в цикле);
- прочность(не должен подвергаться деформации даже при незначительных механических воздействиях);
- химическая инертность (материал не должен взаимодействовать с компонентами ПЦР смеси);
- дешевизна(материал должен быть не дорогим и доступным).
Экспериментально был подобран материал, который обладает всеми вышеперечисленными качествами – это полимерная пленка AikorDecoDesignBlack(производство Германия). Изготовление плашек делали путем термоформовки на металлической матрице с помощью металлического пуансона-позиционера как показано на рис 11. В результате получали планшеты с лунками объем которых бал равен 1,5 мм3
.
2.2 Определение спектра поглощения и эмисии
SYBRgreen
В RealTimeПЦР используют как описано выше несколько видов зондов. В своих исследованиях мы использовали SYBR Green. При выборе зондов мы руководствовались несколькими параметрами, в том числе стоимость, совместимость с системой детекции и т.д. Технологии Molecular beacons и LightCycler стоят ощутимо дорого, и их осмысленно применять лишь в том случае, когда требуется крайне специфичная детекция лишь одного из образующихся PCR-продуктов (например, при анализе SNP). Методики SYBR Green и TaqMan стоят дешевле, однако каждый из них имеет свои особенности. В таблице 1, приведенной ниже, сравниваются методики SYBR Green и TaqMan. В наших исследованиях мы использовали эти системы. Они активно используются в RealTimeПЦР для определения количественных параметров ПЦР непосредственно в ходе реакции. Однако мы для решения поставленных задач пользовались этими методиками для детекции сигнала по конечной точке.
В своей работе мы использовали флуоресцентный интеркалирующий краситель SYBRgreen (10000 кратный сток производства фирмы «ДНК-синтез») и набор компонентов для проведения ПЦР с использованием TaqMan, синтезированных компанией «Синтол» (наработанный продукт амплификации имеет размер около 100 п.н.).
Краситель SYBR Green - исключительно чувствительный флуоресцентный индикатор двухцепочечной ДНК. Применяется для realtime-PCR, а также для прокраски гелей (как агарозных, так и полиакриламидных). Предел обнаружения двухцепочечной ДНК для SYBR Green составляет 1-2 нанограмма для коротких синтетических олигонуклеотидов и до 20 пикограмм высокомолекулярной природной ДНК. SYBR Green I обладает существенно высокой специфичностью в отношении ДНК-дуплекса и является более безопасным по сравнению с другими красителями – он не оказывает высокого уровня мутагенного действия.
| Методика
|
Достоинства
|
Недостатки
|
| SYBR Green
|
Значительно дешевле собрать новую реакцию, т.к. достаточно лишь заказать новые праймеры. |
Невозможность работать больше, чем с одной PCR-реакцией в одной реакционной смеси.
Высокие требования к специфичности реакции - любая двуцепочечная ДНК, в том числе димеры праймеров, будет генерировать репортерную флуоресценцию.
Меньшая точность при малых количествах субстрата.
Окисление со временем
|
| TaqMan
|
Возможность проведения нескольких количествленных реакций в одной смеси.
Меньшие требования к специфичности реакции: репортерная флуоресценция будет генерироваться только при гибридизации пробы со специфическим PCR-продуктом.
Бóльшая точность при малых количествах субстрата. |
Дороговизна. Новый зонд (около $200) для каждой реакции.
Меньше возможностей для оптимизации (если реакция не пошла с данной комбинацией зонда и праймеров, то, скорее всего, придется заказывать новый зонд и новые праймеры). |
Данные о флуоресценции: максимум флуоресценции в комплексе с ДНК составляет 521 нм, максимум возбуждения - 497 нм (второй максимум около 254 нм). Хорошо возбуждается стандартным 488-нм лазером. Квантовый выход флуоресценции - около 0.8 (что в 5 раз превышает квантовый выход комплекса этидий-ДНК). Обычно SYBR Green хорошо работает примерно до C(T) = 35-37, затем точность начинает падать. Диапазон достоверности результатов легко проверить для каждой конкретной пары праймеров, проведя несколько независимых real-time PCR.
В системе TaqManиспользовали зонд, меченный красителем FAMи имеющий следующую последовательность:
5’
(FAM) – СTTGAAAGATCTGCTAGAGTCAGCTTGTCAGCG – (TAMRA) – 3’
. Длина 33 нуклеотида. Молекулярный вес 10137. Длина волны возбуждения FAM 470 нм, длина волны эмиссии 525 нм.
Перед работой определяли спектр поглощения и испускания света интеркалятора на приборе PerkinElmerMPF-44-BFluorescenceSpectrophotometer. Результаты измерений представлены на рис. 13.
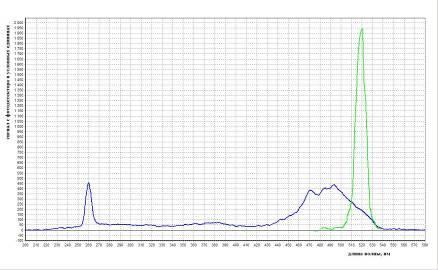
Рис. 13.
График зависимости сигнала фотодетектора от длины волны.
На рисунке синей кривой показан спектр поглощения света красителем. Кривая получена при эмисии образца 520нм. Видно что SYBR поглощает свет длинной 260нм (первый пик). Второй пик приходится на 470 и 495 нм. Зеленая кривая показывает спектр испускания длины волны. Согласно ей, интеркалятор испускает свет с длиной волны 520 нм. Эта кривая получена при возбуждении образца длинной волны 470 нм.
2.3. Протоколы полимеразной цепной реакции:
2.3.1. Постановка реакции с использованием фирменного набора(система SYBgreen)
Для ПЦР в качестве матрицы использовали плазмиду pcDNA3 cavmut – плазмидный вектор, содержащий кДНК мутантного кавеолина (P132L) человека (исходная концентрация 0,7 мг/мл), пару праймеров pc1(исх.конц. 31 мкМ) и pc2 (исх.конц. 12 мкМ), SYBRgreen(10000 кратный раствор) и taq-полимеразу. Фермент поставляется фирмой Sigma в наборе c буфером в лиофильном состоянии в комплекте с растворителем. Кит рассчитан на общий объем смеси 20 мкл. Амлифицировался участок (фрагмент) размером около 650 п.н.
Постановка ПЦР:
· 7 нг матрицы,
· 0,2мкМ pc1,
· 0,2мкМ pc2,
· деионизированная (milliQ) дистиллированная вода до 10 мкл,
· 10мкл растворителя,
· 0,2мкл SYBRgreen(разведенного в 100 раз).
Для эксперимента использовались микропланшеты собственного изготовления. В качестве контроля служила лунка с ПЦР-ной смесью без матрицы. Для отработки условий ПЦР в микрообъеме проводили 30 циклов. Регистрацию флуоресценции проводили по конечной точке. Температурный режим (1 цикл):
| 94о
С 120 сек |
1 цикл |
| 94о
20 сек |
30 циклов
|
| 60о
20 сек |
| 72 о
С 40 сек |
и по окончании нужного количества циклов
72 о
С 120 сек
|
2.3.2. Постановка реакции без использования фирменного набора(система SYBRgreen)
В качестве матрицы использовали плазмиду pcDNA3 cavmut (исходная концентрация 0,7 мг/мл), пару праймеров pc1 (исх.конц. 31мкМ) и pc2 (исх.конц. 12 мкМ), SYBRgreen и комплекс фермента (taq-полимераза) с антителами – так называемый Hotstart, 10-кратный сток буфера, растворы dNTP и MgCl2
. Связывание полимеразы с антителами нужен для того, чтобы фермент начинал свою работу только при высоких температурах (первая температура каждого цикла реакции – 94о
С), а при комнатной оставался неактивным. Общий объем смеси 20 мкл. Амлификации поддавался участок размером около 650 п.н.
Постановка ПЦР:
· 7 нг матрицы,
· 0,2 мкМ pc1,
· 0,2 мкМ pc2,
· деионизированная (milliQ) дистиллированная вода до 14 мкл,
· 2 мкл SYBRgreen,
· 2 мкл буфера (сток х 10),
· 2 мкл 2мМ MgCl2
,
· 0,2 мкл taq-полимеразы (5Е/мл),
· 0,4 мкл 0,2мМ dNTP (нуклеотидтрифосфоты).
Для эксперимента использовались микропланшеты собственного изготовления. В качестве контроля служила лунка с ПЦР-ной смесью без матрицы. Для отработки условий ПЦР в микрообъеме проводили 30 циклов. Регистрацию флуоресценции проводили по конечной точке.
Температурный режим (1 цикл):
| 94о
С 120 сек |
1 цикл |
| 94о
20 сек |
30 циклов
|
| 60о
20 сек |
| 72 о
С 40 сек |
и по окончании нужного количества циклов
72 о
С 120 сек
|
2.3.3. Протокол полимеразной цепной реакции с использованием TaqMan.
На одну пробу объемом 20 мкл смешивают:
· 8,75 мкл смеси А,
· 8,75 мкл смеси Б,
· 0,2 мкл Taq-
полимеразы,
· 2 мкл ДНК.
| 95о
С 5 мин |
1 цикл |
| 95о
С20 сек |
40 циклов
|
| 62о
С20 сек |
| 72 о
С 40 сек |
и по окончании нужного количества циклов
72 о
С 40 сек
|
2.4 Электрофоретическое разделение нуклеиновых кислот в 8% ПААГ
Для приготовления геля было использовано.
| gels |
1 |
2 |
| H2
O (мл) |
3,7 |
7,4 |
| 30%АА/bAA (29:1) (мл) |
1,87 |
3,74 |
| 5х ТВЕ (мл) |
1,4 |
2,8 |
| TEMED (мкл) |
20 |
40 |
| 10% PSA (мкл) |
27 |
54 |
Собирали кассету с вертикальными пластинками. В гель вносили полимеризаторы, перемешивали и заливали 4.2 мл между пластин. После полной полимеризации геля между стекол вставляли гребенку. После полимеризации геля пластины извлекали из кассеты и помещали в камеру для электрофореза. Заливали электродный буфер следующего состава:
0,5 М ЭДТА, рН=8.0,
0,4 BorieAcid,
0,4 Tris.
Извлекали гребенку из геля, не деформируя карманы. Пипеткой или шприцем промывали карманы от неполимеризовавшейся жидкости электродным буфером. Вносили ДНК пробы в карманы пипеткой, V=5 мкл. Подключали камеру к источнику тока. Электрофорез проводили от минуса к плюсу при постоянной силе тока I=20мA на одну пластину, пока лидирующий краситель не выходил из геля (бромфеноловый синий), около 40 минут.
2.5. Система детекции результатов анализа
После амплификации регистрацию флуоресцентного сигнала проводили на экспериментальной установке, собранной на базе бинокулярного микроскопа МБС-11 (Россия ) c телевизионным адаптором TV-A и камерой фирмы «Wateс» (Japan) WAT-120 N. Сигнал с камеры подавался на плату видеозахвата «PixelViewCX 881 P», находящийся на шине IDE персонального компьютера типа IBMPC. С помощью установки регистрировали сигнал (возбуждение – 470 нм, эмиссия – 520 нм), обрабатывали с помощью программы «Qwantа» (оригинальная разработка лаборатории).
Он поделен на 4 части. Первая –Capturewindow – это онлайн окно, в поле которого проэцируется изображение лунки с образцом. Левое темное поле – это поле захвата изображения. Здесь с помощью настроек можно указывать расположение области измерения, границы которого обозначены зеленой окружностью, устанавливать размер и сохранять изображение. Нижние поля – «расчетные данные эксперимента» и «измерения в текущем кластере» дают возможность снимать показания измерений и манипулировать их расчетами. Программа так же позволяет снимать запоминать и обрабатывать показания флуоресценции не только с одного, как показано на рисунке, но и с нескольких рядов лунок с образцами.
Оборудование состоит из двух корпусов. Корпус номер 2 имеет предметный столик, куда кладется анализируемый чип. На корпусе 1 закреплены светодиодная матрица и цифровая ТV-камера. Обьектив камеры имеет съемные оптические фильтры. При анализе образцов с SYBRgreenиспользовали зеленый светофильтр, соответствующий длинне волны испускаемого им света. Светодиодная матрица состоит из нескольких источников света, длина волны которого соответствует спектру возбуждения красителя. На рисунке это изображено синими стрелками. Зелеными показана эмиссия.
Глава 3. РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
3.1. Определение зависимости сигнала флуоресценции от концентрации ДНК
Готовили разведение матрицы с концентрациями 7, 3.5, 1.75, 0.875 и 0.219 нг. К растворам ДНК добавляли одинаковое количество красителя SYBRgreen согласно протоколу описанному в пункте 2.3.1. Образцы объемом 1 мкл помещали в лунки планшетов. Измерение и регистрацию флуоресцентного сигнала проводили с помощью описанной системы детекции и программы «Qwantа». Результаты измерений представлены в таблице 2 и рис. 16.
Таблица 2
Концентрация ДНК (нг)
|
Флуоресцентный сигнал (у.е.)
|
| Измерение
|
Отклонение
|
| 7 |
180 |
+/-15 |
| 3,5 |
170 |
+/-13 |
| 1,75 |
142 |
+/-11 |
| 0,875 |
63 |
+/-7 |
| 0,437 |
41 |
+/-4 |
| 0,219 |
21 |
+/-2 |
| 0 |
17 |
+/-2 |
На графике (рис. 16) видно, что зависимость сигнала флуоресценции от концентрации ДНК имеет некую кривую. Разброс значений по мере роста концентрации ДНК увеличивается.

Рис 16
. График зависимости флуоресцентного сигнала от концентрации ДНК.
3.2. Проведение ПЦР в разных объемах смеси в пробирках
Перед проведением реакции в микроплашках ПЦР провели в пробирках Eppendorfс общим объемом смеси 5, 4, 3, 2 и 1 мкл (исходная концентрация ДНК – 7 нг) в среде масла. Подобная модель миниатюризации реакции может дать ответ на вопрос в каком минимальном объеме возможно прохождение амплификации и насколько эффективно. Данные о приросте сигнала флуоресценции будут своеобразным контролем для реакции смеси соответствующего объема в изготовленных микропланшетах.
ПЦР-смесь заливали минеральным маслом объемом 10 мкл. В результате получали капельки образца в среде масла (рис. 17 А).
   
Рис. 17.
Пробирки с разным объемом ПЦР смеси в среде минерального масла. А – схематическое изображение. Б – реальное изображение капли в объективе камеры.
В качестве контроля служила смесь без матрицы в соответствующих объемах. Амплификацию проводили в амплификаторе «Терцик» производства «ДНК-технология». Реакцию ставили с использованием зондов SYBRgreenи TaqManпо протоколам описанным в пункте 2.3.2 и 2.3.3 соответственно. После амплификации из каждой пробы отбирали по 1 мкл на измерения в стрипах. Тем самым измеряли флуоресценцию в одинаковых объемах. По результатам этих измерений судили об эффективности амплификации в каждой капле. Результаты детектировали на описанном выше оборудовании в пробирках и в перенесенных в микропланшетах (объем анализируемой смеси одинаков во всех пяти случаях – 1 мкл) (рис. 18.).
 А А  Б Б
Рис 18.
Лунка микропланшета с образцом объемом 1 мкл. А – в разрезе, Б – вид в объектив камеры (фотография).
Результаты измерений представлены в таблицах 2 и 3.
Таблица 2.
Объем смеси
(мкл)
|
Образцы с матрицей
(у.е. флуоресценции)
|
Образцы без матрицы
(у.е. флуоресценции)
|
Приращение сигнала
(у.е. флуоресценции)
|
Образцы с матрицей
(у.е. флуоресценции) |
Образцы без матрицы
(у.е. флуоресценции)
|
Приращение сигнала
(у.е. флуоресценции)
|
| в микропробирках |
в объеме 1 мкл в пластиковых микрострипах |
| 1
|
130,43 |
119,12 |
11,31 |
59,44 |
31,38 |
28,06 |
| 2
|
129,82 |
111,55 |
18,25 |
50,89 |
44,01 |
6,88 |
| 3
|
176,10 |
150,34 |
25,76 |
63,01 |
59,86 |
3,14 |
| 4
|
162,77 |
155,13 |
7,64 |
65,70 |
55,88 |
9,82 |
| 5
|
161,55 |
149,38 |
12,17 |
70,00 |
52,70 |
17,30 |
Таблица 3.
Объем смеси
(мкл)
|
Образцы с матрицей
(у.е. флуоресценци)
|
Образцы без матрицы
(у.е. флуоресценции)
|
Приращение сигнала
(у.е. флуоресценции)
|
Образцы с матрицей
(у.е. флуоресценции)
|
Образцы без матрицы
(у.ефлуоресценции)
|
Приращение сигнала
(
у.е. флуоресценции)
|
| в микропробирках |
в объеме 1мкл в пластиковых микрострипах |
| 1
|
218,43 |
199,52 |
18,91 |
98,48 |
78,61 |
19,87 |
| 2
|
226,78 |
202,54 |
24,24 |
90,61 |
81,81 |
8,80 |
| 3
|
187,78 |
156,85 |
30,93 |
92,38 |
76,21 |
16,17 |
| 4
|
199,48 |
166,48 |
33,01 |
112,30 |
97,60 |
14,70 |
| 5
|
215,14 |
186,65 |
28,49 |
– |
65,43 |
– |
По результатам видно, что амплификация прошла во всех пробах, так как во всех случаях число, означающее приращение флуоресцентного сигнала, является положительным. Если обратить внимание на приращение сигнала в образцах измеряемых в микрострипах и сравнить с друг с другом, можно сделать вывод, что амплификация в одном мкл прошла не менее эффективно, чем в остальных объемах. Следующий этап работы – это проведение реакции в пластиковых микропланшетах в объеме смеси 1 мкл.
3.3. Проведение ПЦР реакции в пластиковых микропланшетах
ПЦР проводили по вышеописанному протоколу (используя кит) на планшете с 25 лунками. ПЦР смесь добавляли в лунки объемом 1 мкл. Использовалось 2 вида контрольных проб. Первый контроль – это пцр смесь без матрицы. Второй контроль – это водный раствор SYBRgreen той же концентрации что и в пцр смеси. Схема эксперимента представлена на рис. 19. Измерение и регистрацию флуоресцентного сигнала проводили с помощью описанной системы детекции и программы «Qwantа».

Рис. 19.
Схема нанесения проб в лунки планшета.

Рис. 20.
Результат амплификации в пластиковых микропланшетах.
По результатам эксперимента (рис. 20) видно что в 1 и 3 ряду амплификация прошла, так как интенсивность флуоресценции заметно возросла. Темное пятнышко по центру лунок – это некоторый объем воздуха который образуется при покрытии плашки пленкой. Это указывает на то, что практически весь объем лунки заполнен образцом.
В этих же рядах в лунках под номером 5 сигнал значения не изменил. Это доказывает то, что матрица здесь не присутствовала. Контроль номер 2 также сработал, так как флуоресценция в 4 ряду свое значение не изменила. Второй и пятый ряды практически не флуоресцируют. Это подтверждает наличие в их лунках воды. Значения сигнала флуоресценции каждой лунки представлены в таблице 4. Графическое изображение полученной картины представлено на рис. 21.
| 1 столбец |
2 столбец |
3 столбец |
4 столбец |
5 столбец |
| 1 ряд |
49,82 |
73,54 |
84,25 |
83,68 |
38,91 |
| 2 ряд |
17,61 |
35,76 |
34,41 |
31,66 |
24,56 |
| 3 ряд |
72,26 |
92,30 |
117,95 |
113,88 |
41,02 |
| 4 ряд |
26,88 |
34,04 |
40,36 |
43,36 |
40,88 |
| 5 ряд |
10,34 |
12,81 |
17,81 |
20,89 |
19,08 |
Таблица 4
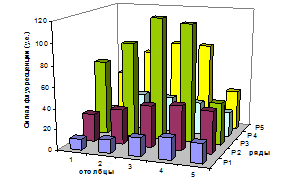
Рис. 21.
Диаграмма, отображающая уровень сигнала в лунках пластиковой микроплашки.
3.4. Постановка ПЦР в микрообъеме с отрицательными контролями
ПЦР ставили по протоколам с использованием SYBRgreenи TaqMan. Общий объем ПЦР смеси составлял 1 мкл. Количество внесенной матрицы одинаково – 7 нг. Контролем служили пробы без матрицы. Измерение и регистрацию флуоресцентного сигнала проводили с помощью описанной системы детекции и программы «Qwantа». Результаты измерений представлены в таблице 5.
По данным таблицы видно, что приращение сигнала флуоресценции сопоставимо с измерениями, полученными при проведении ПЦР в капле смеси объемом 1 мкл в среде масла в обоих случаях. А значит амплификация проходит с не меньшей эффективностью чем на моделях с каплями. Сильный разброс значений в случае SYBRgreen лишний раз доказывает, что работа с микрообъемами – это очень сложный процесс, где даже самые минимальные погрешности в работе (погрешности пипеток, чистота преперата и т.д.) становятся критичными и влияют на результат.
Таблица 5.
| SYBRgreen
|
| С матрицей
(флуоресценция, у.е.) |
Среднее значение (среднее отклонение) |
Без матрицы
(флуоресценция, у.е) |
Среднее значение (среднее отклонение) |
Приращение
(флуоресценция, у.е) |
Среднее значение(среднее отклонение) |
| 41 |
40,8(3)
|
34 |
35,4(4)
|
7 |
5,4(3,5)
|
| 40 |
31 |
9 |
| 38 |
36 |
2 |
| 44 |
39 |
5 |
| 41 |
37 |
4 |
| TaqMan
|
| С матрицей
(флуоресценция, у.е) |
Среднее значение (среднее отклонение) |
Без матрицы
(флуоресценция,у.е) |
Среднее значение (среднее отклонение) |
Приращение
(флуоресценция,у.е) |
Среднее значение(среднее отклонение) |
| 82 |
89,2(8,5)
|
57 |
58,2(1,5)
|
25 |
31(19)
|
| 99 |
55 |
44 |
| 91 |
60 |
31 |
| 89 |
60 |
29 |
| 85 |
59 |
26 |
3.5. Электрофоретическое разделение нуклеиновых кислот в образцах ПЦР-смеси после амплификации
ПЦР ставили по протоколам с использованием SYBRgreenи TaqMan. Амплификация проходила в микропланшетах в объеме смеси – 1 мкл (на геле эти образцы обозначены прописной буквой «ч») и как контроль в амплификаторе «ТерЦик» под минеральным маслом в объеме 20 мкл(на геле – это образцы с индексом «а»). Заглавные буквы Tи Sозначают ПЦР с системой TaqManи SYBRgreen соответственно. Индексы 1, 2 и 3 в системе TaqMan означают использование разных растворов полимераз и в принципе важного значения для этого опыта не имеют, поэтому их можно расценивать как 3 повтора. Индекс «чдо
» у одного из Sозначает ПЦР-смесь до амплификации. В карман геля наносили по 5 мкл образца. М – маркер – смесь нуклеиновых кислот размером от 50 до 1000 п.н.
На рис. видно, что амплификация прошла во всех пробах системы TaqMan. Шесть полос, проявившихся на геле чуть ниже уровня полосы маркера с цифрой 100 означают, что во всех этих пробах прошла реакция и наработался фрагмент ДНК размером около 100 п.н. На дорожках, куда вносили пробы после амплификации в «Терцике» полосы проявились сильнее, чем на дорожках с пробами из чипа. Значит наработанного фрагмента в плашках меньше, чем в макрообъеме под маслом. Это обстоятельство опять же объясняется форматом микрообъемов, где погрешности каждого из этапов работы становится критичным. В случае с SYBRgreen также наблюдается подобная картина. Полоски не проявились в случае с образцом не прошедшим амплификацию и проявились на дорожках лунок, куда вносили образцы после амплификации. Следовательно амплификация в них прошла и наработался фрагмент размером около 600 п.н.
Гель фотографировали с зеленым светофильтром, пропускающим соответствующую длину волны красителя (520 нм). На геле хорошо заметны зеленые полосы крашенной наработанной в процессе амплификации ДНК и исходной матрицы(полосы в самом начале карманов), которая имеет большой размер (5000 п.н.) и за время разделения успела только войти в гель. Опять же количество наработанного фрагмента в чипе меньше (хоть и не значительно по сравнению с системой TaqMan), чем в макрообъемной ПЦР-смеси. Однако нужно обратить внимание на то, что верхняя полоса в лунке S18
ч окрашена слабее, чем в лунке S18
а. Это говорит о том, что изначально количество матрицы в образце было меньше, что опять же связано с погрешностями в работе с микрообъемами.
Выводы.
· Созданы экспериментальные образцы микропланшетов, позволяющие проводить полимеразную цепную реакцию в общем объеме ПЦР-смеси – 1 мкл.
· Подобраны условия эффективной полимеразной цепной реакции в объеме реакционной смеси 1 мкл.
· Отработаны условия ПЦР в микрообъеме (1 мкл) – формате биологических микрочипов.
Заключение.
Сейчас без преувеличения можно сказать: развитие "технологии манипулирования с наследственной информацией" сравнимо по темпам роста с микроэлектроникой и информатикой, нередко эффективно сочетается с ними. И только уникальное сочетание преимуществ традиционных методов и технологий приведет человека к решению ранее недоступных задач в этой интересной и перспективной области. Подобные направления позволяют устанавливать структуру и изучать процессы на уровне генов всего генома, белков всей клетки или клеток всей ткани.
В ходе выполненной работы создана система, которая позволила объединить преимущества таких методов как RealTimeПЦР и микрочипового анализа. Внедрение основы этой разработки в медицину может положить начало создания уникальных тест систем и инструментов клинической диагностики.Количественный ПЦР особенно необходим для обнаружения и мониторинга некоторых вирусных заболеваний ( таких как вирусный гепатит С и В, СПИД). Биочип является уникальным устройством, позволяющим проводить одновременно большое количество анализов на одной подложке. Перевод количественного ПЦР в формат чипа сделает анализ быстрым и экономичным.
Список литературы
1. А.Шляпникова, Ю.М.Шляпников, В.Н.Афанасьев, Г.В.Афанасьева, А.В.Гаврюшкин, И.П.Белецкий. «Исследование качества амино-модифицированных подложек, используемых для гибридизационного анализа» Институт теоретической и экспериментальной биофизики РАН. УДК 547.963.3
2. Шляпникова Е.А., Шляпников Ю.М., Грановский И.Э., Афанасьева,Г.В., Гаврюшкин А.В., Белецкий И.П «Биологические микрочипы в медицине» Институт теоретической и экспериментальной биофизики РАН.
3. Шишкина И.Г., Левина А.С., Зарытова В.Ф. «Аффинные сорбенты, содержащие нуклеиновые кислоты и их фрагменты» // Успехи химии//2001 г. №70(6), с.581,.
4. Иванов, И., Ершов Г., Барский, В., Бельговский, А., Кириллов, Е., Крейндлин, Э., Паринов, С., Мологина, Н., и Мирзабеков А., Диагностика генетических мутаций на олигонуклеотидных микрочипах. (1997 г.) Мол. Биол., 31, с. 159-167.
5. «Биочипы в биологии и медицине XXI века» Академик А.Д. Мирзабеков. Выступление на научной сессии общего собрания РАН
6. Барский В., Колчинский А., Лысов Ю., Мирзабеков А. Биологические микрочипы, содержащие иммобилизованные в гидрогеле нуклеиновые кислоты, белки и другие соединения: свойства и приложения в геномике // Мол. биол., 2002. Т. 36. С. 563-584.
7. Новейшие технологии в генодиагностике: полимеразная цепная реакция в реальном времени (Real-Time PCR). Екимов А.Н., Шипулин Г.А., Бочкарев Е.Г. Рюмин Д.В. Центральный научно-исследовательский институт эпидемиологии Министерства Здравоохранения РФ.
8. Л. И. Патрушев «Искусственные генетические системы» Том 1 Москва, Наука, 2004 С.228.
9. Момыналиев К.Т., Говорун В.М. «Методы развития ДНК-диагностики в клининеческой медицине» Лаборатория генной инженерии и иммуногенетики НИИ физико-химической медицины МЗ РФ, Москва 2003
10. Ekins R.P. (1987) An overview of present and future ultrasensitive nonisotopic immunoassay development. Clin. Biochem. Revs. 8, 12-22.
11. Bernard P.S., Pritham G.H., Wittwer C.T. Color multiplexing hybridization probes using the apolipoprotein E locus as a model system for genotyping. //Analytycal Biochemistry.-1999. - № 273.- p. 221-228.
12. Coutlee, F., He Y., Saint-Antoine P., Olivier C. , Kessous A. Coamplification of HIV type 1 and beta-globin gene DNA sequences in a nonisotopic polimerase chain reaction assay to control for amplification efficiency.// AIDS Res. Hum. Retroviruses.-1995.-№ 11.- p.363-371
13. Heid C.A. Real-time quantitative PCR. // Genome Res.-1996.-№ 6.-p. 986-994.
14. Higuchi R. Kinetic PCR Analysis: Real-time monitoring of DNA amplification reactions// Biotechnology.-1993.-№ 11.- 1026-1030.
15. Sykes P.J., Neoh S.H., Brisco M.J., Hughes E., Condon J., Morley А.А. Quantitation of targets for PCR by use of limiting dilution.// BioTechniques.-1995.-№ 13.- p.444-449.
16. Tyagi S., Kramer F.R. Molecular beacons: probes that fluoresce upon hybridization.// Nat Biotechnol.-1996.-№ 14.- p.303-308.
17. Timofeev E., Mirzabekov A. (1998) Immobilization of DNA in polyacrylamide gel for the manifacture of DNA and DNA-oligonucleotide microchips and protein microchips. Anal.Biochem., 259, 34-41.
18. Adessi C., Matton G., Ayala G., Turcatti G., Mermod J., Mayer P. , Kawashima E. Solid phase DNA amрlification: characterisation of primer attachment and amplification mechanism //Nucleic Acids Research. – 2000. - Vol. 51. - No. 20.
19. Bing D., Boles C., Rehman F., Audeh M., Belmarsh M., Kelley B., Adams C.. Bridge amplification: a solid phase PCR system for the amplification and detection of allelic differences in single copy genes //Genetic Identity Conference Proceedings, Seventh International Symposium on Human Identification. - 1996.
20. Brookes A., Lehvaslaiho H., Siegfried M., Boehm J., Yuan Y., Sarkar C., Bork P., Ortigao F. HGBASE: a database of SNPs and other variations in and around human genes //Nucleic Acids Res. – 2000. – Vol. 28. - № 1- pp. 356-60.
21. Mercier J., Slater G. Solid Phase DNA Amplification: A Brownian Dynamics Study of Crowding Effects //Biophysical Journal.- 2005. – Vol. 89.
|