Реферат
по биологии
на тему:
"Эмбриональные стволовые клетки в изучении функции генов в процессах дифференцировки и развития"
2009
Введение
Сообщение о получении культуры ЭС клеток мыши было сделано одновременно в двух работах в 1981 г.. В настоящее время ЭС клетки подразделяют на три типа.
1. Клетки, изолированные из внутренней клеточной массы бластоцисты млекопитающих, именно эти клетки принято называть эмбриональными стволовыми клетками.
2. Клетки эмбриональных карцином.
3. Первичные половые клетки зародыша.
На рис. представлена общепринятая в настоящее время схема получения ЭС клеток и их использование для получения химерных животных. В последние годы ЭС клетки используются в различных направлениях исследований.
Кроме экспериментов по получению химерных животных и направленному изменению генов с целью изучения их функций в процессе развития и на уровне взрослого организма, ЭС клетки используются в экспериментах по клонированию животных и изучению начальных стадий эмбрионального развития invitro. Большой интерес в последние годы вызывает направление, связанное с изучением путей направленной дифференцировки ЭС и стволовых клеток млекопитающих и, что особенно важно человека, в определенные типы клеток invitro. Способность управлять этими процессами позволит в перспективе использовать такие клетки в заместительной терапии различных тяжелых заболеваний человека, таких как нейродегенеративные, онкологические и др.
ЭС клетки обладают высокой пролиферативной активностью и способностью в течение длительного времени в культуре поддерживаться в недифференцированном состоянии. Для сохранения недифференцированного фенотипа ЭС клеток в культуре требуется наличие фидерного слоя, который может быть представлен первичными эмбриональными фибробластами или перевиваемыми фибробластами мыши линии STO.
На рис. показан рост ЭС клеток на фидерном слое эмбриональных фибробластов мыши. Другой метод предупреждения дифференцировки ЭС клеток включает добавление в культуральную среду в отсутствие фидера LIF, который представляет собой гемопоэтический регулятор, индуцирующий дифференцировку клеток миелоидной лейкемии линии Ml. Успешно используются и ряд других цитокинов для сохранения недифференцированного состояния ЭС клеток: интерлейкин-6, онкостатин, цилиарный нейротрофический фактор. В качестве методических приемов предупреждения дифференцировки ЭС клеток используются: добавление в среду LIF в присутствии фидерного слоя, фидерные клетки, трансформированные рекомбинантным вектором и экспрессирующие LIF, и применение рекомбинантного LIF в отсутствие фидерного слоя.
Все описанные линии ЭС клеток мыши сходны по своим морфологическим характеристикам. Клетки имеют крупное ядро, содержащее преимущественно эухроматин и несколько ядрышек. Для них свойственно высокое ядерно-цитоплазматическое отношение: цитоплазма представлена узким ободком, окружающим ядро. Клеточный цикл 18—20 ч, П,-фаза очень короткая. Кариотипический анализ 35 линий ЭС клеток показал, что 25 из них имеют кариотип ХУ, а остальные ХО и XX. Авторы полагают, что ЭС клетки предпочтительней образуются из клеток с мужским кариотипом, поскольку вторая Х-хромосома может явиться дестабилизатором плюрипотентности. В основном все ЭС клетки имеют нормальный кариотип 2п = 40, изменения в кариотипе могут приводить к потере плюрипотентности.
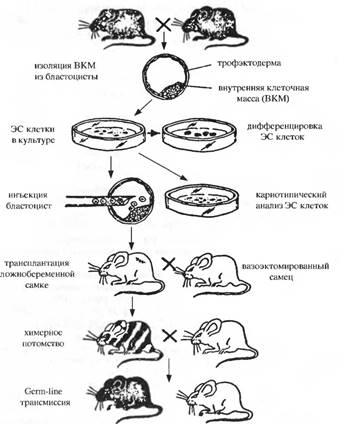
Схема получения ЭС клеток и их использование при изучении процессов направленной дифференцировки в системах invitroи invivo, а также при создании трансгенных животных
Особенностью ЭС клеток является высокая активность эндогенной щелочной фосфатазы. Методом количественного анализа было установлено, что активность эндогенной щелочной фосфатазы стволовых клеток тератокарцином, внутренней клеточной массы бластоцисты, эпибласта 5-6-дневного эмбриона, первичных половых клеток в 10 раз превышает таковую у дифференцированных соматических клеток. Для ЭС клеток характерен также высокий уровень теломеразной активности, который коррелирует со степенью недифференцированности ЭС клеток. Сохранение недифферен цированного фенотипа ЭС клеток в культуре может быть подтверждено использованием антител к специфическим поверхностным антигенам ЭС, таких как ЕСМА-7, SSEA-1.
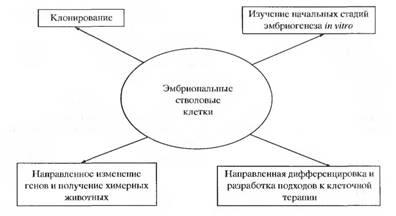
Основные направления исследований, в которых в настоящее время используются эмбриональные стволовые клетки млекопитающих
При определенных условиях культивирования ЭС клетки формируют эмбриоидные тела, которые представляют собой зачатки эндодермы, эктодермы и мезодермы, напоминая постимплантационное эмбриональное развитие. Если эмбриоидные тела прикрепляются к субстрату, то клетки, формирующие внутренние слои таких тел, способны в дальнейшем дифференцироваться в широкий спектор тканей, такие как мышечная, нервная, эпителиальная и др. Способность ЭС клеток к дифференцировке invitroиспользуется как модель для исследования процессов клеточной дифференцировки в разные типы тканей.
После инъекции ЭС клеток в бластоцисты они способны участвовать в образовании всех тканей, включая и линии зародышевых клеток химерного организма. Способность ЭС клеток дифференцироваться invivoоценивается по частоте химер и степени колонизации тканей химерного потомства, а также по частоте генеративных химер. Это свойство клеток в сочетании с методом гомологичной рекомбинации, которая позволяет направленно изменять геном ЭС клеток, дает возможность создавать трансгенных животных с заданным генотипом.
В 1982 г. впервые было показано, что соматические клетки млекопитающих обладают ферментативным механизмом, способным осуществлять гомологичную рекомбинацию между хромосомной и экзогенной ДНК, введенной в соматические клетки. Частота гомологичной рекомбинации достаточно низкая и в зависимости от локуса может быть от 10-3
до 10-7
. Существует две возможности для гомологичной рекомбинации между экзогенной и геномной последовательностями ДНК: замещение и внедрение. В случае замещения происходит двойной кроссинговер между гомологичными последовательностями вектора и генома, в случае внедрения происходит простой кроссинговер. Замещение приводит к инактивации гена, т.е. к его "нокауту".
В первых экспериментах на ЭС клетках для таргетинга был избран ген hprt. Ген hprtлокализован на Х-хромосоме, поэтому мужские ЭС клетки гемизиготны по hprtи достаточно инактивации только одной копии гена для получения селективного фенотипа. Hprtмутанты могут расти в среде, содержащей аналог пурина, 6-тиогуанин, тогда как клетки, имеющие нормальный фермент, на этой среде не растут. Этими авторами впервые был сконструирован вектор, в котором бактериальный ген пео, обеспечивающий устойчивость к антибиотику G418, вставлен в восьмой экзон клонированного фрагмента гена hprt. Двойная резистентность клеток свидетельствует о гомологичной рекомбинации, приводящей к разрушению гена. Было показано, что одна гомологичная рекомбинация происходит на 1000 случайных интеграции. В данной работе сам таргетируемый ген являлся селективным маркером. Для таргетирования неселективных генов был разработан метод позитивно-негативной селекции. Вектор для таргетинга содержит позитивный селективный маркер и негативный маркер - ген тимидинкиназы вируса герпеса HSV, которые расположены в разных местах вектора и каждый имеет свой промотор. Гомологичная рекомбинация с таргетированным геном приводит к встраиванию гена пео и элиминации гена tk, а негомологичная рекомбинация к встраиванию в хромосому обоих маркеров. Селекция гомологичных рекомбинатов проводится на среде, содержащей G418 и противовирусный препарат ганцикловир. Ганцикловир способен фосфорилироваться герпесной тимидинкиназой и при последующем встраивании в ДНК при репликации приводит к гибели клеток, экспрессирующих эту тимидинкиназу. Ген tkприсутствует в клетках при случайном встраивании вектора и отсутствует в клетках, претерпевших гомологичную рекомбинацию. Таким образом последние селектируются, будучи пео- и ганцикловиррезистентными, тогда как клетки, включившие вектор путем случайной интеграции, будут иеорезистентны, но ганцикловир чувствительны. Другим геном, обеспечивающим отрицательную селекцию, является фрагмент гена дифтерийного токсина, кодирующего его Б-цепь. Продукт этого гена токсичен для эукариотических клеток за счет ингибирования синтеза белка. Если ген ДТ-А встраивается в геном вместе с вектором и экспрессируется, то происходит гибель клетки.
Существует еще ряд подходов для выявления клонов ЭС клеток. Более подробную информацию об этих подходах можно найти в соответствующем обзоре. Одним из важных факторов, влияющих на частоту гомологичной рекомбинации, является степень гомологии между хромосомной и экзогенной ДНК. В основном все линии ЭС клеток, которые используются для таргетинга, выделены из линии мышей 129Sv. В связи с этим в работах по гомологичной рекомбинации вектора конструируют на базе генов, клонированных из банка генов этих линий мышей.
Использование ЭС клеток в сочетании с методом гомологичной рекомбинации открыло новые возможности для направленного мутагенеза. В основном, в большинстве работ получены мыши, у которых в результате мутации произошло выключение изучаемых генов. Анализ мышей, гомозиготных по Аш-аллелям, выявил несколько типов фенотипического проявления индуцированных мутаций:
1. Летальный эффект в эмбриогенезе или сразу после рождения.
2. Нарушение развития, функций, морфологии различных органов и гибель до наступления половой зрелости.
3. Нормальное развитие, жизнеспособность, плодовитость, отсутствие отклонений от нормы.
Например, в случае эмбриональных леталей происходит инактивация таких генов, как: nfl, Rb, N-туе, GAP, c-jun, c-myb. Гибель после рождения наблюдается при выключении следующих генов; c-abl, BDNF, trk, N-myc, TGF-3. Инактивация генов: CNTF, Dlx-2, FNAR-1, Ki-ras, c-mos, AT-2 приводит к нарушению развития, функций и морфологии разных органов. Нормальное развитие без изменения жизнеспособности, плодовитости наблюдали при инактивации таких генов, как: ароЕ, N-ras, MyoD, pim-1, c-fos. Очевидно, в этих случаях функции определенных генов могут выполняться другими родственными генами.
Стратегия генного таргетинга позволяет изучать функции отдельных генов как в развитии, так и во взрослом состоянии и моделировать на животных наследственные аномалии, встречающиеся у человека, с целью исследования их патогенеза и фенотипического проявления.
В настоящее время методы выделения ЭС клеток разработаны для разных видов животных: свиньи, овцы, коровы, норки, кролика и хомячка. В 1998 г. появилось два сообщения о выделении ЭС клеток человека из бластоцист и из первичных половых клеток зародыша. Несмотря на широкое распространение линий ЭС клеток и их интенсивное использование в экспериментах, остается еще достаточно много вопросов по вопросам их культивирования и дифференцировки. В настоящей работе представлены результаты экспериментов по морфологической характеристике начальных стадий эмбрионального развития, влиянию продуктов некоторых генов на пролиферацию и дифференцировку этих клеток в культуре, а также данные по таргетингу гена, кодирующего фактор роста фибробластов.
Характеристика эмбриональных стволовых клеток мыши
Отсутствие экспериментальных подтверждений бессмертности ЭС клеток и сложность их культивирования послужили основанием для проведения исследований по выявлению и поддержанию плюрипотентных свойств, отличающих эмбриональные клетки от соматических клеток культуры тканей. В настоящей работе были использованы две линии ЭС клеток мыши D3 и R1, обе линии выделены из бластоцист линии мышей 129/Sv. Одним из основных критериев плюрипотентности ЭС клеток принято считать форму колоний. Однако этот показатель во многом носит условный характер. По форме колоний иногда трудно отличить истинно эмбриональные клетки от клеток, спонтанно трансформированных в эмбриональную карциному. При фидерном способе культивирование формируются монослойные колонии округлой или удлиненной формы, с четко выраженными краями по периферии. Дифференцировка ЭС клеток приводит к морфологической гетерогенности и появлению колоний с неровными краями и тенденцией к вертикальному росту. Исследования ультраструктуры ЭС клеток показали, что клетки имеют относительно крупное ядро, содержащее преимущественно эухроматин, одно или два ядрышка, в которых доминируют гранулярные компоненты. В отличие от общепринятых представлений о том, что ЭС клетки лишены большинства органелл, в их цитоплазме были обнаружены многочисленные свободные рибосомы, шероховатый эндоплазматический ретикулум представлен единичными цистернами, что свидетельствует о низком уровне синтетической активности, комплекс Гольджи, около которого располагались 1-2 центриоли, митохондрии и многочисленные лизосомы.
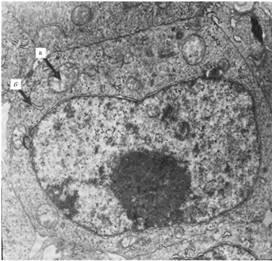
Ультраструктура ЭС клетки. Х5800. Видно крупное ядро клетки, а также эндоплазматический ретикулум и митохондрии
Аналогичная ультраструктура была выявлена в ранних мышиных зародышах на стадии бластоцисты. Клетки внутренней клеточной массы и ЭС клетки прошедшие, в культуре несколько пассажей, на ультраструктурном уровне имели большое сходство, что свидетельствует об общности их происхождения.
Были обнаружены небольшие межлинейные различия в пролиферативной активности и способности к дифференцировке, что, возможно, определялось генетической принадлежностью и возрастом ЭС клеток, т.е. количеством пассажей.
В популяции ЭС клеток преобладали клетки среднего размера. Они обладали адгезивными и пролиферативными свойствами, при контакте с фидером давали нормальные по внешнему виду монослойные колонии, а также были способны к дифференцировке в эмбриоидные тела. Более мелкие и более крупные, оставались не прикрепленными к субстрату и через некоторое время погибали или формировали колонии дифференцированных клеток. При этом морфологическая дифференцировка сопровождалась увеличением продолжительности стадии метафазы, а также частоты анеуплоидных и полиплоидных клеток и их гибели по механизму апоптоза.
Цитохимическое окрашивание ЭС клеток на выявление активности эндогенной щелочной фосфатазы позволяет объективно оценить степень дифференцированное этих клеток. Окрашиваются колонии, сформированные недифференцированными эмбриональными клетками. Клетки фидера не дают положительной реакции на выявление активности щелочной фосфатазы.
Разные линии мышиных ЭС клеток гетерогенны по морфологии, требовательны к составу питательных сред, сыворотки и качеству фидера. Вопрос о том, какие из ЭС клеток являются плюрипотентными, т.е. способными нормально развиваться в составе целого зародыша, является важнейшим при решении задач биологии развития и эмбриотехнологии.
Трансформация ЭС клеток геном
lif
Показано, что для поддержания ЭС клеток в плюрипотентном состоянии необходимо их культивирование на фидерном слое или в присутствии некоторых цитокинов, в частности LIF. В связи с этим были начаты исследования по изучению роли цитокина LIF в регуляции и поддержании плюрипотентных свойств ЭС клеток мыши invitro. Эти исследования необходимы для выявления роли цитокина в процессах регуляции межклеточных взаимодействий и генетической стабильности.
Для проведения трансформации геном lifиспользовали плазмиду pcDNA3, несущую кодирующую последовательность гена lifмыши, и ген пео, обеспечивающий устойчивость к антибиотику G418. Трансформацию клеток R1 проводили с помощью метода электропорации. Трансформированные геном lifклоны были способны развиваться в культуре, при переносе их на подложку из желатины, с последующей селекцией на выживаемость в среде с высоким содержанием G418. Массовая гибель после трансформации наблюдалась на 3-й сутки культивирования. У трансформированных клеток R1 обнаруживается низкий уровень пролиферативной активности. В селективной среде с G418 клетки формируют небольшие плоские колонии без признаков морфологической дифференцировки в течение 10 дней. При окрашивании на выявление активности эндогенной щелочной фосфатазы G418 резистентные клоны дают положительную реакцию. По морфологии колоний можно говорить о том, что сама процедура трансформации, по-видимому, не сопряжена с потерей плюрипотентности ЭС клеток.
Для детектирования уровня экспрессии гена lif, устойчивые к G418 клоны клеток линии R1 были проанализированы методом РТ-ПЦР, с использованием соответствующих праймеров. Было показано, что экспрессия гена обнаруживается в клонах с видимыми признаками морфологической дифференцировки. Возможно, что высокий уровень эндогенного LIF в трансфицированных клетках тормозит их пролиферативную активность и способствует переходу в дифференцированное состояние.
Необходимость применения фидерного слоя при культивировании ЭС клеток связано с продукцией LIF клетками первичных эмбриональных фибробластов, которые обеспечивают достаточный уровень концентрации этого фактора в непосредственной близости от ЭС клеток. Попытка стабилизировать плюрипотентные свойства ЭС клеток мыши линии R1 путем трансформации их геном подтверждает его участие в процессах раннего развития и дифференцировки.
Дифференцировка ЭС клеток
in
vitro
Одним из основных признаков ЭС клеток invitroявляется их способность дифференцироваться с образованием эмбриоидых тел различной сложности, которые представляют собой зачатки всех трех зародышевых листков. Дифференцировка ЭС клеток может происходить как спонтанным путем, так и под действием некоторых химических реагентов и ростовых факторов. Известно, что активин А и трансформирующий фактор роста бетта индуцирует дифференцировку клеток мезодермального происхождения соматических мышечных и кардиомиоцитов. Другая группа факторов - ретиноевая кислота, фактор роста фибробластов и эпидермальный фактор роста - направляет дифференцировку мезодермальных и эктодермальных клеток. Наконец, фактор роста нервов и фактор роста гепатоцитов участвуют в развитии всех трех эмбриональных листков, их воздействие приводит к появлению клеток печени и поджелудочной железы, мышечных, хрящевых, кроветворных и нейрональных клеток. Интересно отметить, что при разных концентрациях ретиноевой кислоты можно добиться преимущественной дифференцировки, либо в нейрональном, либо в миогенном направлении. Исследования генетического контроля миогенной и эпителиальной дифференцировки ЭС клеток показало, что последовательность экспрессии тканеспецифических генов такая же, как и в процессе нормального развития.
Существует ряд фактов, указывающих, что некоторые клеточные прото- и онкогены принимают определенное участие в пролиферации и дифференцировке клеток. Действие ТФА, стимулирующего дифференцировку клеток нейробластомы человека, одновременно повышает активность c-src. Микроинъекция мутантного онкогена c-Ha-rasв клетки феохромоцитомы крысы PC-12 вызывает их дифференцировку и появление нейроноподобных клеток. Однако в этих работах использовались злокачественные клетки, поэтому представляет интерес исследовать возможность индуцировать дифференцировку нормальных плюрипотентных стволовых клеток путем воздействия на них клеточных прото- и онкогенов.
Одной из задач наших исследований явилось проведение сравнительного анализа действия регуляторных генов вируса иммунодефицита человека типа 1 tatи nefна пролиферацию и дифференцировку ЭС клеток мыши invitro. Продукты генов tatи nefимеют противоположное влияние на клеточные гены. Ген tat, обладая онкогенным потенциалом, активирует транскрипцию некоторых клеточных генов и повышает пролиферацию клеток. Ген nef, подавляет экспрессию отдельных клеточных генов, и снижает рост клеток, вместе с тем кооперирует с геном tatпри образовании фокусов трансформации. Для проведения этих исследований была отработана методика получения эмбриоидных тел в линии R1, При культивировании клеток RI в суспензии на 2-3-и сутки они образовывали плотные шарообразные агрегаты, называемые простыми эмбриоидными телами. При дальнейшем культивировании эмбриоидные тела увеличиваются в своих размерах и усложняются в строении. При переносе эмбриоидных тел на подложку с желатиной, они прикрепляются к субстрату, происходит миграция клеток из эмбриоидного тела по поверхности и их дифференцировка в различные клеточные типы. В результате проведенных экспериментов показано, что эффективность образования эмбриоидных тел клетками линии R1 зависит от количества клеток, высеваемых на поверхность чашки без фидерного слоя, способных образовывать эмбриоидные тела и времени культивирования эмбриоидных тел до начала дифференцировки.
Известно, что бутират натрия вызывает ряд эффектов на клетках млекопитающих в культуре, включая снижение репликации ДНК, которая приводит к остановке деления клеток, изменению морфологии клеток, изменению уровня экспрессии некоторых генных продуктов. Показано, что бутират натрия индуцирует дифференцировку клеток тератокарциномы. Нами были проведены эксперименты по спонтанной дифференцировке, а также под влиянием бутирата в клетках R1. Обнаружено появление кластеров сокращающихся клеток, предположительно кардиомиоцитов, на 2-3-и сутки, после прикрепления к подложке эмбриоидных тел. Время развития сокращающихся клеток invitroколеблется от 2 до 10 сут. Частота пульсации этих клеток также менялась в зависимости от времени после посева эмбриоидных тел. В первые сутки частота не превышала 60 сокращений в минуту, затем она снижалась. Очевидно, способность ЭС клеток к дифференцировке invitroявляется уникальной моделью для исследования процессов клеточной дифференцировки в разных типах тканей. Особенно важно, что эта клеточная система обладает способностью к направленной дифференцировке при воздействии эндогенных и экзогенных факторов. В связи с этим мы проводили исследование по изучению влияния регуляторных генов вируса иммунодефицита человека типа 1 tat и nefна пролиферацию, морфологию и дифференцировку ЭС клеток мыши. Для проведения трансфекции ЭС клеток линии R1 использовали следующие рекомбинантные плазмиды; pTat, содержащую ген tatпод промотором цитомегаловируса и ген пео, устойчивость к неомицину, pNef, несущую ген nef под контролем промотора цитомегаловируса
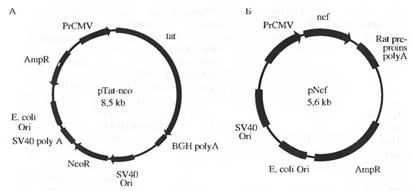
Схема плазмиды pTat. 3,1 кб Sall/PstI-фрагмент изолята НХВ-2 HIV-1, в котором делетировав ген nef, содержит 1-й транслируемый экзон гена tat, интрон и 2-й транслируемый экзон гена tat. Ген находится под управлением промотора цитомегаловируса в векторе pcDNA3. Плазмида несет также ген устойчивости к неомицину; схема плазмиды pNef. Плазмида содержит ген nefизолята HXD-3 HIV-1 под цитомегаловирусным промотором в векторе pBC12/CMv
Для выявления трансфектантов ЭС клеток в варианте с геном nefпроводили котрансформацию с плазмидой pSV2neo. Параллельно клетки R] трансфицировали плазмидой pSV2neoи использовали как контрольные. В результате проведения трансфекции и селекции эмбриональных стволовых клеток мыши получены 3 линии клеток; пеок- контроль. Изменения в морфологии наблюдали только для клеток линии nef. Как видно из рис. при изучении пролиферативной активности клеток всех трех выделенных линий обнаружено, что пролиферация клеток линии tatбыла выше, а полиферация клеток линии nefснижалась примерно в 1,5 раза по сравнению с контрольными клетками линии пеок.
Для изучения возможного влиния генов tatи nef па дифференцировку ЭС клеток проводили эксперименты, где определяли следующие показатели для трех линий клеток: 1) время, необходимое для формирования эмбриоидного тела; 2) количество сформированных эмбриоидных тел; 3) размер эмбриоидного тела; 4) появление клеток с различной тканеспецифичностью. В результате проведенных экспериментов было показано, что время необходимое для образования эмбриоидных тел, одинаково для клеток линии пеок, tat, nef. Клетки линии пеок и nefформируют примерно одинаковое количество эмбриоидных тел - от 3 до 6 - на 3-й сутки после посева, тогда как клетки линии tatформируют в основном только одно эмбриоидное тело. Значительных различий в размерах эмбриоидных тел для этих линий не обнаружено. На 5-й день после прикрепления эмбриоидных тел клеток линии пеок к подложке появились участки сокращающихся клеток-кардиомиоцитов, что было подтверждено результатами иммуноцитохимического анализа. Количество эмбриоидных тел с сокращающимися кардиомиоцитами увеличивалось в клетках линии пеок в течение 5 дней. В клетках линии не кардиомиоциты появились только на 10-й день после прикрепления эмбриоидных тел к подложке, их количество заметно не увеличивалось в период проведения эксперимента. В эмбриоидных телах, сформированных клетками линии tatне было обнаружено появление кардиомиоцитов в течение 10 дней наблюдения. Результаты представленных экспериментов позволяют сказать, что под влиянием гена tatнаблюдается тенденция повышения пролиферативной активности ЭС клеток мыши и ее подавление под влиянием гена nef, что согласуется с данными, полученными на клетках крысы двух линий: псевдонормальных клетках Rat-2 и клетках феохромоцитомы PC-12. Полученные данные также позволяют предположить, что регуляторные гены ВИЧ-1 tatи nefмогут оказывать влияние как на ранние стадии дифференцировки ЭС клеток, так и на процесс дифференцировки этих клеток в кардиомиоциты.
Разработка методов направленной тканеспецифической дифференцировки, помимо теоретического значения для изучения молекулярных механизмов клеточной дифференцировки, имеет и прикладное значение, такие клетки в перспективе могут быть использованы в клеточной терапии и трансплантации у человека.
Для изучения путей дифференцировки ЭС клеток в разные типы клеток invivoиспользуются подходы по пересадке таких клеток в различные органы взрослых животных с последующей регистрацией судьбы этих клеток. Для объективного обнаружения пересаженных клеток необходимо иметь "маркированные" линии ЭС клеток. Нами были получены линии ЭСК, трансфицированные геном "зеленого белка". Такие клетки легко детектируются на срезах органов и тканей с помощью флуоресцентной микроскопии. На рис. ПО показаны ЭС клетки линии R1, трансфицированные плазмидой, содержащей ген "зеленого белка".
Проведенные нами совместно с Институтом биологии гена РАН и Институтом биологии развития РАН по пересадке "зеленых клеток" в различные отделы мозга крысы, показали, что такие клетки могут сохранять жизнеспособность в чужеродном организме не менее одной недели.
Гомологичная рекомбинация в ЭС клетках мыши
В нашем отделе была проведена работа по нокаутированию гена PDGF-Aв ЭС клетках мыши и получению на основе этих клеток химерной мыши.
PDGF-A- белок, фактор роста тромбоцитов. Существует три типа димеров PDGF: А А, АВ и ВВ, состоящих из двух полипептидных цепей - PDGF-A и PDGF-B, которые кодируются двумя различными генами. Эти гены являются гомологичными и кодируют длинную и короткую формы PDGF, получающиеся путем альтернативного сплайсинга. В свою очередь существует 3 вида рецепторов для PDGF, кк и кк, также состоящих из полипептидов -PDGFR и PDGFR, кодируемых разными генами. Митогенная активность PDGF реализуется при его взаимодействии с рецептором, при этом PDGFR-a может связываться как с А-, так и с В-цепью PDGF, а PDGFR - только с В-цепью. Основным местом синтеза PDGF являют ся тромбоциты, хотя в незначительных количествах они синтезируются в эндотелии, моноцитах и макрофагах, гладкой мускулатуре, фибробластах, цитотрофобластах плаценты, нейронах и в глии. В большинстве клеточных типов экспрессия PDGF происходит при функциональной или митогенной активации, хотя в мегакариоцитах и нейронах она, по-видимому, конститутивна. По некоторым данным, помимо митогенной активности, PDGF принимает участие в процессах дифференцировки, хемотаксиса, в регуляции синтеза компонентов межклеточного матрикса. Обнаружена экспрессия PDGF и рецепторов к этим ростовым факторам как на самых ранних, доимплантационных, стадиях эмбрионального развития, так и при дифференцировке мезенхимы и нервной ткани PDGF продуцируют также многие опухоли, такие, например, как глиобластомы, нейробластомы, меланомы и остеосаркомы.
Вектор замещения для таргетинга конструировали на основе гена PDGF-A, проклонированного из библиотеки генов линии мышей 129Sv.
На рис. представлены этапы конструирования и окончательная структура соответствующего вектора. Этот вектор включал в себя фрагмент, содержащий экзоны 3 и 4. В экзон 3 был вставлен ген пео, обеспечивающий устойчивость к антибиотику неомицину. В качестве промотора гена пео использовали промотор гена фосфоглицерокиназы. Для проведения негативной селекции против случайного встраивания в геном использовали кассету с геном субъединицы А дифтерийного токсина. В работе использовали линию ЭС клеток w9.5, выделенную из линии мышей 129SvJ. Для получения химерных мышей ЭС клетки инъецировали в полость 3,5-дневных бластоцист мышей линии C57BI/6. Далее бластоцисты переносили в рог матки мышей линии BALB/c в сроки псевдобеременности 2,5-3,5 дня. Трансфекцию ЭС клеток проводили методом электропробоя. После электропорации ЭС клеток и последующей селекции было получено около 400 G418 устойчивых клонов ЭС клеток мыши, из которых 130 были заморожены и использованы для выделения ДНК. Среди первых 5 клонов, проанализированных с помощью блотгибридизации, был обнаружен один клон с таргетированным геном PDGF-А.
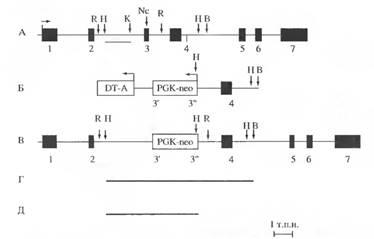
Схема строения гена PDGF-A, вектора, использованного для таргетинга, и мутантного гена PDGF-Aсо встроенной в его 3-й экзон PGK-иео-кассетой. Г - Handlll-фраг-мент гена PDGF-Aдикого типа, Д - Hindlll-фрагмент гена. DT-A - кассета с геном субъединицы А дифтерийного токсина. PGK-neo-кассета с геном неомицинфосфотрансферазы под промотором гена фосфоглицерокиназы. Зонд отмечен линией под А. Заштрихованными и нумерованными прямоугольниками изображены экзоны гена PDGF-A, а линиями междуними - нитроны. Стрелками указаны направления транскрипции генов. Сайты рестрикции: З - Hindlll, А' - Kpnl, В - BamHI, Ж - EcoRl, Mr - Ncol
Следующим этапом работы было получение химерных мышей путем инъекции клеток клона 1. В бластоцисты мыши. В результате пересадки 13 бластоцист родилась одна химерная самка со смешанной пигментированной шерстью черного цвета и цвета агути, что свидетельствует о способности таргетированных ЭС клеток принимать участие в развитии волосяных фолликулов в эмбриогенезе.
У гомозиготных мышей с нокаутированным геном PDGF-Aнаблюдались патологии во многих тканях мезенхимного происхождения. При развитии легких эта мутация приводила к эмфиземе и невозможности формирования альвеол. Как и ожидалось, гомозиготные по мутантному гену мыши погибали либо на эмбриональных стадиях, либо фазу после рождения. Эта работа уже дала первую важную информацию о роли PDGF-А. Однако поскольку имеются данные о различном фенотипическом проявлении гомозиготных "нокаутированных" генов в различных линиях мышей, одним из возможных вариантом продолжения этой работы является получение линии мышей, гомозиготной по "нокаутированному" гену PDGF-А с целью более детального анализа функций этого гена.
Заключение
В последнее время все большее внимание исследователей направлено на возможность клонирования млекопитающих путем переноса ядер в энуклеированные ооциты. Преимущество этого метода заключается в том, что все клетки в потомстве, включая зародышевые, представлены генотипом ЭС клеток. ЭС клетки сельскохозяйственных животных могут служить источником тотипотентных ядер, которые могут быть использованы в качестве доноров для пересадки ядер с целью получения генетически идентичных трансгенных животных причем практически в неограниченном количестве. Широкие перспективы открываются в плане возможного использования эмбриональных стволовых и стволовых клеток для клеточной терапии различных тяжелых заболеваний человека, таких как, например, диабет, онкологические и нейродегенеративные заболевания. Необходимым для достижения этой цели условием является получение invitroкак можно более "гомогенных" клеточных культур. В связи с этим чрезвычайно важным является поиск факторов как белковой так и небелковой природы, способных регулировать процессы дифференцировки этих клеток в определенном направлении. Таким образом, комплексные исследования соматических клеток млекопитающих и человека, включая эмбриональные стволовые клетки, открывают широкие возможности как для продолжения исследований фундаментальных молекулярно-генетических процессов, лежащих в основе жизнедеятельности этих клеток, так и для практической медицины XXI в.
|