Кафедра селекции, семеноводства, хранения и переработки сельскохозяйственной продукции
КОНТРОЛЬНАЯ РАБОТА №1
по дисциплине “Физиология растений”
Выполнил:
студент III курса заочного отделения
агрономического факультета
Проверила:
Вопросы
1. Физико-химические свойства цитоплазмы. Гетерогенность цитоплазмы.
2. Ферменты, их химическая природа и функциональное значение. Факторы, влияющие на активность ферментов.
3. Активная роль корневой системы в поглощении воды и минеральных веществ.
4. Действие недостатка воды на растение.
5. Фотосинтез, его значение. Современные представления о сущности фотосинтеза.
6. Каротиноиды, их физиологическая роль.
7. Химизм аэробной фазы дыхания. Заслуга Г. Кребса.
1. Физико-химические свойства цитоплазмы. Гетерогенность цитоплазмы
Кинетика химических реакций в цитоплазме обусловливается сложным сочетанием факторов, среди которых структурные особенности протоплазмы имеют большое значение. Белки протоплазмы отличаются большими химическими возможностями вследствие отличия их строения, химической природы, гетерополярности и поэтому могут вступать в безграничное количество реакций с различными веществами, которые содержатся в протоплазме или поступают извне. В результате этих реакций может измениться форма макромолекулы, что приведет к изменению ее химической активности. Таким образом, изменчивость свойств белков — важная особенность живого вещества.
Цитоплазма построена по коацерватному типу и представляет сложную коллоидную систему из белковых, углеводных и липидных соединений. В разработанной известным советским ученым А. И.Опариным теории о происхождении жизни на Земле большое значение придается выделению органических веществ, белоксодержащих комплексов в форме коацерватных капель из первичных водных растворов.
Белки относятся к гидрофильным коллоидам. Такими же свойствами обладают и другие соединения, входящие в состав протоплазмы. Коллоидная природа протоплазмы имеет существенное биологическое значение. Благодаря наличию большого количества мельчайших частиц в коллоидных системах развиваются огромные суммарные поверхности, которые играют чрезвычайно большую роль. На них может происходить связывание, адсорбция разнообразнейших активных веществ и прежде всего тех, которые снижают поверхностное натяжение. На мицеллах происходит связывание ферментов и других соединений, адсорбируются различные питательные вещества. Все это создает условия для различных химических реакций.
Кроме рассмотренных свойств белков, наблюдается еще явление денатурации. При этом гидрофильные коллоиды — белки — становятся гидрофобными, теряют стойкость и вследствие этого легко коагулируют. Такая типичная денатурация происходит при нагревании белков. Денатурированные белки, т. е. белки, которые утратили свои естественные свойства (выпали в осадок), имеют способность адсорбировать красители. Внешне белки могут казаться неизменными, но поглощению красителя можно определить начало явления денатурации.
Белковые вещества, как амфотерные соединения, вследствие реакций с электролитами изменяют свой заряд, что отражается на состоянии коллоидной системы, а также на ее растворимости. С электролитами связаны величина и знак заряда биоколлоидов протоплазмы, соотношение между процессами гидратации и дегидратации, коацервации и т. д.
Важную роль во всех этих процессах играет поверхность протоплазмы; она является ареной для осуществления процессов адсорбции и десорбции, что влияет на движение частиц, которое может иметь большую скорость, проходить одновременно в противоположных направлениях и влиять также на свойства самой протоплазмы — вязкость, эластичность, проницаемость и др.
Особенности протоплазмы не дают возможность рассматривать ее как истинно золеобразную жидкость, поскольку она по упругости приближается к гелю. Явление взаимного превращения золя в гель наблюдается на протяжении всей жизнедеятельности клетки. На состояние протоплазмы влияют концентрация водородных ионов, а также соотношение между содержанием одно- и двухвалентных катионов. В присутствии кальция коагуляция белков в протоплазме происходит при более низкой температуре. Свойства протоплазмы обусловливаются сложностью многофазной, полидисперсной, коллоидной системы. Цитоплазма имеет три слоя: внешний — плазмалемма, внутренний — тонопласт и лежащий между ними — мезоплазма. Пограничные слои плазмалеммы и тонопласт вязкие и эластичные, а мезоплазма более текучая и менее эластичная.
Межмицеллярные пространства в протоплазме, кроме воды, содержат еще и липоиды, которые находятся в непрочной связи с некоторыми боковыми цепочками белковых веществ. Эти боковые цепочки заканчиваются одной или двумя гидрофобными группами СН3, обладающими способностью присоединять к себе жиры. Кроме того, молекулы липоидных веществ имеют гидрофильные группы СООН, СОН, NH2, которые определяют способность липоидов взаимодействовать с водой. Следовательно, гидрофобные группы молекул будут ориентированы к плазмалемме, а гидрофильные — к мезоплазме. Липиды способны снижать поверхностное натяжение жидкостей; согласно законам физической химии, они концентрируются главным образом на поверхности.
Во взрослых клетках, которые имеют вакуоли, на внутренней поверхности протоплазмы, граничащей с клеточным соком, также образуется обогащенный липидами внешний слой, аналогичный плазмалемме; одновременно происходит скопление содержащихся в клеточном соке липидов возле поверхности вакуоли, которая граничит с протоплазмой. Поэтому тонопласт богаче липидами, чем мезоплазма. Структура протоплазмы чрезвычайно подвижна, и имеющиеся в протоплазме вещества непрерывно вступают во взаимодействие как друг с другом, так и с органическими веществами или минеральными солями, которые поступают в клетку или вырабатываются протоплазмой.
Так, под влиянием сахара структура протоплазмы может измениться (из золя перейти в гель).
Таким образом, протоплазма является сложной гетерогенной коллоидной структурой, которая включает большое количество различных компонентов. Дисперсной средой является комплексный гидрозоль с высоким содержанием белковых и других макромолекул, Сахаров, неорганических солей, например фосфатов. Важную роль играет вода, которая насыщает всю систему коллоидов протоплазмы, образуя непрерывную фазу.
В живой протоплазме постоянно происходят процессы новообразования и распада различных веществ, коагуляция коллоидов и их обратное превращение в золи, образование коацерватов, гелей и т. д. Происходящие в протоплазме процессы непосредственно зависят от состояния и свойств структур, из которых она состоит. Изменения протоплазменных структур под воздействием внешних условий имеют приспособительный характер.
2. Ферменты, их химическая природа и функциональное значение. Факторы, влияющие на активность ферментов
Ферменты подразделяются на одно- и двухкомпонентные. Первые состоят только из молекул белка, вторые из белковой части, получившей название апофермента, и соединения небелковой природы, называемой простетической группой. В двухкомпонентных ферментах белок-носитель называют еще фероном, а небелковую активную группу — агоном. У двухкомпонентных ферментов, небелковая часть которых легко отделяется от апофермента, проететические группы называют коферментами. Размеры кофермента во много раз меньше размеров белковой молекулы (апофермента).
Коферментами ферментов пиридиновых дегидрогеназ являются:
- никотинамидадениндинуклеотид (НАД);
- никотинамидадениндинуклеотидфосфат (НАДФ);
- флавиннуклеотиды — коферменты флавиновых ферментов;
- производные фолиевой кислоты — коферменты фермента глицинтрансформиминазы.
Специфическая деятельность ферментов является одним из важнейших факторов организации процессов обмена веществ в живом организме, их согласованности и направленности.
Многочисленные биологические катализаторы в организме — ферменты, отличающиеся исключительной специфичностью и эффективностью действия, ускоряют только определенные превращения данного вещества.
Распределение скоростей ферментативных превращений, которые создаются в организме, в значительной мере определяет специфичность процессов обмена веществ.
Действия ферментов согласованны: продукты катализа одного фермента поступают к другому, а не рассеиваются в содержимом клетки; реакции, выделяющие энергию, тесно связаны с реакциями, требующими затрат энергии. Установлено, что окисление сахаров и жиров происходит с выделением энергии, тогда как синтез белков требует затрат ее.
Активность ферментов зависит от условий внешней среды, окружающей молекулу фермента. Каждый фермент работает в определенных границах температуры и рН. Для большинства ферментов температурный оптимум лежит в пределах 40—50°С. В клетке одновременно работает множество ферментных систем, и каждый фермент требует определенной реакции среды.
Наивысшая активность большинства растительных ферментов отмечается при слабокислой или нейтральной реакции, характерной для растительных клеток. Такое влияние рН объясняется непосредственным действием концентрации водородных ионов на свойства центра, определяющие образование фермент-субстратного комплекса. Кроме того, ионы водорода оказывают влияние на степень ионизации субстрата и молекулы ферментного белка.
Скорость ферментативной реакции в сильной степени зависит от концентрации субстрата в среде, но если достаточно субстрата, то и от содержания фермента.
Активность ферментов в сильной степени зависит от содержания в реакционной среде различных дополнительных ионов и соединений. Вещества или ионы, увеличивающие каталитическую активность ферментов, получили название активаторов.
Роль активаторов ферментов выполняют ионы различных металлов: К+, Са2+, Mg2+, Fe2+, Cа2+ и др. Активация может осуществляться одним или несколькими ионами. Например, амилаза, катализирующая расщепление крахмала и липаза, — распад жиров, активируются ионами Са2+; алкогольдегидрогенеза, катализирующая окисление спиртов до альдегидов, -Zn2+; пероксидаза и каталаза — Fe2+; аргиназа, участвующая в гидролитическом расщеплении аргинина, — Со2+; Mn2+; Ni2+.
Для проявления максимальной активности фермента требуется определенная концентрация ионов-активаторов в среде.
Усиление активности ферментов под действием ионов объясняется прежде всего тем, что многие ферменты содержат их в своей молекуле и представляют собой так называемые металлоферменты.
К металлоферментам относятся каталаза и пероксидаза, содержащие железо, амилаза, в состав которой входит кальций; нитратредуктаза, содержащая молибден. В ряде случаев активность ферментов в присутствии ионов металлов поддерживается образованием координационных связей между активным центром фермента и субстратом.
Существуют вещества, подавляющие действие ферментов, которые получили название ингибиторов ферментов. Ингибиторы делят на два класса: общие и специфические. К общим ингибиторам относят соли тяжелых металлов — свинца, серебра, ртути, вольфрама, которые денатурируют белок и, следовательно, подавляют действие ферментов.
Наибольшее значение имеют специфические ингибиторы, которые находят практическое применение. Они действуют только на одну ферментативную реакцию. Присоединяясь к активному центру фермента, ингибитор препятствует образованию комплекса фермент — субстрат, вследствие чего часть молекул фермента переходит в неактивное состояние и скорость ферментативной реакции замедляется или прекращается.
3. Активная роль корневой системы в поглощении воды и минеральных веществ
Вследствие интенсивного расхода воды, особенно в процессе транспирации, растения нуждаются в активном ее поступлении. Для этого существует корневая система. У хлебных злаков корневая система проникает на глубину 1,5-2 м и распространяется широко в стороны. В зависимости от вида растений распределение корневой системы в почве различно.
С физиологической точки зрения, корневая система неоднородна. Далеко не вся поверхность корня участвует в поглощении воды.
В первичном строении корня (рис. 2.3) различают несколько тканей: корневой чехлик, апикальную меристему, ризосферу, первичную кору, эндодерму, перицикл и проводящие ткани, сосредоточенные в центральном цилиндре. Растущая часть корня обычно не превышает 1 см в длину и состоит из меристемы (1,5-2 мм от кончика) и зоны растяжения (2-7 мм).
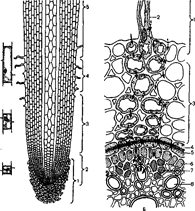
Рис. 2.3. Схема строения корня:
А — продольный разрез: 1 — корневой чехлик; 2 — меристема; 3 — зона растяжения; 4 — зона корневых волосков; 5 — зона ветвления; Б — поперечный разрез (по М. Ф. Даниловой): 1 — ризодерма; 2 — корневой волосок; 3 — паренхима; 4 — эндодерма; 5 — пояски Каспари; 6 — перицикл; 7— флоэма; 8 — ксилема.
Стрелки — пути передвижения веществ, поглощаемых из наружного раствора. Сплошные стрелки — путь раствора по симпласту; прерывистые — по апопласту.
На самом кончике корня находится корневой чехлик, клетки наружных слоев которого секретируют слизь (полисахаридную) и слущиваются при перемещении корня в почве. Клетки центральной части чехлика (статоциты) содержат много амилопластов, заполненных крахмалом (статолиты) и участвующих в восприятии корнем направления действия силы тяжести. Основными функциями корневого чехлика являются: а) предохранение апикальной меристемы корня от повреждения при соприкосновении с почвой; б) облегчение продвижения кончика корня в почве; в) восприятие геотропического стимула.
Апикальные клетки меристемы корня (1-2% от общего числа клеток в меристеме) отличаются от остальных значительно более редким делением, медленным синтезом ДНК и РНК и белков, меньшим числом плазмодесм с соседними клетками. Они получили название покоящегося центра. При повреждении апикальной меристемы фонд ее клеток восстанавливается за счет деления клеток покоящегося центра.
За время жизни в меристеме корня каждая клетка делится 6-7 раз. Количество клеток увеличивается вдоль оси корня, формируется перицикл.
Прекратившие деление, клетки корня переходят к растяжению также в направлении оси корня. Это приводит к пространственному разобщению делящихся клеток меристемы и клеток, растущих растяжением. В зоне растяжения заканчивается дифференцировка флоэмы и формируются элементы протоксилемы.
Дифференцировка тканей корня заканчивается в зоне корневых волосков, где завершается образование основных тканей корня (рис. 2.2): ризодермы, первичной коры, эндодермы и системы тканей центрального цилиндра.
Ризодерма — это чаще всего однослойная ткань, покрывающая корень снаружи. У большинства видов растений клетки ризодермы обладают способностью формировать корневые волоски.
При дальнейшем увеличении возраста клеток, а также расстояния от кончика корня, корневые волоски исчезают, начинается кутинизация и опробковение клеточных стенок. Поглощение воды происходит главным образом клетками зоны растяжения и зоны корневых волосков.
Рассмотрим строение корня в зоне корневых волосков.
Поверхность корня этой зоны покрыта ризодермой. Это однослойная ткань с двумя видами клеток: формирующими и не формирующими корневые волоски. Клетки, формирующие корневые волоски, отличаются особым типом обмена веществ. Корневые волоски растут путем растяжения клеточной оболочки, которое происходит с большой скоростью (0,1 мм/час). Для их роста необходим кальций.
За ризодермой до перицикла идут клетки коры. Кора состоит из нескольких слоев паренхимных клеток. Важной особенностью коры является развитие системы крупных межклеточников. На границе коры и центрального цилиндра развивается один слой плотно прилегающих друг к другу клеток — эндодерма, для которой характерно наличие поясков Каспари. Цитоплазма клеток эндодермы плотно прилегает к клеточным оболочкам. По мере старения вся внутренняя поверхность клеток эндодермы, за исключением пропускных клеток, покрывается суберином. Поэтому клетки эндодермы являются основным физиологическим барьером для передвижения воды.
В центральном цилиндре расположены проводящие ткани корня. Сосуды ксилемы контактируют с паренхимными клетками центрального цилиндра, которые активно транспортируют ионы в проводящие элементы ксилемы. Контакт осуществляется через полуокаймленные поры во вторичной клеточной стенке со стороны сосуда и поры со стороны паренхимной клетки. Между этими клетками нет плазмодесм.
4. Действие недостатка воды на растение
Влияние засухи на растение.
Если в почве имеется недостаточное количество доступной для растения воды, то отрицательное влияние перегрева становится особенно сильным. Перегрев вызывает повреждение растения, называемое запалом. Запал обнаруживается через некоторое время в виде различно окрашенных некротических пятен на листьях. На пшенице появляются желтые пятна, на овсе — красные, у большинства растений — коричневые.
Встречается и другой вид повреждений от атмосферной засухи — захват. Он наблюдается реже, чем запал, и проявляется в том случае, когда при сравнительно не очень высоких температурах наблюдаются сильный ветер и большая сухость воздуха. При этом листья просто высыхают, сохраняя зеленую окраску.
Недостаток воды в тканях растений (водный дефицит) может возникнуть в жаркую солнечную погоду к середине дня, при этом увеличивается сосущая сила листьев, что активирует поступление воды из почвы. Растение регулирует уровень водного дефицита открытием или закрытием устьиц. В этот период происходит временное завядание листьев. Обычно в вечерние и утренние часы это явление устраняется.
Отсутствие в почве доступной для растения воды приводит к глубокому завяданию. Это завядание чаще всего приводит к гибели растения. Характерным признаком устойчивого водного дефицита является сохранение его в тканях утром, прекращение выделения пасоки из срезанного стебля. Действие засухи приводит в первую очередь к уменьшению в клетках свободной воды, что нарушает гидратные оболочки белков цитоплазмы и сказывается на функции белков-ферментов.
При длительном завядании снижается активность ферментов синтеза и активируются гидролитические процессы, что приводит к возрастанию содержания в клетках низкомолекулярных белков. В результате гидролиза полисахаридов в тканях накапливаются растворимые углеводы. В листьях снижается количество РНК, наблюдается распад полирибосомных комплексов, возрастает концентрация вакуолярного сока, облегчается выход ионов из клеток.
Происходит снижение скорости фотосинтеза из-за недостатка СО2, нарушения синтеза хлорофилла и АТФ, изменения в течение фотохимических реакций и задержки оттока ассимилятов из листьев.
При обезвоживании у растений, не приспособленных к засухе, значительно усиливается интенсивность дыхания, а у засухоустойчивых растений такое явление не наблюдается.
В условиях водного дефицита тормозятся клеточное деление и особенно растяжение, что приводит к формированию мелких клеток. Вследствие этого задерживается рост листьев, стебля растения в целом.
При водном дефиците происходит гидролиз полимеров, в том числе и белков. Распад белков идет с образованием аммиака, который может оказывать отравляющее действие на клетки.
Приспособление растений к засухе. Известно, что растения неодинаково реагируют на перегрев и обезвоживание в разные периоды онтогенеза. У каждого вида в онтогенезе имеется такой период, когда недостаток воды резко сказывается на течении всех физиологических процессов, этот период называется критическим периодом. Из этого, однако, не следует, что остальные периоды своего развития растение не нуждается в воде и не страдает от ее недостатка.
Установлено, что вязкость цитоплазмы, ее эластичность в критический период резко падают, что и является одним из условий высокой чувствительности растений к перегреву и обезвоживанию в этот период развития. В критический период происходят интенсивные ростовые процессы и образование новых органов — цветков.
Ф.Д. Сказкин считал, что высшие растения в своем онтогенезе повторяют черты далекого прошлого и во время формирования половых органов и оплодотворения нуждаются в повышенной обводненности тканей. Сказкин считает, что началом критического периода следует считать момент появления материнских клеток пыльцы в археспориальной ткани пыльников, а концом данного периода — оплодотворение.
У растений засушливых местообитаний — ксерофитов — вырабатывались приспособления, позволяющие переносить засухи. Группа ксерофитов очень разнообразна. По способности переносить условия засухи различают следующие их типы (по П.А. Генкелю):
5. Фотосинтез, его значение. Современные представления о сущности фотосинтеза
Фотосинтез — это процесс преобразования энергии света в химическую энергию органических соединений. Синтезированные органические соединения служат основным источником энергии для всей биосферы. Процесс синтеза органического вещества за счет углерода в живой природе осуществляется с помощью фотосинтеза растений, бактериального фотосинтеза и хемосинтеза.
Ведущее значение фотосинтеза может характеризоваться следующими данными. В период интенсивного роста суточные приросты сухой массы составляют в среднем 150 кг, а в лучших случаях — 500 кг на 1 га. При этом корни усваивают в виде ионов 2 кг азота, 0,5 кг фосфора, 4 кг калия и 4 кг других элементов. За это же время листья усваивают 1000 кг СО2. При средней урожайности корней сахарной свеклы 300 ц с 1 га растения за вегетационный период усваивают 150 кг азота, 30 кг фосфора, 160 кг калия и 4200 кг углерода, что соответствует поглощению 20 т СО2. В урожае аккумулируется 167,2 ГДж энергии. Это необходимо иметь в виду при обосновании целей и задач земледелия, которое представляет собой систему использования основной функции зеленых растений — фотосинтеза.
Фотосинтез растений мог возникнуть лишь на определенном этапе существования и развития жизни на Земле как дальнейшее усложнение и развитие менее совершенных форм фиксации углекислого газа. Очевидно, типы ассимиляции СО2 в процессе эволюции обмена веществ развивались от гетеротрофного к автотрофному (фотосинтезу) через хемотрофию, фоторедукцию. Фотосинтез осуществляют высшие растения, водоросли и некоторые бактерии.
Английский ботаник и химик С. Гейлс (1727) высказал предположения, что растения значительную часть «пищи» получают из воздуха. Подобные идеи высказывал и российский ученый, М. В. Ломоносов. В 1753 году, изучая различные воздушные явления, М. В. Ломоносов писал: «Преизобильное орошение тучных дерев, которые на бесплодном песку корень свой утверждали, ясно изъявляет, что листами жирный тук из воздуха впитывают...». Так возникла идея о воздушном питании растений.
Началом экспериментальных работ в области фотосинтеза послужили опыты английского химика Дж. Пристли. В 1771 году он обнаружил, что растения мяты, помещенные в стеклянный кувшин, опрокинутый в сосуд с водой, «исправляют» в нем воздух, «испорченный» горением свечи или дыханием мыши. В связи с этим выводом и последующими опытами Пристли в 1774 году открыл кислород. Через год независимо от него кислород был открыт во Франции А. Л. Лавуазье, который и дал название этому газу.
В 1766 году шведский химик К. В. Шееле повторил опыты Пристли, но не получил того же эффекта, который был описан Пристли. Объяснил противоречие голландский врач Я. Ингенхауз, который доказал, что зеленые растения выделяют кислород только на свету.
Швейцарский исследователь Ж. Сенебье в 1782 году установил, что растения на свету поглощают СО2, и назвал это явление «углеродным питанием».
В 1804 году швейцарский исследователь Т. Соссюр установил, что растения на свету поглощают СО2 и воду и выделяют кислород, и сделал вывод, что органическая масса растений образуется за счет СО2 и воды, т. е. вода является необходимым элементом питания.
Французские химики П. Ж. Пелетье и Ж. Кавенту в 1817 году выделили из листьев зеленый пигмент и назвали его хлорофиллом. В 1865 году немецкий физиолог Ю. Сакс продемонстрировал, что на свету в листьях образуется крахмал и он находится в хлоропластах. Сам процесс обнаружения крахмала в листьях стали называть «пробой Сакса».
Первые опыты по выяснению роли листа образования кислорода при фотосинтезе были сделаны немецким физиологом Т. В. Энгельманом (1881). Было установлено, что кислород на свету выделяют хлоропласты.
Значением света в процессе фотосинтеза занимался американский физик Дж. У. Дрепер (1846), который вслед за Ю. Саксом и В. Пфеффером считал, что фотосинтез лучше протекает в желтых лучах. Но уже в то время было известно, что хлорофилл имеет два максимума поглощения в красной и синей частях спектра. Исходя из этих данных, делает вывод, что процесс фотосинтеза не подчиняется закону сохранения энергии, а свет действует как раздражитель. Против этого утверждения выступили Ю. Р. Майер и Г. Гельмгольц, сформулировавшие закон сохранения и превращения энергии. На основании теоретических рассуждений они утверждали, что зеленые растения поглощают лучистую солнечную энергию и превращают ее в химическую, т. е. фотосинтез — это процесс трансформации энергии в энергию химических связей.
Экспериментальные доказательства этой точки зрения были получены российским физиологом К. А. Тимирязевым в 1875 году. Тимирязев постулировал, что при ассимиляции СО2 хлорофилл служит оптическим сенсибилизатором (т. е. веществом, увеличивающим чувствительность к свету) и что он непосредственно участвует в процессе фотосинтеза, необратимо переходя из восстановленного состояния в окисленное. Он сформулировал идею о космической роли фотосинтеза.
Таким образом, к концу 19-го столетия сложилось представление об общем уравнении фотосинтеза.
6. Каротиноиды, их физиологическая роль
Кроме хлорофиллов, в клетках листа в небольшом количестве (0,07—0,2 % сухой массы) содержатся каротиноиды.
Это соединения, растворимые в жирах и органических растворителях, с длинной цепью сопряженных двойных связей.
Поглощая коротковолновые лучи, каротиноиды исполняют роль экрана, защищающего хлорофилл и клетку от их губительного воздействия. Это большая группа пигментов желтого, оранжевого и красного цвета. Каротиноиды широко распространены в природе: их обнаружено больше трехсот. Однако в фотосинтезе участвуют лишь некоторые из них.
Термин «каротиноиды» ввел в науку М.С. Цвет для желтых спутников хлорофилла, т. е. веществ, сопровождающих хлорофилл в пластидах. В пыльце растений, околоцветников каротиноиды находятся во внепластидном состоянии. Каротиноидов в хлоропластах обычно в три раза меньше, чем хлорофиллов (отношение хлорофиллов к каротиноидам равно 3).
Основная часть молекул всех каротиноидов — восемь соединенных между собой остатков изопрена, образующих цепь. Каротиноиды (рис. 3.8) делятся на соединения с открытой цепью — ациклические (например, ликопин) и содержащие в своей молекуле один или два цикла (α-, β-каротин). Кроме того, по химическому строению каротиноиды делятся на бескислородные, содержащие только углерод и водород, имеющие элементарную формулу С40Н56 каротины, а также окисленные, которые кроме углерода и водорода содержат еще кислород — ксантофиллы.
Каротиноиды растворимы в органических растворителях, такие, как сероуглерод и ацетон, но нерастворимы в воде (кроме кроцина). Многие каротиноиды растворимы в спирте, петролейном эфире, хлороформе, они хорошо растворяются в жирах.
В химическом отношении молекулы каротиноидов характеризуются большим числом сопряженных двойных связей, являющихся хромофорными группами. Каротиноиды поглощают свет в сине-фиолетовой области спектра и имеют три максимума, положение которых определяется главным образом количеством двойных связей. Флюоресцируют каротиноиды от слабо желтой до зеленой окраски. В месте нахождения двойных связей может легко происходить присоединение кислорода, в результате чего образуются перекиси и окраска исчезает. К группе бескислородных каротиноидов относятся каротины: ликопин, содержащийся в плодах томатов. К окисленным каротиноидам относятся ксантофиллы: лютеин, криптоксантин, содержащийся в желтых зернах кукурузы, кожуре мандаринов и др.
Каротиноиды имеют большое количество конъюгированных двойных связей, поэтому они способны к окислительно-восстановительным реакциям.
Поглощение света каротиноидами, а, следовательно, их окраска обусловлены наличием конъюгированных двойных связей, β-каротин имеет два максимума поглощения, соответствующие длинам волн 482 и 452 нм. Красные лучи, поглощаемые хлорофиллами, каротиноидами не поглощаются. Каротиноиды в отличие их от хлорофилла не обладают способностью к флюоресценции. Подобно хлорофиллу, каротиноиды в хлоропластах вступают во взаимодействие с белками.
Уже сам тот факт, что каротиноиды всегда присутствуют в хлоропластах, позволяет считать, что они принимают участие в процессе фотосинтеза, но их роль вспомогательная. Они поглощают определенные участки спектра света и передают энергию на хлорофилл, одновременно защищая молекулу хлорофилла от необратимого фотоокисления. Возможно, каротиноиды принимают участие в кислородном обмене при фотосинтезе. У высших растений, мхов, зеленых и бурых водорослей осуществляется светозависимое взаимопревращение ксантофиллов.
7. Химизм аэробной фазы дыхания. Заслуга Г. Кребса
Вторая фаза дыхания — аэробная — требует присутствия кислорода. В аэробную фазу вступает пировиноградная кислота. В присутствии достаточного количества О2
пируват полностью окисляется до СО2 и Н2О.
Еще в 1910 году шведским химиком Т. Тунбергом было доказано участие органических кислот в процессе дыхания. В 1935 году А. Сент-Дьерди в Венгрии установил, что добавление органических кислот к живой ткани резко активирует поглощение ею кислорода.
В 1937 году Г. А. Кребс предложил схему последовательности окисления три- и дикарбоновых кислот до СО2 через «цикл лимонной кислоты» за счет отнятия водорода. Этот цикл и был назван его именем. (Рис. 4.3).
В цикле окисляется не сама пировиноградная кислота, а ее производное — активный ацетат. Таким образом, первым этапом окислительного расщепления пировиноградной кислоты (ПВК) является процесс образования активного ацетата в ходе окислительного декарбоксилирования. Окислительное декарбоксилирование ПВК осуществляется при участии сложной ферментной системы пируватдегидрогеназы. Эта ферментная система включает в себя три фермента и пять коферментов (тиаминпирофосфат, липовая кислота, коэнзим А, ФАД и НАД). В результате этого процесса образуется активный ацетат (ацетил-КоА), восстановленный НАД (НАДН) и выделяется первая молекула СО2.
Ферментная система пируватдегидрогеназа ингибируется АТФ. При накоплении АТФ выше определенного уровня превращение ПВК прекращается. Это один из механизмов регулирования аэробной фазы дыхания.
Образовавшийся в ходе этапа активный ацетат вступает в цикл три- и дикарбоновых кислот.
Второй этап начинается с взаимодействия активного ацетата с енольной формой щавелево-уксусной кислоты, результатом реакции является образование лимонной кислоты. В этой реакции принимает участие фермент цитратсинтетаза.
Следующий этап цикла включает две реакции и катализируется ферментом аконитазой. В первой реакции в результате дегидратации лимонной кислоты образуется цисаконитовая. Во второй реакции цисаконитовая кислота гидратируется в изолимонную. Изолимонная под действием НАД или НАДФ - зависимой изоцитратдегидрогеназы окисляется в нестойкое соединение — щавелево-янтарную кислоту, которая декарбоксилируется с образованием оскетоглутаровой кислоты.
Подобно ПВК, α-кетоглутаровая кислота подвергается реакции окислительного декарбоксилирования с аналогичной мультиферментной системой. В ходе этой реакции α-кетоглутаровая кислота превращается в сукцинил-КоА, образуется НАДН и выделяется одна молекула СО2.
Подобно активному ацетату, сукцинил-КоА является высокоэнергетическим тиоэфиром. Однако если в случае с ацетил-КоА энергия тиоэфирной связи расходуется на синтез лимонной кислоты, энергия сукцинил-КоА может трансформироваться в образовании фосфатной связи АТФ. При участии сукцинил-КоА-синтетазы из сукцинил-КоА, АДФ и Н3РО4 образуется янтарная кислота (сукцинат), АТФ, регенерируется молекула KoA-SH. АТФ образуется в результате субстратного фосфорилирования.
На следующем этапе янтарная кислота окисляется до фумаровой. Реакция катализируется сукцинатдегидрогеназой, коферментом которой является ФАД. Фумаровая кислота под действием фумаразы, присоединяя воду, превращается в яблочную кислоту (малат). И, наконец, на последнем этапе яблочная кислота с помощью НАД-зависимой малатдегидрогеназы окисляется в щавелево-уксусную (ЩУК). А ЩУК вступает в новый цикл.
Следует отметить, что большинство реакций цикла обратимо, однако ход цикла в целом практически необратим. Причина этого в том, что в цикле есть две сильно экзооргические реакции — цитратсинтетазная и сукцинил-КоА-синтетазная.
На протяжении одного оборота цикла при окислении ПВК происходит выделение трех молекул СО2, включение трех молекул Н2О и удаление пяти пар атомов водорода. Роль воды в цикле Кребса подтверждает правильность уравнения Палладина, который отмечал, что дыхание идет с участием Н2О, кислород которой включается в окисляемый субстрат, а водород с помощью «дыхательных пигментов» (по современным представлениям — коферментов дегидрогеназ) переносится на кислород. Большинство ферментов цикла Кребса локализовано (рис. 4.4) в матриксе митохондрий, аконитаза и сукцинатдегидрогеназа — во внутренней мембране митохондрий.
Третий этап цикла Кребса — это электрон-транспортная цепь (ЭТЦ). В процессе окисления пировиноградной кислоты в цикле Кребса образовались пары водорода 2Н, которые мы можем рассматривать как 2Н+ + 2е-. Именно в таком виде они и передаются по цепи переносчиков. Движущей силой транспорта водорода в дыхательной цепи (рис. 4.5) является разность потенциалов. В связи с этим расположение отдельных переносчиков в дыхательной цепи, также как и в цепи фоосинтетической, определяется величиной их окислительно-восстановительного потенциала. В начале цепи расположен НАД, обладающий наибольшей величиной окислительно-восстановительного (ОВ) потенциала (-0,32В), а в конце — кислород с наиболее положительной величиной (+0,82В).
Остальные переносчики расположены между ними в порядке последовательного повышения положительного потенциала. Это и позволяет электронам передаваться по направлению к кислороду.
Б. Чане и др. (США) в 50-х годах предложили следующее расположение переносчиков в ЭТЦ:
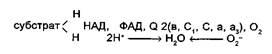
Пара электронов от НАДН или сукцината передается по ЭТЦ до кислорода, который, восстанавливаясь и присоединяя два протона, образует воду.
Окислительное фосфорилирование.
Перенос электронов от НАДН к молекулярному кислороду через ЭТЦ митохондрий сопровождается потерей свободной энергии.
В. А. Энгельгардом еще в 30-х годах было показано, что при аэробном дыхании накапливается АТФ. Он первый высказал идею о сопряжении между фосфорилиро-ванным АДФ и аэробным дыханием. В 1937—1939 гг. биохимики В. А. Белицер в СССР и Г. Калькар в США установили, что при окислении промежуточных продуктов цикла Кребса, в частности янтарной и лимонной кислот, образуется АТФ. В анаэробных условиях фосфорилирования не происходит.
Процесс фосфорилирования АДФ с образованием АТФ, сопряженный с переносом электронов по ЭТЦ митохондрий, получил название окислительного фосфорилирования. В ЭТЦ синтезируется три молекулы АТФ. Так, первая молекула АТФ образуется при переходе пары электронов от пиридиновой к флавиновой дегидразе. Вторая — при переносе пары электронов от цитохрома б к цитохрому с, и третья — при переносе пары электронов от цитохрома а к цитохромоксидазе (а3).
Места образования АТФ в дыхательной цепи не являются случайными. Именно на этих участках выделяется достаточное для образования молекул АТФ количество энергии.
Показателем сопряженности окисления и фосфорилирования служит коэффициент фосфорилирования Р/О. В данном случае коэффициент Р/О = 3. Существуют вещества, разобщающие процесс окисления и фосфорилирования (динитрофенол, соли тяжелых металлов). Вся энергия в данном случае превращается в тепло, а коэффициент Р/О = 0. Такое явление может наблюдаться и при засухе или заболевании организма.
По поводу механизма окислительного фосфорилирования существует три теории: химическая, конформационная и хемиосмотическая.
В настоящее время наибольшим признанием пользуется хемиосмотическая теория английского биохимика Н. Митчелла (1961). Он высказал предположение, что поток электронов через систему молекул-переносчиков сопровождается транспортом ионов Н+ через внутреннюю мембрану митохондрий. В результате на мембране создается электрохимический потенциал ионов Н+, включающий химический или осмотический градиент и электрический градиент (мембранный потенциал). Согласно хемиосмотической теории, электрохимический трансмембранный потенциал ионов Н+ и является источником энергии для синтеза АТФ за счет обращения транспорта ионов Н+ через протонный канал мембранной Н+ АТФ-азы.
Теория Митчелла исходит из того, что переносчики перешнуровывают мембрану, чередуясь таким образом, что в одну сторону возможен перенос и протонов, и электронов, а в другую — только электронов. В результате ионы Н+ накапливаются на одной стороне мембраны. Энергия запасенная, таким образом, используется для синтеза АТФ как результат разрядки мембраны при обратном транспорте протонов через АТФ-азу.
Список используемой литературы
1. Алешин Е.П., Пономарёв А.А. Физиология растений. – 2-е изд., перераб. и доп. – М.: Агропромиздат, 1985. – 255 с., ил. - (Учебники и учеб. пособия для сред. с.-х. учеб. заведений).
2. Кириллов Ю.И., Кокин Г.А. Физиология растений: Учебное пособие. Курган, издательство «Зауралье», 1998, 304 с. ил.
3. Лебедев С.И. Физиология растений. – 2-е изд., перераб. и доп. – М.: Колос, 1982. – 463 с., ил. – (Учебники и учеб. пособия для высш. с.-х. учеб. заведений).
|