МИНИСТЕРСТВО НАУКИ И ОБРАЗОВАНИЯ УКРАИНЫ
УКРАИНСКИЙ ГОСУДАРСТВЕННЫЙ ХИМИКО -ТЕХНОЛОГИЧЕСКИЙ УНИВЕРСИТЕТ
КАФЕДРА БИОТЕХНОЛОГИИ
И БЕЗОПАСНОСТИ ЖИЗНЕДЕЯТЕЛЬНОСТИ
КУРСОВАЯ РАБОТА
ТЕМА: АМИЛОЛИТИЧЕСКИЕ ФЕРМЕНТЫ
Выполнил
………………………………………………………ст.гр.5-Ф-69
Клименко Е.О.
Проверил
……………………………………………профессор Лихолат Е.А.
Днепропетровск
2009 г.
СОДЕРЖАНИЕ
1. Введение
……………………………………………………………………4
2.
Общая часть
………………………………………………………………8
2.1. Основные понятия энзимологии ………………………………………...8
2.1.1. Понятие о ферментах как о биологических катализаторах………8
2.1.2. Классификация и номенклатура ферментов……………………....8
2.1.3. Химическая природа и строение ферментов. …………………….9
2.1.4. Механизм действия ферментов и их специфичность…………….9
2.1.5. Кинетика ферментативных реакций. ……………………………..12
2.1.6. Ингибирование ферментов………………………………………...13
2.1.7.Активация ферментов………………………………………………14
2.1.8. Единицы активности ферментов………………………………….15
2.1.9. Внутриклеточная регуляция ферментативной активности……...15
2.1.10.Методы определения активности ферментов……………………16
2.1.11. Получение ферментных препаратов…………………………….19
3. Амилолитические препараты
..................................................................21
3.1. Источники получения амилаз………………………………………….21
3.2. Механизм действия и свойства амилаз………………………………...21
3.3. Компьютерное моделирование структуры амилолитических ферментов
……………………………………………………………….26
4. Получение микробных ферментных препаратов
……………………32
4.1. Основные сведенья…………………………………………………….32
4.2. Микроорганизмы – продуценты ферментов…………………………..32
4.2.1. Микроскопические грибы…………………………………………32
4.2.2. Дрожжеподобные организмы……………………………………..34
4.2.3. Бактерии……………………………………………………………34
4.3. Производственные способы культивирования микроорганизмов – продуцентов ферментов…………………………34
4.3.1. Поверхностное культивирование микроскопических
грибов……………………………………………………………………..35
4.3.2. Глубинное культивирование микроорганизмов…………………40
4.4. концентрирование ферментных растворов……………………………42
4.5. Оборудование для получения амилолитических ферментов…………43
4.5.1Назначение…………………………………………………………..43
4.5.2.Области применения………………………………………………..43
4.5.3.Устройство………………………………………………………….44
4.5.3.1Модуль для приготовления питательной среды (МПС)………..44
4.5.3.2.Модуль стерилизационный (МС)……………………………….45
4.5.3.3.Модуль ферментационный (МФ)………………………………45
4.5.3.4.Модуль интерактивного управления (МУ) ……………………45
4.5.3.5.Выполняемые функции…………………………………………46
4.5.3.6.Примеры использования ОКА-01………………………………47
Литература
………………………………………………………………..49
1. Введение
Изучение ферментов представляет особый интерес, так как эта область знания находится на стыке биологических и физических наук. С одной стороны, ферменты имеют исключительное значение в биологии. Жизнь зависит от сложной совокупности химических реакций, осуществляемых специфическими ферментами, и любое изменение деятельности ферментов может повлечь за собой серьезные последствия для живого организма. С другой стороны, ферменты как катализаторы все больше и больше привлекают внимание физико-химиков. Изучение механизма действия ферментов составляет само по себе одну из самых увлекательных областей современного научного исследования.
В настоящее время наука о ферментах - энзимология превратилась в большую бурно развивающуюся отрасль знания с многочисленными ответвлениями, тесно связанную со многими науками, особенно с биохимией, физической химией, бактериологией, микро-биологией, генетикой, ботаникой, сельским хозяйством, фармакологией, токсикологией, патологией, физиологией, медициной и химической технологией. Кроме того, она имеет важное практическое применение в столь различных областях, как, например, пивоварение и другие бродильные производства, борьба с вредителями в сельском хозяйстве и химическая война.
Многие научные работники в различных частях света посвятили себя изучению различных энзимологических проблем; учреждены специальные институты по изучению ферментов, существует целый ряд энзимологических журналов, и по отдельным вопросам энзимологии к настоящему времени имеется обширная литература.
Иногда трудно себе представить, что энзимология возникла сравнительно недавно; зарождение этой области науки можно отнести к первой половине XIX в., но особенно бурное развитие она претерпела в течение последних 35 лет. Хотя явления брожения и переваривания были известны с незапамятных времен, первое ясное представление о ферменте, по-видимому, было дано только в 1833 г. Пайоном и Персо, когда они обнаружили в осадке, образующемся при добавлении спирта к солодовому экстракту, термолабильное вещество, обладающее способностью превращать крахмал в сахар. Это вещество, которое мы теперь называем амилазой, было названо ими диастазой (от греческого слова  — разделять) благодаря его способности отделять растворимый декстрин от нерастворимой оболочки крахмальных зерен. Название «диастаза» впоследствии было распространено на все ферменты как видовое наименование. — разделять) благодаря его способности отделять растворимый декстрин от нерастворимой оболочки крахмальных зерен. Название «диастаза» впоследствии было распространено на все ферменты как видовое наименование.
В 1898 г. Дюкло предложил последние три буквы этого видового названия (суффикс «аза»)прибавлять к корню названия того вещества, на которое данный фермент действует. Этот принцип был положен в основу применяемой ныне номенклатуры ферментов; однако несколько названий
пищеварительных ферментов, оканчивающиеся на «ин», удержались оканчи-
вающиеся на «ин», удержались до настоящего времени. Поскольку число известных ферментов продолжает увеличиваться, в названии фермента стали указывать не только природу вещества, на которое он действует, но также и характер катализируемой им реакции (например, лактатдегидрогеназа).
Сразу же после открытия ферментов многие исследователи обратили внимание на сходство между их действием и действием дрожжей при бро-жении. Поэтому к агентам, вызывающим брожение, стали применять термин «фермент». Во второй половине XIX в. разгорелся большой спор между Либихом, придерживавшимся того взгляда, что брожение и сходные процессы обусловлены действием химических веществ, и Пастером, утверждавшим, что брожение неотделимо от жизнедеятельности клеток. Отсюда возникло разделение ферментов на «неорганизованные» и «организованные», т.е., как сказали бы мы теперь, на экстрагируемые ферменты и ферменты, функционирующие в живой клетке. Для того чтобы избежать применения этих неудобных наименований, В. Кюне в 1878 г. ввел в употребление термин «энзим». Поскольку среди ученых имели хождение неправильные представления о причинах, побудивших Кюне предложить этот термин, мы цитируем высказывание самого Кюне:
«Таким образом, мы не намереваемся выдвигать какую-либо особую гипотезу, а только констатируем, что в дрожжах имеется что-то, что обладает той или иной ферментативной активностью. Это название, однако, применимо не только к инвертину дрожжей. Оно должно подчеркнуть, что более сложные организмы, из которых можно получить такие ферменты, как пенсин, трипсин и т. п., не столь резко отличаются от одноклеточных организмов, как это иногда принято думать».
Спор Пастера и Либиха был разрешен, после того как Бухнеру удалось получить систему ферментов, осуществляющих брожение, из дрожжевого экстракта, не содержащего дрожжевых клеток. Однако название «фермент» сохранилось в Германии до настоящего времени.
Достигнутые к концу XIX в. замечательные успехи в области стереохимии органических веществ, представляющих биологический интерес, сделали возможным изучение пределов действия ферментов, или их так называемой «специфичности». Эмилю Фишеру мы обязаны развитием представления о специфичности ферментов и о близком стерическом соответствии между ферментом и субстратом. На основании своих наблюдений над субстратами известной структуры Фишер выдвинул свое знаменитое положение о том, что фермент подходит к субстрату так, как ключ к замку. Специфичность составляет в настоящее время очень важный раздел учения о ферментах. Вследствие существования близкого соответствия между ферментом и субстратом фермент может действовать только на очень ограниченный ряд субстратов, и это является причиной существования большого числа различных ферментов. Изучение специфичности ферментов, разумеется, невозможно без выделения индивидуальных ферментов в чистом виде.
Серьезных работ по получению ферментов в чистом виде до 1920 г. не было. Большинство ранних работ по очистке ферментов было проведено Вильштеттером и его сотрудниками между 1922 и 1928 гг.. Несколько работ было проведено в течение этого периода и другими исследователями (в качестве примера можно указать опыты Диксона и Кодама по очистке ксантиноксидазы), но ни водном из этих случаев не была достигнута полная чистота. Следующим важным достижением было получение ферментов в кристаллическом виде. Первым ферментом, полученным в кристаллическом виде, оказалась уреаза, кристаллы которой получил Самнер в 1926 г.. Однако эти первые кристаллы были далеко не чистыми. За этой работой вскоре последовала серия классических работ Нортропа и его сотрудников по выделению в кристаллическом виде протеолитических ферментов. И хотя еще 20 лет назад число полученных в чистом виде ферментов было очень мало, в наше время число ферментов, полученных в чистом и кристаллическом виде, уже достигает 100, а очищенных до той или иной степени превышает 500.
Первая работа по очистке ферментов была встречена резкой критикой. Указывалось, что «нефизиологично» выделять ферменты из клетки и что ценность представляют только те методы, которые позволяют изучать ферменты в неповрежденных клетках. Позднее, когда были получены первые
белковые кристаллы, обладающие высокой ферментативной активностью, возникли сомнения относительно возможности кристаллизации самих ферментов. Предполагалось, что ферменты лишь адсорбированы на кристаллах инертного белка. Нортроп, однако, привел убедительные доказательства того, что полученные им белковые кристаллы сами являются ферментами. С тех пор были выделены в чистом виде многие ферменты, причем все они также оказались белками. Таким образом, представление о ферменте с течением времени менялось. Сначала этим словом обозначали неопределенные влияния или свойства, присущие некоторым препаратам, потом — определенные химические вещества и, наконец, специфические белки.
Главный интерес в ранний период развития энзимологии сосредоточивался на изучении ферментов пищеварения и брожения; только значительно позднее была установлена важность внутриклеточных ферментов. Фактически серьезных работ по очистке внутриклеточных ферментов до 1937 г. не было, и это несмотря на то, что сравнительно мало ферментов встречается в естественных выделениях живых клеток. Однако с 1937 г. положение полностью изменилось, и то громадное увеличение числа известных ферментов, которое мы имеем теперь, обусловлено главным образом открытием новых внутриклеточных ферментов. Это расширение наших знаний о ферментах живого вещества привело в свою очередь к значительно более глубокому пониманию механизма многих основных
жизненных процессов, особенно лежащих в основе жизни обменных процессов, связанных с запасанием и использованием энергии. Наши знания о таких процессах, как фотосинтез, дыхание, биологическое окисление, брожение, синтез многих необходимых для роста органических веществ, выполнение механической или осмотической работы, значительно пополнились благодаря выделению и изучению соответствующих ферментов.
Возможность получать ферменты в чистом виде позволила в последние годы предпринять их количественное изучение с помощью физико-химических методов. Учение о кинетике ферментов, в основе которого лежат классические работы Михаэлиса, начатые в 1913 г., достигло к настоящему времени значительного развития и продолжает развиваться дальше преимущественно в направлении изучения механизма ферментативного катализа. Использование других методов, в том числе изотопных и оптических, а также химическое изучение активных групп ферментов тоже проливает свет на способ их действия.
Очистка ферментов способствовала также более глубокому пониманию их высокой специфичности, являющейся одним из наиболее характерных и важных свойств ферментов. Это свойство резко отличает ферменты от других катализаторов. Благодаря специфичности возможно существование полиферментных систем, осуществляющих цепи реакций, из которых состоят
различные обменные процессы.
Значительные успехи были достигнуты в области изучения свойств и функций ферментов. Однако в этой области предстоит еще очень большая работа: много важных ферментов еще только предстоит открыть; другие, на существование которых мы уже имеем указания, в настоящее время ожидают изучения.
Мы считаем, что в настоящее время энзимология достигла той стадии, когда в ближайшем будущем можно надеяться пожать обильные плоды всего громадного числа исследований, которые были проведены в последние годы. Особенно большие успехи достигнуты при изучении одной из самых увлекательных проблем энзимологии — механизма действия ферментов.
Наконец, никогда не следует забывать, что ферменты имеют столь большое значение вследствие того, что сама жизнь тесно связана с ферментным катализом. По необходимости развитие энзимологии происходило главным образом в направлении изучения изолированных ферментов. Только в последние годы появились возможности для развития биологической стороны предмета, а именно для изучения ферментов и ферментных систем непосредственно в живой клетке.
2. ОБЩАЯ ЧАСТЬ
2.1. ОСНОВНЫЕ ПОНЯТИЯ ЭНЗИМОЛОГИИ
2.1.1. Понятие о ферментах как о биологических катализаторах.
Ферменты
(лат. Fermentum – брожение, бродильное начало) – это специфические белки, способные во много раз ускорять биохимические реакции, протекающие в живых организмах, не входя при этом в состав конечных продуктов реакции. Т.е. ферменты (или другое их название – энзимы) являются биологическими катализаторами биохимических реакций.
Все биохимические реакции, протекающие в клетках микроорганизмов, в растительных и животных организмах, катализируются соответствующими ферментами. Переваривание, всасывание и усвоение пищевых веществ, синтез и распад белков, нуклеиновых кислот, жиров, углеводов и других соединений в тканях и клетках любого организма представляет собой совокупность ферментативных реакций. Любое функциональное проявление живого организма (дыхание, размножение и др.) обеспечивается действием соответствующих ферментных систем.
Ферменты обладают определенными свойствами, отличающих их от неорганических катализаторов: высокой субстратной специфичностью, высокой скоростью проведения ферментативной реакции, способностью ускорять реакции в физиологических условиях, т.е в условиях, характерных для жизнедеятельности клеток (соответствующая температура, реакция среды и др.)
2.1.2. Классификация и номенклатура ферментов.
В настоящее время число различных известных реакций, катализируемых ферментами, составляет более 2 тыс. и число их непрерывно возрастает. Для того, чтобы ориентироваться в этом множестве биохимических превращений, нужна некоторая систематика. Она создана Международным союзом по биохимии (InternationalUnionofBiochemistry, IUB) и рекомендована к повсеместному использованию. Согласно этой классификации все ферменты подразделяют на 6 классов в зависимости от типа катализируемых превращений:
1.Оксидоредуктазы
. К этому классу ферментов относятся практически все ферменты, катализирующие окислительно-восстановительные реакции.
2.Трансферазы
. К этому классу относятся ферменты, катализирующие различные превращения, представляющие собой перенос радикала от молекулы-донора к молекуле-акцептору и не являющиеся процессом гидролиза. В общем виде эти превращения можно записать так:
XY + Z X + YZ (X, Z X + YZ (X, Z H2
O или OH-
) H2
O или OH-
)
3. Гидролазы
– ферменты, катализирующие разнообразные реакции гидролиза. Подклассы в этом классе выделяют в соответствии с типом гидролизуемых связей.
4. Лиазы
. Ферменты этого класса катализируют в одном направлении гидролитическое расщепление субстрата с образованием кратной связи или, реже, цикла, а в другом направлении – присоединение по кратной связи.
5. Изомеразы
– это ферменты, катализирующие различные процессы изомеризации.
6.Лигазы (синтетазы)
. Этот класс составляют ферменты, катализирующие реакции конденсации или присоединения, сопряженные с гидролизом одной из пирофосфатных связей в молекуле АТФ или ГТФ.
2.1.3. Химическая природа и строение ферментов.
Все ферменты – белки, имеющие, как и другие белки, первичную, вторичную, третичную и часто четвертичную структуру.
Ферменты могут быть как простыми, так и сложными белками. Молекулы ферментов, представляющие собой простые белки, целиком построены из полипептидных цепей и при гидролизе распадаются только на аминокислоты. Примером ферментов такого рода являются гидролитические ферменты пепсин, трипсин, уреаза, лизоцим и др. Большинство природных ферментов относится к сложным белкам, содержащих в составе своей молекулы помимо полипептидных цепей небелковый компонент. Как белковая часть такого сложного фермента (апофермента), так и небелковый компонент его молекулы (кофермент) в отдельности лишены ферментативной активности. Только соединившись вместе и образовав так называемый холофермент (полный фермент), они приобретают свойства, характерные для биокатализаторов. В роли коферментов могут выступать различные соединения. Так, ряд коферментов является производными витаминов. По химическому строению многие коферменты представляют собой нуклеотиды, некоторые – органические производные фосфорной кислоты или пептиды и их производные (например, фолиевая кислота, глутатион, КоА). Таким образом, между ферментами, нуклеотидами и витаминами существует определенная функциональная связь.
2.1.4. Механизм действия ферментов и их специфичность.
Для объяснения механизма действия ферментов было предложено много теорий. Все они исходят из экспериментально установленного факта, что биокатализаторы, так же как и неорганические катализаторы, не могут сделать возможным протекание процессов или реакций, которые с точки зрения термодинамики невозможны. Все теории катализа допускают, что катализаторы тем или иным путем снижают энергию активации катализируемой реакции и таким образом резко увеличивают скорость химического превращения. Катализаторы лишь сокращают время, необходимое для достижения химической реакцией состояния подвижного
равновесия. Состояние подвижного равновесия обратимой химической реакции в присутствии катализатора становится стационарным. Следовательно, при наличии катализатора в равной мере возрастает скорость как прямой, так и обратной реакции.
Начало современным представлениям о механизме действия ферментов было положено работой Михаэлиса и Ментен, опубликованной в 1913 г. В этой работе впервые высказывалась идея о том, что основой действия ферментов является образование фермент-субстратного комплекса, который затем распадается с образованием продуктов ферментативной реакции и освобождением фермента в исходном состоянии.
В процессе образования и превращениях фермент-субстратных комплексов принято различать несколько стадий:
1. Присоединение молекулы субстрата к ферменту.
2. Последовательное превращение первичного фермент-субстратного комплекса в один или несколько активированных комплексов.
3. Отделение конечных продуктов реакции от фермента.
Специфичность, которую проявляют ферменты в отношении субстрата как при образовании фермент-субстратного комплекса, так и при каталитическом химическом превращении, обусловлена существованием в молекуле фермента специфического участка, называемого активным центром. Та часть активного центра, которая принимает непосредственное участие в каталитическом действии, получила название каталитического участка, а место, где осуществляется связывание фермента с субстратом, - контактного участка, или контактной площадки.
К 80-м гг. 20 в. был идентифицирован аминокислотный состав активных центров многих ферментов. В активный центр фермента могут входить аминокислотные остатки, сравнительно далеко отстоящие друг от друга в полипептидной цепи и сблизившиеся в результате ее ”перекручивания” при образовании вторичной и третичной структур ферментного белка. Так, в молекуле химотрипсина (протеолитического фермента животного происхождения) в состав активного центра входят остаток гистидина, находящийся в полипептидной цепи в положении 57, остаток серина в положении 195 и остаток аспарагиновой кислоты в положении 102.
Расположение аминокислотных остатков, формирующих контактный участок в молекуле фермента, способствует такому пространственному размещению субстрата, которое наиболее благоприятно для его атаки каталитическими (функциональными) группами активного центра. Например, реакция А + В  АВ, катализируемая соединениями, имеющие активные группы С, D и Е, в обычных условиях должна была бы происходить только в том случае, если бы одновременно столкнулись все пять реагентов. Вероятность такого столкновения ничтожно мала, поэтому скорость образования продукта реакции АВ в этом случае практически равна нулю. В том случае, когда эту реакцию катализирует фермент, требуется АВ, катализируемая соединениями, имеющие активные группы С, D и Е, в обычных условиях должна была бы происходить только в том случае, если бы одновременно столкнулись все пять реагентов. Вероятность такого столкновения ничтожно мала, поэтому скорость образования продукта реакции АВ в этом случае практически равна нулю. В том случае, когда эту реакцию катализирует фермент, требуется
только его взаимодействие с веществами А и В, поскольку группы С, D и Е уже включены в состав молекулы фермента. В результате эффекта близости реакционных групп в молекуле скорость катализируемой ферментом реакции увеличивается на несколько порядков. Кроме того, когда субстрат присоединяется к активному центру фермента, пространственная конфигурация молекулы субстрата изменяется и определенные химические связи в ней ослабевают.
Каталитическая активность фермента определяется структурой его молекулы в целом. Другие аминокислотные остатки, помимо входящих в активный центр, необходимы для формирования и стабилизации определенной конформации активного центра. Они также образуют участки, с которыми специфически взаимодействуют различные вещества, регулирующие активность фермента. В молекуле фермента имеются такие участки, за счет которых он может взаимодействовать с другими ферментами (при образовании мультиферментных комплексов) или белками мембран (мембранные ферменты).
Важнейшим свойством ферментов является их субстратная специфичность, т.е. способность не только ускорять реакции, но и избирательно катализировать лишь определенный путь превращения конкретного вещества – субстрата. По признаку специфичности действия все ферменты можно разделить на две группы:
1. Ферменты, обладающие абсолютной специфичностью.
2. Ферменты, обладающие относительной специфичностью.
Абсолютная специфичность проявляется тогда, когда фермент действует лишь на одно единственное вещество и катализирует только определенное превращение этого вещества. К абсолютной специфичности относят также стереохимическую (оптическую) специфичность. В живой природе встречаются лишь определенные пространственные формы соединений: аминокислоты в белках – только L-ряда, сахара – преимущественно D-ряда. Ферменты всегда действуют только на один из двух оптических изомеров вещества (L- или D-изомер). Специфичность ферментов может быть направлена и на другие типы стереоизомеров. Так, ферменты, действующие на транс-изомер, обычно неактивны в отношении цис-изомера того же вещества.
Наряду с ферментами, которым свойственна абсолютная специфичность, имеется большая группа ферментов, обладающих относительной или групповой специфичностью. Например, пепсин (протеолитический фермент животного происхождения) катализирует расщепление белковых молекул, которые могут существенно различаться друг от друга как по аминокислотному составу, так и по физико-химическим
свойствам. Объясняется это тем, что местом действия пепсина является пептидная связь – CO – NH – .
2.1.5. Кинетика ферментативных реакций.
Скорость ферментативной реакции зависит прежде всего от природы фермента, который может обладать низкой или высокой ферментативной активностью, а также от концентрации субстрата, величины рН среды, температуры, присутствия ингибиторов или активаторов и др.
Установлено, что при прочих равных условиях и при наличии избытка субстрата начальная скорость ферментативной реакции пропорциональна концентрации фермента.
Зависимость между концентрацией субстрата и скоростью ферментативной реакции графически можно выразить кривой, показанной на
рисунке 2.1.

Рис. 2.1. График, выражающий зависимость между скоростью ферментативной реакции (V) и концентрацией субстрата (S) при постоянной концентрации фермента: отрезок абсциссы Км
соответствует константе Михаэлиса (концентрация субстрата, при которой скорость данной ферментативной реакции составляет половину от максимальной – Vмакс
).
Скорость реакции, соответствующая восходящей части кривой, пропорциональна концентрации субстрата (S); в верхней части кривой скорость реакции приближается к максимальному значению (Vмакс
) и почти не зависит от концентрации субстрата.
Исходя из представлений об образовании фермент-субстратных
комплексов в ходе ферментативной реакции Михаэлис и Ментен рассчитали весьма важную константу, позже названную константой Михаэлиса (Км
), которая в числовом выражении равна той концентрации субстрата (моль/л), при которой скорость реакции составляет половину максимальной (т.е. когда в любой момент в комплексе с субстратом находится и половина молекул фермента). Таким образом, Км
является мерой сродства между ферментом и субстратом, причем низкие величины Км
свидетельствуют о высокой степени такого сродства.
Существует еще одна величина, характеризующая ферментативную реакцию, которая получила название ”число оборотов фермента”. Величина это показывает, сколько молекул субстрата превращается в продукты реакции за единицу времени в расчете на одну молекулу фермента.
Ферментативные реакции, подобно обычным химическим реакциям, ускоряются при повышении температуры. Установлено, что повышение температуры на каждые 10о
С увеличивает скорость ферментативной реакции в 1,5 – 2 раза (такая закономерность для большинства ферментов проявляется в интервале от 10 до 30о
С). Показано, что возрастание температуры на 1о
С повышает активность фермента на 4 – 10 %. Однако при температуре 50о
С и выше происходит постепенное снижение активности фермента. Дальнейшее нагревание (до 60 – 70о
С) приводит к постепенной денатурации и инактивации фермента. Степень инактивации зависит также от длительности температурного воздействия.
Ферменты весьма чувствительны к изменениям величины рН среды. Каждый фермент имеет оптимум рН, при котором он наиболее активен. Для большинства ферментов оптимальная величина рН близка к нейтральной ( рН=7,0). Таким образом, максимальная активность ферментов проявляется при физиологических значениях рН, а в кислой и щелочной средах их активность снижается. Из этого правила имеются, впрочем, исключения. Например, пепсин, содержащийся в желудочном соке активен лишь в очень кислой среде, а трипсин (протеолитический фермент животного происхождения, содержится в соке поджелудочной железы) при этих значениях рН полностью ингибируется. Оптимум действия трипсина лежит в пределах рН=8, т.е. в щелочной среде.
Кроме температуры, величины рН, концентрации субстрата большое влияние на активность ферментов оказывает присутствие ряда химических соединений. Одни из них повышают активность ферментов, другие, напротив, подавляют ее. Иногда, впрочем, соединения, являющиеся ингибиторами для одних ферментов, могут быть активаторами для других (чаще всего это относится к ионам металлов).
2.1.6. Ингибирование ферментов
Процесс ингибирования ферментов может быть обратимым и ингибирование ферментативных реакций. Конкурентное ингибирование возникает тогда, когда в среде одновременно присутствуют субстрат исходный по структуре с ним ингибитор (структурный аналог субстрата). В этом случае субстрат и ингибитор конкурируют за активный центр фермента.
Тормозящее действие конкурентного ингибитора на фермент зависит от концентрации субстрата. Если в окружении фермента находится много молекул субстрата и мало молекул ингибитора, то во взаимодействие с
активным центром фермента значительно чаще вступает субстрат. В этом случае ферментативная реакция не прекращается, а лишь замедляется. Если в инкубационной среде создать избыток субстрата, то конкурентное ингибирование прекращается.
В случае неконкурентного торможения (ингибирования) ингибитор не имеет структурного сходства с субстратом. Поэтому повышение концентрации субстрата не может привести к вытеснению ингибитора из комплекса фермент-ингибитор, как это происходит при конкурентном торможении.
2.1.7.Активация ферментов
Наряду с ингибиторами ферментов существуют также и активаторы ферментов. Весьма часто роль активаторов выполняют ионы металлов. Нередко действие иона металла в качестве активатора заключается в связывании субстрата с ферментом. В ряде случаев действие ионов металлов абсолютно специфично, т.е. для активации данного фермента требуется наличие определенного иона. Однако имеются ферменты, способные в одинаковой степени активироваться одними и теми же ионами металлов. Ион металла может являться постоянным компонентом активного центра фермента. Такие ферменты получили название истинных металлоферментов. В других случаях ион металла не связан постоянно с ферментом и может функционировать только как мостик между ферментом и субстратом. В этом случае образование фермент-субстратного комплекса идет ступенеобразно. Чаще сначала ион металла соединяется с ферментом, а затем уже возникает комплекс фермент-металл-субстрат.
В качестве активаторов ферментов могут выступать также цистеин, свободные SH-группы. Активирующий эффект подобных соединений заключается в том, что они восстанавливают дисульфидные связи (– S – S – связи) неактивного фермента с образованием свободных SH-групп, которые необходимы ферменту для проявления его каталитической активности.
Другим примером активации является превращение неактивных
предшественников ферментов (проферментов, или зимогенов) в активные ферменты. В этом случае в основе активации лежит гидролиз одной или нескольких пептидных связей в молекуле профермента под действием протеолитических ферментов. Так, в состав полипептидной цепи профермента трипсина (протеолитического фермента животного происхождения) – трипсиногена – входят 229 аминокислотных остатков. В результате гидролиза пептидной связи между 6-м и 7-м аминокислотными остатками от полипептидной цепи трипсиногена отщепляется N-концевой гексапептид, после чего создаются условия для формирования активного центра трипсина.
2.1.8. Единицы активности ферментов.
Определение количественного содержания ферментов в биологических объектах представляет известные трудности, потому что за редким исключением ферменты в тканях присутствуют в ничтожно малых концентрациях. Поэтому непосредственно определяют не концентрацию фермента, а активность фермента. Об активности фермента судят по скорости ферментативной реакции, т.е. по скорости убыли субстрата или по скорости образования продуктов реакции.
Международный биохимический союз рекомендовал международную единицу активности фермента (
МЕ)
. Одна международная единица соответствует тому количеству фермента, которое катализирует превращение 1мкмоль субстрата за 1мин. В оптимальных для этого фермента условиях. Однако МЕ не может считаться единицей системы СИ, т.к. минута является внесистемной единицей.
Единицей активности ферментов в СИ является катал (кат) и его производные (мкат и др.). Один катал – это количество фермента, которое необходимо для каталитического превращения одного моля субстрата за 1 сек.
2.1.9. Внутриклеточная регуляция ферментативной активности.
1) Простейший тип регуляции ферментативной активности, т.е. регуляции метаболических процессов, затрагивает основные параметры, влияющие на скорость ферментативной реакции (рН среды, температура, концентрация субстрата и коферментов, присутствие ингибиторов и активаторов). Например, любой процесс, способный изменить реакцию среды внутри клетки, может влиять на скорость той или иной ферментативной реакции. Если в норме концентрация субстрата в клетке ниже значения константы Михаэлиса, то скорость ферментативной реакции будет резко изменяться в зависимости от содержания субстрата.
2) Другой общий путь регуляции метаболических процессов обеспечивается действием т.н. аллостерических ферментов. Важные представления о механизме этого явления были сформулированы французскими исследователями Ф. Жакобом, Моно и Шанже в 1963 г. Они остулировали положение, согласно которому снижение или повышение активности ферментов invivo обычно является следствием изменения стерической конфигурации – конформации молекул фермента в результате присоединения соответствующих ингибиторов или активаторов, в роли которых выступают различные метаболиты, к другим, некаталитическим участкам фермента. Эти участки получили название аллостерических или регуляторных центров (греч. аllos – иной, другой). В результате присоединения ингибитора к аллостерическому центру фермента (аллостерический ингибитор) взаимодействие его активного центра с субстратом становится невозможным, несмотря на то, что активные группировки в каталитическом участке фермента остаются неблокированными. Напротив, под влиянием аллостерического активатора происходят такие изменения конфигурации активного центра, которые приводят к повышению каталитической активности фермента.
3) Третьим типом регуляции ферментативной активности является изменение концентрации фермента в результате стимулирования или ингибирования его биосинтеза либо изменения скорости его катаболизма.
4) Кроме того, существует еще регуляция ферментативной активности с помощью взаимозаменяемых форм ферментов. Например, фосфорилаза мышечной ткани, катализирующая расщепление гликогена, может находится в двух формах – (а) и (б). В физиологических условиях активной является преимущественно фосфорилаза в форме (а). Но, под влиянием некоторых факторов (под действием гормонов, например) фосфорилаза (а) может превратиться в фосфорилазу (б).
2.1.10.Методы определения активности ферментов
Прежде чем преступить к выделению фермента, необходимо избрать и тщательно отработать метод определения активности, под контролем которого производится выбор наиболее эффективных приемов очистки ферментов, а затем и выполнение последовательных стадий его препаративного получения. Активность фермента меняется при различных условиях реакции и зависит от температуры, рН среды, от концентраций субстратов и кофакторов. Учитывая это, при определении активности фермента на разных стадиях очистки необходимо строго соблюдать одни и те же условия. Желательно не ограничиваться определением активности по одному какому-либо методу. Количество субстрата, превращаемого в условиях теста по определению активности фермента, должно быть пропорционально количеству последнего и времени инкубирования. Если же нет такой пропорциональности, то активность рассчитывают по предварительно построенному калибровочному графику, отражающему зависимость скорости реакции от количества единиц фермента. Когда ход реакции не линеен во времени, следует определять начальную скорость реакции (по тангенсу угла наклона касательной к начальному отрезку кривой превращения). Для этого легче всего применять такие методы изменения активности, которые позволяют непрерывно следить за ходом превращения во времени: спектрофотометрические методы, потенциометрические, полярографические и т.п. Для измерения скорости ферментативной реакции необходимо выбрать буфер, который не тормозит исследуемую активность и обеспечивает поддержание рН раствора, близкой к оптимальной для данного фермента. Реакцию проводят при температуре, лежащей в большинстве случаев в пределах 25-400С. При исследовании ферментов, требующих присутствия кофакторов (ионов металлов, коферментов и др.), концентрация которых может снижаться по мере очистки фермента, в реакционную смесь следует добавлять недостающие кофакторы, например соли металлов, АТФ, НАДФ и т.п. Также для определения активности ферментов вводят стабилизаторы в состав опытных смесей. Во многих случаях добавление желатина, альбумина и других добавок предотвращает денатурацию ферментного белка.
Спектрофотометрические методы. Спектрофотометрические методы основаны на поглощении света в определенных участках спектра многими соединениями, являющимися активными группами ферментов, субстратами или продуктами реакции. Положение максимума поглощения при определенной длине волны определяется наличием в исследуемом материале определенных групп - аналитических форм. Для измерения спектров используют специальные приборы - спектрофотометры, фотометрические абсорбциометры и др. Этот метод отличается высокой чувствительностью, быстротой определения, малым расходованием фермента и реактивов и позволяет следить за течением реакции во времени. Для этого реакционную смесь помещают в кювету, вставленную в термостатируемый кюветодержатель. Через малый промежуток времени после добавления фермента (или субстрата) и быстрого перемешивания измеряют поглощение при длине волны, характерной для используемого субстрата или конечного продукта, образующегося в данной реакции. С помощью спектрофотометрического метода можно измерять непосредственно концентрацию некоторых ферментов (после достаточной очистки) по величине характерных максимумов поглощения прочно связанных коферментов (простетических групп). Необходимо иметь произвольно выбранную единицу фермента, с помощью которой можно было бы количественно выразить чистоту и активность различных фракций. В большинстве случаев выбор единицы зависит от избираемого метода определения. В случае спектрофотометрического метода такой единицей может служить количество фермента, которое вызывает определенное изменение в оптической плотности за определенное время при данных условиях опыта; если определяется продукт реакции, то единицей будет количество фермента, которое вызывает образование определенного количества вещества в минуту, и т.д. Тогда удельную активность фермента, которая является мерилом чистоты ферментного препарата, выражают числом единиц в 1 мг вещества (белка).
Для целей определения ферментов могут быть использованы не только измерение поглощения света, но также измерения флюоресценции - спектрофлюорометрические методы. Такое определение активности фермента в ряде случаев по чувствительности превосходит спектрофотометрические методы на целый порядок величины. Некоторые коферменты и субстраты обладают сильной флюоресценцией. НАД и НАДФ в восстановленном состоянии имеют сильную флюоресценцию и не флюоресцируют в окисленном состоянии. Поэтому спектрофлюорометрию используют для изучения кинетики и механизма действия никотинамидных и флавиновых ферментов.
Колориметрические (фотометрические) методы. В основе этих методов лежит измерение при помощи визуального или фотоэлектрического колориметра окрашенного продукта, образующегося при взаимодействии субстрата или продукта действия фермента со специфическими реактивами, которые обычно добавляются в фиксированную опытную пробу, взятую после остановки ферментативной реакции. Эти методы весьма разнообразны. Разработаны количественные методы, основанные на биуретовой реакции, при которой состав белка, очевидно не должен сказываться на результатах определения, т.к. эта реакция на пептидные связи, а не на боковые группы в белке. Метод Горнелла и соавторов, основанный на измерении полосы поглощения в области 550-650 нм, используется для определения значительных количеств (1-10 мг) белка в пробе. Предлагаются биуретовые микрометоды, основанные на поглощении ультрафиолетовых лучей медно-белковыми комплексами: они позволяют определять 0.1 - 2.0 мг белка в пробе. Число небелковых веществ, которые могут влиять на определения с помощью биуретовой реакции, невелико, но следует вносить поправки на те вещества, которые присутствуют в высоким концентрациях (соли аммония, сахароза). Наиболее ценными являются те фотометрические методы, которые позволяют следить во времени за ходом ферментативной реакции без ее прекращения по изменению окраски хромогенного субстрата или добавленного индикатора. Метод Фолина и Чиокальто, предложенный для определения количества белка, основан на хромогенной природе некоторых боковых групп аминокислот, а также на присутствии в белках пептидных связей. Этот метод обладает высокой чувствительностью (на пробу достаточно 0.01 - 0.1 мг белка), но многие небелковые вещества мешают определению.
В настоящее время для определения количества белка широко пользуются измерениями интенсивности поглощения света при 280 нм, которое обусловлено присутствием в белке ароматических аминокислот. Количество этих аминокислот в разных белках различно и ,следовательно, интенсивность неодинакова. В кювете толщиной 1 см у раствора, содержащего 1 мг “усредненного” белка в 1 мл, оптическая плотность равна 1 при длине волны 280 нм. Нуклеиновые кислоты поглощают при 280 нм, но можно сделать поправку на их присутствие, проводя измерения и при 260 нм и при 280 нм. Очень важна быстрота измерения активности ферментов. То же относится и к измерению количества сухого остатка или количества белка. Тем самым предпочитают быстрый метод определения белка путем измерения величины поглощения при 280 нм. Выигранное время важнее, за счет того, что удельное поглощение выделяемого белка иногда значительно отличается от средней величины поглощения смеси белков, присутствовавших в исходном материале.
Манометрические методы. Эти методы используются при определении активности фермента в тех случаях, когда в исследуемых реакциях один из компонентов находится в газообразном состоянии. К таким реакциям относится главным образом те, которые связаны с процессами окисления и декарбоксилирования, сопровождающимися поглощением или выделением кислорода и углекислоты, а также реакции, в которых выделение или связывание газа происходит в результате взаимодействия продуктов ферментативного превращения с добавленным в систему реактивом. Наблюдение за ходом реакции во времени проводится в специальных приборах - манометрических аппаратах Варбурга.
Другие методы. Сюда относится обширный ряд методов, включающих поляриметрию, вискозиметрию и кондуктометрические измерения и т.п. Также определение активности можно выполнять, используя методы хроматографии и электрофореза на бумаге. Эти методы высокочувствительны и специфичны, что делает их во многих случаях незаменимыми; они позволяют значительно сократить расход фермента на измерение активности, но не всегда применимы ввиду продолжительности разделения веществ в процессе хроматографии (и электрофореза).
2.1.11. Получение ферментных препаратов.
Технические препараты ферментов. Комплексный амилолитический ферментный препарат получают путем выращивания плесневых грибов на твердой питательной среде с последующей сушкой и измельчением полученной массы. Более активный препарат фермента получают путем экстракции такого “грибного солода” с последующим выпариванием и сушкой. Еще более активные ферментные препараты можно выделить из культуральной жидкости путем осаждения амилазы ацетоном и дальнейшим высушиванием коагулятом при температуре 27-270С. Для осаждения фермента часто используют и сульфат аммония. Предварительно культуральную жидкость выпаривают при температуре 400 С до 40%-ного содержания сухих веществ. Коагулят сушат вместе с наполнителем.
Комплекс ферментов протеолитического и амилолитического действия получают при помощи культуры Bacillus subtilis. Это аэробные, грамположительные, подвижные палочки. Для этих бактерий характерен очень богатый комплекс гидролитических ферментов. В качестве источников питания они могут использовать белки, углеводы, спирты, органические кислоты. Bacillus subtilis культивируют как методом поверхностного культивирования на отрубях, так и в жидких средах особого состава по методу глубинного культивирования.
Целлюлолитические ферментные препараты. Производство целлюлаз основывается на использовании культуры гриба Trichoderma viride. Существующие в настоящее время способы получения целлюлаз в глубинной культуре предполагает выращивание микроорганизмов-продуцентов целлюлаз на питательной среде, содержащей в качестве источников углерода, как правило, очищенную целлюлозу, или же содержащие ее природные субстраты. Но получение целлюлазы с использованием в качестве основного компонента среды природной целлюлозы (например, древесные опилки) сопряжено с рядом технологических трудностей. Более рационально использование питательной среды, содержащей растворимый “индуктор”. Такой питательной средой может быть молочная сыворотка, основным компонентом которой является лактоза (предварительно от молочной сыворотки отделяют белок). В качестве продуцента может быть использован гриб Trichoderma lignorum, позволяющий получить весь комплекс целлюлолитических ферментов, необходимый для расщепления природных целлюлозусодержащих субстратов.
3.АМИЛОЛИТИЧЕСКИЕ ПРЕПАРАТЫ
Амилолитические препараты широко выпускаются в нашей стране и за рубежом. В основном это крупнотоннажное производство. Амилазы находят применение почти во всех областях, где перерабатывается крахмалсо-держащее сырье. Амилазы используют для осахаривания зернового и картофельного крахмала. Самым большим потребителем амилолитических ферментов является спиртовая и пивоваренная промышленности, где в настоящее время солод (проращенное зерно) успешно заменяется амилолитическими ферментными препаратами.
3.1. Источники получения амилаз
Амилазы очень широко распространены в природе. Они синтезируются многими микроорганизмами (бактерии, грибы, актиномицеты, дрожжи), животными и растениями. До развития ферментной промышленности главным промышленным источником получения амилаз в европейских странах было проросшее зерно (солод). Для медицинских целей амилазы получали из животного сырья. В настоящее время главным источником амилаз являются микроорганизмы, особенно бактерии, грибы и реже дрожжи.
3.2. Механизм действия и свойства амилаз
Субстратами для действия амилаз являются крахмал, состоящий из амилозы и амилопектина, продукты частичного гидролиза крахмала и гликоген. Крахмал
- растительный полисахарид с очень сложным строением. Это двухкомпонентное соединение, состоящее из 13-30% амилозы и 70-85% амилопектина. Оба компонента неоднородны, их молекулярная масса (М. м.)
колеблется в широких пределах и зависит от природы крахмала. Амилоза - это неветвящийся полимер, в котором остатки глюкозы соединены a-1, 4-гликозидной связью; степень полимеризации около 2000. В «аномальных» амилозах с одной-двумя a-1, 6-связями полимеризация может возрасти до 6000 (рис.1). Амилоза практически не обладает восстанавливающей способностью, так как в каждой молекуле амилозы имеется только одна свободная альдегидная группа.
Молекула амилозы представляет собой растянутую спираль, шаг которой составляет 10,6 А и в каждый виток входит 3 остатка глюкозы. Максимальная длина молекулы амилозы достигает 7000 А. В растворе спираль сжимается за счет увеличения витка, в котором уже участвует 6 остатков глюкозы. При вхождении молекул йода в спираль амилозы возникает характерный синий цвет. Строго говорить о величине молекулы амилозы нельзя, т. е. даже из одного образца крахмала извлекается амилоза, с величиной молекулы от 500 до 2000 остатков глюкозы. Амилопектин имеет большую молекулярную массу, чем амилоза, и более сложное строение. Это ветвящийся полисахарид. Предполагается, амилопектин ветвится дихотомически, т. е. число концевых звеньев всегда на единицу больше числа звеньев, дающих ветвление, а сумма этих чисел дает общее число звеньев по всей цепи (см. рис. 2.2).
Механизм действия. К группе амилотических ферментов относятся a- и b-амилазы, глюкоамилаза, пуллуланаза, изоамилаза и некоторые другие ферменты. Амилазы бывают двух типов: эндо- и экзоамилазы. Четко выраженной эндоамилазой является а-амилаза, способная к разрыву внутримолекулярных связей в высокополимерных цепях субстрата.
Глюкоамилаза и b-амилаза являются экзоамилазами, т. е. ферментами, атакующими субстрат с нередуцирующего конца.
При изучении механизма действия амилаз имеются определенные сложности, и прежде всего они заключаются в том, что субстрат - крахмал неоднороден и имеет различные характеристики по степени полимеризации гли-козидной цепи и количеству ветвлений.
Реакции, катализируемые амилазами, имеют две стадии: короткую -предстационарную и длительную - стационарную. Во время первой стадии эндоамилаза быстро уменьшает молекулярную массу субстрата, образуя смесь линейных и разветвленных олигосахаридов. Второй этап реакции продолжается, пока продукты гидролиза не перестанут окрашиваться йодом; он протекает значительно медленнее и зависит от индивидуальных свойств фермента и его природы. Поэтому конечные продукты гидролиза а-амилазами могут быть различными. Первая стадия воздействия фермента на субстрат хотя и носит неупорядоченный характер, имеет для всех видов a-амилаз схожий механизм.
Существует две гипотезы о механизме действия экзоамилаз на субстрат. Первая гипотеза предполагает, что, воздействуя на субстрат по одноцепочечному или «молниеобразному» механизму, экзоамилаза образует фермент-субстратный комплекс с захватом нередуцирующего конца цепи.
Дальнейшее продвижение фермента по этой цепи происходит до полного ее гидролиза. По второй гипотезе (b- и глюкоамилаза действуют на субстрат путем механизма множественной атаки, т. е. фермент образует комплекс с молекулой субстрата, затем через несколько этапов этот комплекс распадается и фермент связывается с новой молекулой субстрата. Иными словами, при множественной атаке происходит нечто среднее между неупорядоченным механизмом и одноцепочечной, «молниеобразной» атакой. Для полного гидролиза по этому механизму однамолекула субстрата должна образовывать много раз фермент-субстратные комплексы. При этом возможен гидролиз нескольких связей в одном каталитическом акте.
Механизм воздействия амилаз на субстрат может быть рассмотрен с нескольких позиций: вид разрываемой связи (a-1,4 или a-1,6); тип воздействия на субстрат (эндо- или экзо-); влияние на скорость гидролиза степени полимеризации субстрата;
4) возможность гидролиза олигосахаридов;
способность фермента к множественной атаке субстрата.
Наличие признаков амилаз, отраженных в 3 и 4 позициях, при действии на линейные субстраты может свидетельствовать о существовании у этих ферментов подцентровой структуры. Вероятно, активный центр амилазы может состоять из нескольких подцентров, каждый из которых может вступать в контакты с глюкозным остатком. Энергия взаимодействия (А;), выраженная в единицах свободной энергии (кДж/моль), определяет подцентровое сродство фермента к субстрату. Это сродство индивидуально и может быть как положительным, так и отрицательным. Вероятность существования подцентровых структур амилаз помогает установить строение активного центра амилаз, дает более четкое объяснение субстратной специфичности, но не дает объяснений механизма гидролиза разветвленных субстратов.
a-Амилаза. а-Амилаза (а-1,4-глюкан-4-глюканогидролаза, КФ 3.2.1.1.) является эндоамилазой, вызывающей гидролитическое расщепление a-1,4-гли-козидных связей внутри высокополимеризованного субстрата. Фермент назван a-амилазой потому, что он высвобождает глюкозу в a-мутамерной форме.
a-Амилаза - водорастворимый белок, обладающий свойствами глобулина и имеющий М. м.
45 000-60 000. Своего рода исключением является a-амилаза В. macerans, которая имеет М. м.
130 000. Есть указания, что некоторые термостабильные a-амилазы имеют М. м. 14 000-15 000, но в их молекулах содержится в 2-3 раза больше атомов кальция.
Все а-амилазы относятся к металлоэнзимам, содержание в них Са колеблется от 1 до 30 г атом на 1 г моль фермента. Полное удаление кальция приводит к инактивации фермента. Повторное введение кальция в среду может частично восстановить его активность. a-Амилаза В. subtilis с помощью иона цинка способна образовывать димерную форму, чего лишены другие a-амилазы. Все a-амилазы устойчивы к воздействию протеаз. Они богаты тирозином и триптофаном. Глютаминовая и аспарагиновая кислоты составляют 25% массы белка. Наличие этих кислот в а-амилазе связывают с их оса-харивающей способностью. Так, разжижающие a-амилазы не имеют сульфгидрильных групп, а осахаривающие содержат один остаток цистеина. Сравнительно мало или совсем отсутствуют в а-амилазах содержащие серу аминокислоты. Некоторые a-амилазы грибного происхождения имеют углеводный фрагмент, в состав которого могут входить манноза, ксилоза, гексозоамин, но функции его не установлены.
В зависимости от вида микроорганизма свойства ос-амилаз могут сильно отличаться не только по механизму воздействия на субстрат и конечным продуктам, но и по оптимальным условиям для проявления максимальной активности.
Действуя на целое крахмальное зерно, a-амилаза атакует его, разрыхляя поверхность и образуя каналы и бороздки, т.е. как бы раскалывает зерно на части. Клейстеризованный крахмал гидролизуется ею с образованием на окрашиваемые йодом продукты - в основном состоящие из низкомолекулярных декстринов. Процесс гидролиза крахмала многостадийный. В результате воздействия a-амилазы на первых стадиях процесса в гидролизате накапливаются декстрины, затем появляются неокрашивающиеся йодом тетра- и тримальтоза, которые очень медленно гидролизуются a-амилазой до ди- и моносахаридов.
Все a-амилазы проявляют наименьшее сродство к гидролизу концевых связей в субстрате. Некоторые же а-амилазы, особенно грибного происхождения, на второй стадии процесса гидролизуют субстрат более глубоко с образованием небольшого количества мальтозы и глюкозы. Схему гидролиза под действием а-амилазы можно записать следующим образом:
а-Амилаза
Крахмал,---------------->
а-Декстрины + Мальтоза + Глюкоза
гликоген (много) (мало) (мало)
b-Амилаза. b-Амилаза (a-1,4-глюкан мальтогидролаза, КФ 3.2.1.2) - активный белок, обладающий свойствами альбумина. Каталитический центр фермента содержит сульфгидрильные и карбоксильные группы и имидозольный цикл остатков гистидина. b-Амилаза -экзофермент концевого действия, проявляющий сродство к предпоследней b-1,4-связи с нередуцирующего конца линейного участка амилозы и амилопектина.
В отличие от a-амилазы b-амилаза практически не гидролизует нативный крахмал, тогда как клейстеризованный крахмал гидролизуется ею с образованием мальтозы b-конфигурации, поэтому данная амилаза по аналогии с a-амилазой называется b-амилазой. Если гидролизу подвергается амилоза, то гидролиз идет полностью до мальтозы. Незначительное количество декстринов может образовываться при гидролизе «аномальных» амилоз, так как гидролиз b-амилазой идет только по линейной цепи до a-1,6-связей. Если субстратом для b-амилазы служит амилопектин, то гидролиз идет в значительно меньшей степени. b-Амилаза отщепляет фрагмент с нередуцирующего конца участка от внешних линейных ветвей, имеющих по 20-26 глюкозных остатков, с образованием 10—12 молекул мальтозы. Гидролиз приостанавливается на предпоследней a-1,4-связи, граничащей с a-
1,6-связью. В гидролизате накапливается 54-58% мальтозы, остальное составляют высокомолекулярные декстрины, содержащие значительное количество а-1,6-связей - так называемые b-декстрины. Действие b-амилазы на крахмал можно записать в виде следующей схемы:
b-Амилаза
Крахмал,---------------->
Мальтоза + р-Декстрин
гликоген (54-58%) (42-46%)
b-Амилазы проявляют большую стабильность в отсутствие ионов Са2+
.
Молекулярная масса b-амилазы растений достаточно высока, она составляет от 50 000 до 200 000. Фермент может состоять из одной или четырех субъединиц до 50 000 каждая. Фермент содержит SH-группы и чувствителен к действию тяжелых металлов. Считается, что (b-ами-лаза обладает высокой способностью к множественной атаке субстрата. Для амилозы средней молекулярной массы в одном присоединении фермента к субстрату возможно отщепление до четырех остатков мальтозы. При увеличении молекулярной массы субстрата возможно и большее количество мест атаки.
Глюкоамилаза. Глюкоамилаза (а-1,4-глюкан глюкогидролаза, КФ 3.2.1.3.) широко распространена в природе. Она синтезируется многими микроорганизмами и образуется в животных тканях, особенно в печени, почках, плаценте кишечника и т. д. Фермент в литературе известен под различными названиями: амилоглюкозидаза, g-амилаза, лизосомальная a-глюкозидаза, кислая мальтаза, матулаза и экзо-1,4-a-глюкозидаза. Глюкоамилаза катализирует последовательное отщепление концевых остатков a-D-глюкозы с нередуцирующих концов субстрата. Это фермент с экзогенным механизмом воздействия на субстрат. Многие глюкоамилазы обладают способностью так же быстро, как и a-1,4-связь, гидролизовать a-1,6-глюкозидные связи. Но это происходит только в том случае, когда за a-1,6-связью следует a-1,4-связь, поэтому декстран ими не гидролизуется. Отличительной особенностью глюкоамилаз является способность в десятки раз быстрее гидролизовать высокополимеризованный субстрат, чем олиго- и дисахариды.
В литературе высказывается мнение, что механизм атаки субстрата глюкоамилазой может быть двух типов: либо одноцепочечный, либо множественной атаки, и что активный центр имеет подцентровую структуру.
Почти все глюкоамилазы являются гликопротеидами, содержащими от 5 до 35% углеводов, которые состоят из олиго-, ди- и моносахаридов. Углеводный компонент может быть целостным фрагментом или же разбитым на индивидуальные соединения, которые прикрепляются к белку через треонин и серин. Например, у глюкоамилазы A. niger их 20. Большинство известных глюкоамилаз имеет оптимум рН при 4,5-5,2, реже - при 5,7-6,0, в основном для дрожжевых глюкоамилаз.
РН-стабильность микробных глюкоамилаз лежит в широком диапазоне -от 2,5 до 9. Термостабильность глюкоамилаз лежит в интервале от 30 до 45°С и редко повышается до 55-60°С. Глюкоамилазы различного происхождения заметно отличаются по молекулярной массе, которая, по данным различных авторов, имеет значения от 48 000 до 210 000. Следует заметить, что далеко не все микробные глюкоамилазы способы полностью гидролизовать крахмал до глюкозы. Еще в 60-х годах И.Фукумото предложил все микробные глюкоамилазы разделить на два типа: полностью гидролизующие крахмал до глюкозы и гидролизующие крахмал до глюкозы на 80-85%.
В то время предполагалось, что степень гидролиза зависит только от свойств глюкоамилаз и их происхождения. Позже было показано, что при росте культуры параллельно накапливаются и другие амилолитические ферменты, обладающие не только гидролитическим, но и трансферазным действием. Это гликозилтрансфераза и a-амилаза. Даже в случае, если система открытая и продукт гидролиза (глюкоза) постоянно удаляется из системы, процесс может дойти до полного гидролиза крахмала до глюкозы. Если же система закрытая и концентрация субстрата велика, то при достижении определенной концентрации глюкозы в реакционной среде в результате переноса глюкозильных остатков на глюкозу, ди- и олигосахариды начинают накапливаться изомальтоза, паноза, нигероза, изомальтотриоза и другие сахара, которые имеют горький вкус. В результате процесс не может дойти до полного превращения крахмала в глюкозу и возникает ошибочное представление, что глюкоамилаза не полностью гидролизует крахмал. Сама же глюкоамилаза может проявлять небольшую трансферазную активность, но только при концентрации глюкозы свыше 60-70%. Поэтому ранее принятое деление глюкоамилаз на два типа следует считать необоснованным.
3.3. Компьютерное моделирование структуры амилолитических ферментов
Используя данные рентгеноструктурного анализа и программу MolScript, методом компьютерного моделирования построена трехмерная модель макромолекулы глюкоамилазы из Aspergillus awamori, что позволило по-новому оценить многочисленные экспериментальные данные, полученные классическими методами (электронная спектроскопия, ЯМР, ИК спектроскопия и т.д.). Показаны особенности строения активного центра данного фермента, обозначены функционально важные группы. Внесены дополнения в представления о механизме катализа амилолитическими ферментами.
При изучении механизмов ферментативного катализа классическими методами (ИК и УФ- спектроскопия, электронная спектроскопия, ЯМР и т.д.) дополнительную информацию вносит компьютерное моделирование белковых молекул с использованием данных рентгеноструктурного анализа (РСА). Способность глобулярных белков образовывать качественные монокристаллы, в которых все атомы идентичны и ориентированы одинаковым образом, позволила изучать их с помощью метода кристаллографии. Известно, что данные рентгеноструктурного анализа всегда правильно отражают укладку белковой цепи, а в подавляющем большинстве случаев буквально воспроизводят активную конформацию фермента. Следовательно, они являются надежной и пока что единственной основой для количественного описания механизма каталитического акта на атомно-молекулярном уровне. Метод кристаллографии позволяет расшифровывать трехмерные структуры комплексов ферментов с ингибиторами, которые в той или иной мере соответствуют химическим стадиям катализа. Располагая рентгеноструктурными данными, одна часть которых отвечает реальному исходному состоянию фермента, а другая - фермент-ингибиторным комплексам, становится возможным воссоздание механизма каталитического акта, а, следовательно, можно проследить динамику поведения белковой макромолекулы. Разрабатываемые в настоящее время программы компьютерного моделирования белковых молекул на основе результатов кристаллографии позволяют получать наглядные трехмерные изображения исследуемых объектов, и по-новому осмыслить данные экспериментальных исследований кинетико-термодинамических свойств ряда ферментов, полученные за последнее десятилетие, а, следовательно, выявить дополнительные аспекты молекулярных механизмов ферментативного превращения субстрата. В этой связи целью данной работы является анализ топологии амилолитических ферментов с использованием программы MolScript по данных рентгеноструктурного анализа. Повышенный интерес к амилазам обусловлен
их широким применением в медицине, пищевой и легкой промышленности в
качестве эффективных биокатализаторов. Исследования физико-химических свойств, кинетико-термодинамических параметров, особенностей фермент-субстратных взаимодействий при осуществлении акта катализа амилолитическими ферментами способствовали существенному изменению и усовершенствованию многих существующих технологий и созданию новых. Однако, очевидным является недостаток информации о пространственной организации макромолекул ферментов данной группы.
Объектом наших экспериментальных исследований является глюкоамилаза из Aspergillusawamori, препарат Г-20Х производства Ладыжинского завода ферментных препаратов. Для определения активности глюкоамилазы использовали глюкозооксидазный метод. Определение общего количества белка в ферментном препарате проводили по методу Лоури. Исследование соотношения типов вторичной структуры осуществляли методом инфракрасной спектроскопии на спектрометре SpecordM-80 (Германия). Определение молекулярной массы глюкоамилазы, а также изучение четвертичной структуры фермента с использованием додецилсульфата натрия в концентрации 3,5×10-5
моль/л для разрушения надмолекулярного уровня организации, проводили с помощью гель-хроматографии на сефадексе G-200. Количественное определение SH-групп осуществляли методом, основанном на измерении прироста оптической плотности в области 255 нм при присоединении n-меркурибензоата к SH-группам. Для количественного определения S-S групп использовали метод, основанный на измерении оптической плотности (l=412нм.) при образовании нитротиобензоата. Определение числа ионогенных групп проводили на автоматической установке, разработанной на кафедре биофизики и биотехнологии ВГУ, путем сравнения кривых титрования белка. Изучение пространственной структуры глюкоамилазы осуществляли с использованием программы MolScript.
Программа MolScript используется для моделирования протеиновых и протеидовых структур. Исходными данными для программы является *.in
файл, в котором указывается файл рентгеноструктурного анализа, объекты для вывода и точное их описание с помощью графических параметров. Для создания в первом приближении входных файлов MolScript используется программа MolAuto. Эта программа распознает вторичную структуру в файле pdb и выводит соответствующие команды для схематического описания данной структуры. MolAuto имеет несколько ключей, позволяющих влиять на свойства объектов. Программа не способна выбирать удобный ракурс, поэтому пользователю необходимо выбирать его самостоятельно. MolAuto может быть частью потока UNIX с программой MolScript: например для быстрого получения изображения структуры в режиме OpenGL. Каждый формат, поддерживаемый MolScript, обладает своими специфическими свойствами. Поэтому выбор формата производится с учетом установленного программного обеспечения и желаемого набора свойств полученной модели.
Программа MolScript использует одну фиксированную правостороннюю
систему координат с ценой деления 1 ангстрем. Точка обзора зафиксирована и измениться не может. Для просмотра молекулы с разных ракурсов атомные координаты необходимо трансформировать, т.е. повернуть и/или транслировать в пределах системы координат. Молекулярные координаты считываются из pdb файла, а сами координаты атомов и остатков, входящих в данный файл, используются графическими командами. Координаты выбранных атомов трансформируются матрицей, определенной одной или несколькими операциями.
При изучении топологии для двумерных статических и трехмерных моделей проводилось распознавание субъединиц ферментов в разных масштабах по различным направлениям. Топологические аспекты рассматривались как для фрагментов, так и для целостных молекул амилолитических ферментов, что позволило ввести обозначения функционально значимых групп.
Классические методы изучения особенностей протекания ферментативного катализа позволяют рассматривать широкий круг вопросов. Так, экспериментальным путем нами было исследовано влияние различных физико-химических факторов на конформацию макромолекулы глюкоамилазы из Aspergillusawamori и связанных с ней механизмов активации и инактивации данного фермента; изучены кинетико-термодинамические аспекты реакции гидролиза крахмала. Методом гель-хроматографии определена молекулярная масса глюкоамилазы, составляющая 106 кДа. Исследование аминокислотного состава глюкоамилазы показало, что молекула фермента содержит 26,8% аминокислотных остатков с алкильными боковыми цепями (Ala, Val, Leu, Met, Ile), определяющими высокую устойчивость фермента к органическим растворителям. Обнаружено высокое содержание в молекуле глюкоамилазы Ser и Thr. Результаты наших экспериментов и анализ данных литературы по секвенированию глюкоамилазы (А.Е. Алешин и др.,1994) позволяют сделать
заключение о том, что молекула фермента имеет 6 SH-групп и 4 дисульфидных связи, стабилизирующих его третичную структуру. Изучение надмолекулярного уровня организации глюкоамилазы показало наличие четвертичной структуры, представленной двумя идентичными субъединицами с молекулярной массой 53600Да, обладающими каталитической активностью. Ее стабильность определяется водородными связями и гидрофобными взаимодействиями и может зависеть от небольших изменений в пространственной структуре каждой из взаимодействующих субъединиц. Причем четвертичная структура, по-видимому, обладает способностью передачи структурных перестроек от одной субъединицы к другой и является одним из факторов регуляции функциональной активности молекулы белка.
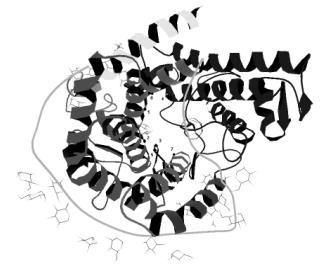
Рис. 1. Пространственная структура глюкоамилазы
Результаты исследований влияния n-меркурибензоата на каталитическую активность глюкоамилазы позволяют сделать заключение о том, что в состав активного центра данного фермента не входят имидазольная группа гистидина и SH-группы, а в гидролизе крахмала принимают участие карбоксильные группы аспарагиновой и глутаминовой кислот.
Используя данные рентгеноструктурного анализа и программу MolScript, методом молекулярного моделирования показано, что активный центр глюкоамилазы расположен в сквозной полости (~ 1,5нм), ограниченной следующими аминокислотными остатками: Leu-58, Leu-130, Leu-177, Leu-319, Trp-178, Trp-417, Phe-187 (рис1).
Установлено, что в гидролизе крахмала принимают участие карбоксильные группы Asp-55, Glu-179, Glu-400 (рис.2).
Активный центр фермента располагается в щели, прикрытой двумя кластерами молекул H2
O: первый - в области Leu-58 (12 молекул H2
O), второй – в углублении активного центра (7 молекул). Показано, что с одной стороны щели активного центра сосредоточены Asp-55 и Glu-179, а с противоположной – Glu-400.
Результаты рентгеноструктурного анализа (Алешин А.Е. и др, 1994 [10]) показали, что поверхность молекулы глюкоамилазы гликозилирована: 10 аминокислотных остатков серина и треонина имеют маннозу (Ser-453, Ser-455, Ser-459, Thr-457 и др.).
Данный фермент характеризуется наличием N-терминального домена, состоящего из 440 аминокислотных остатков, O-гликозилированного участка, имеющего 70 аминокислот и C-терминального крахмалсвязывающего домена (100 аминокислот). Каталитический домен имеет 2 N-гликозилированных участка, причем контакт между N-гликозилированными цепями и полипептидом стабилизируется остатком маннозы с помощью водородной связи или ионизированными молекулами воды, чем и определяется стабильность фермента и возможность образования надмолекулярных структур. O-гликозилированный домен имеет остатки Gly перед и после C-конца, которые представляют собой изгибы, определяющие взаимодействие крахмалсвязывающего домена с каталитическим и соответствующую ориентацию молекулы субстрата. Гликозилирование предотвращает скопление молекул субстрата в связывающих центрах и обеспечивает стехиометрическое связывание.
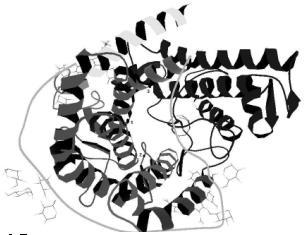
Рис. 2. Пространственная структура макромолекулы глюкоамилазы
Методом компьютерного моделирования установлено, что для молекул глюкоамилазы характерна плотная упаковка гидрофобного ядра в виде 13 α-спиральных участков (рис.3), а так же антипараллельных β-структур, образующих 11 петель (рис.4).

Рис. 3. Топология
α-спиралей в макромолекуле глюкоамилазы
|
Рис. 4. Топология β-слоев в макромолекуле глюкоамилазы |
Во вторичной структуре присутствуют, кроме того, неупорядоченные участки в количестве 19 (рис.5). В процентном соотношении данных структур по протяженности, аморфные участки преобладают над остальными, что подтверждено также методом ИК-спектроскопии (таблица).
Рис. 5. Топология неупорядоченных участков в макромолекуле глюкоамилазы
Таким образом, компьютерное моделирование позволяет получать дополнительную информацию о структуре белковых макромолекул и проводить расшифровку механизма ферментативного катализа.
В настоящее время продолжаются работы по проблемам изучения топологии различных амилолитических ферментов, таких как α-амилаза, β-амилаза, инулаза и др. Обсуждаются вопросы топологической самоорганизации исследуемых белковых макромолекул, определяемой взаимосвязью первичной и вторичной структуры.
4. ПОЛУЧЕНИЕ МИКРОБНЫХ ФЕРМЕНТНЫХ ПРЕПАРАТОВ
4.1. Основные сведения
Микроорганизмы способны синтезировать разнообразные ферменты. В зависимости от состава питательной среды и условий культивирования они легко переключаются с синтеза одного фермента на синтез другого. У микроорганизмов сравнительно короткий цикл развития (10... 100 ч), вследствие чего можно получать сотни «урожаев» в год.
Продуцентами ферментов могут быть бактерии, грибы, дрожжи и актиномицеты. Для промышленного получения ферментных препаратов используют как природные штаммы микроорганизмов, выделенные из естественных сред, так и мутантные, отселекционированные в результате воздействия на природные штаммы физических и химических мутагенов.
Микроорганизмы синтезируют одновременно комплекс ферментов, но некоторые из них, особенно мутантные штаммы, продуцируют один фермент в значительных количествах. Для лучшего использования крахмалсодержащего сырья в спиртовом производстве осахаривающие материалы должны содержать не только амилолитические ферменты, но и ферменты, гидролизующие другие углеводы сырья — целлюлозу и гемицеллюлозы. Для обеспечения дрожжей азотистым питанием имеют значение и протеолитические ферменты.
Несмотря на то, что для успешного осахаривания нужен комплекс ферментов, отбор микроорганизмов-продуцентов до сих пор проводился главным образом по высокой активности амилолитических ферментов — a-амилазы и глюкоамилазы.
4.2. Микроорганизмы – продуценты ферментов
Наиболее часто в качестве продуцентов амилолитических ферментов в спиртовом производстве используют микроскопические грибы, реже — дрожжеподобные организмы и споровые бактерии.
4.2.1. Микроскопические грибы
Для получения амилаз широко применяют микроскопические грибы рода Aspergillus, видов: niger, oryzae, usamii, awamori, bata-tae, рода Rhizopus, видов: delemar, tonkinensis, niveus, japonicum и др., а также отдельные штаммы Neurosporagrassa и Mucor.
Микроскопические грибы очень широко распространены в природе; основное место их обитания — почва. Несмотря на наличие многих родов и видов микроскопических грибов, все они характеризуются нитевидным строением тела и специфическим строением плодоносящих органов. Тело гриба состоит из длинных переплетенных нитей сероватого или белого цвета, называемых гифами. Они распространяются по поверхности питательного субстрата, образуя мицелий, и частично врастают в него. Некоторые гифы, поднимающиеся над поверхностью в виде легкого пушка, имеют более сложное строение и представляют собой органы плодоношения, называемые конидие- или спорангиеносцами. У мукоровых грибов на конце спорангиеносца находится шаровидное вздутие, окруженное оболочкой, внутри которого образуются споры. У аспергиллов конец конидиеносца имеет булавовидное утолщение, от которого отходят удлиненные клетки, называемые стеригмами; от стеригм отшнуровываются более мелкие круглые клетки — конидии.
Отделившиеся конидии или споры, попадая в благоприятные условия, начинают прорастать, затем гифы ветвятся, образуя мицелий; при истощении питательных веществ в среде гриб переходит в стадию споро- или конидиеобразования. Споры и конидии микроскопических грибов содержат пигменты, что и придает зрелым культурам характерную окраску.
Аспергиллы — типичные аэрофилы, поэтому они могут развиваться только на поверхности твердой или жидкой среды или в жидкой, достаточно аэрируемой среде. Оптимальная температура для большинства аспергиллов 25...30 "С, для некоторых — до 35 °С. Большинство грибов при поверхностном культивировании могут переносить кратковременное повышение температуры до 40 °С и даже 45 °С без заметной потери активности ферментов. Оптимальная влажность среды для них около 65 %.
Для питания аспергиллов необходимы углеводы, азотистые и минеральные вещества. В качестве источника углевода, кроме моносахаридов, многих олиго- и полисахаридов, могут служить спирты и органические кислоты, однако для накопления амилазы в среде обязательно должны присутствовать крахмал, декстрины или мальтоза. В средах, содержащих другие сахара, в том числе глюкозу, грибы не образуют амилазы. Источником азота могут быть белки и их гидролизаты, аммонийные соли и нитраты.
Среда должна содержать соединения, в состав которых входят сера, фосфор, калий, магний и микроэлементы. Большинство микроскопических грибов усваивают серу из сульфатов, а фосфор — из фосфатов. Аспергиллы не нуждаются в готовых витаминах и факторах роста, так как способны сами синтезировать их из более простых соединений, имеющихся в среде. Препараты ферментов из микроскопических грибов включают, как правило, широкий набор ферментов, поэтому могут полностью заменять зерновой солод.
На спиртовых заводах стали широко применять высокоактивный по глюкоамилазе штамм Asp. awamori 466 и ВУД-Т2, выращиваемые на концентрированном кукурузном сусле (18 % сухих веществ). Готовая культура имеет активность до 250 ед. ГлА на 1 мл, других ферментов образует мало.
4.2.2. Дрожжеподобные организмы
Амилолитические ферменты синтезируют также некоторые дрожжи и дрожжеподобные грибы родов Saccharomyces, Candida, Endomycopsis и Endomyces.
В спиртовом производстве нашли применение End. bispora и End. species 20-9, выращиваемые глубинным способом и продуцирующие главным образом активную глюкоамилазу; ос-амилаз-ная активность проявляется слабо. Высокоактивный End. bispora имеет разветвленный мицелий, образует бластоспоры; гифы — септированные, зернистые; на твердых агаризованных средах образуют колонии с воздушным серовато-белым мицелием, на жидких питательных средах — гифы и некоторое количество бластоспор.
Дрожжеподобные грибы в спиртовом производстве самостоятельно не применяют, так как они не содержат других ферментов, необходимых для нормального осахаривания сусла из крах-малсодержащего сырья. Обычно их используют в смеси с ферментными препаратами из микроскопических грибов или бактерий.
4.2.3. Бактерии
Активные амилазы способны синтезировать многие бактерии: Вас. subtilis, Вас. diastaticus, Вас. mesentericus, Вас. macerans и Вас. polymycus и др.
Бактерии — продуценты амилолитических ферментов представляют собой палочки длиной 1,2...1,3 мкм и диаметром 0,6...0,8 мкм. Палочки соединяются по две, три, иногда образуют цепочки. Цикл развития бактерий короче, чем микроскопических и дрожжеподобных грибов. Например, культуру Вас. diastaticus выращивают в глубинных условиях при температуре 60 °С в течение 10... 12 ч. Бактерию Вас. subtilis-82, применяемую в настоящее время на спиртовых заводах как продуцент a-амилазы в смеси с препаратами глюкоамилазы, выращивают в течение 48...60 ч при температуре 30...35 'С.
Особенность бактерий — их способность образовывать высокоактивную термостойкую a-амилазу, необходимую для разжижения и декстринизации крахмального клейстера на стадии под-варивания замесов и осахаривания сусла.
4.3. Производственные способы культивирования микроорганизмов – продуцентов ферментов
Существует два способа культивирования микроорганизмов — продуцентов ферментов: поверхностный и глубинный.
Первый способ, применяемый для культивирования микроскопических грибов, характеризуется развитием мицелия на поверхности твердого или жидкого субстрата. На жидком субстрате образуется пленка мицелия, продуцирующего не только амилолитические ферменты, но и органические кислоты, инактиви-рующие их, поэтому используют твердые субстраты с развитой поверхностью — пшеничные отруби, дробину барды, картофельную мезгу и др.
Максимальная активность ферментов достигается при культивировании грибов на пшеничных отрубях. Дробина барды бедна питательными веществами, и активность ферментов в культурах грибов, выращенных на ней, в 4...5 раз ниже, чем на отрубях. Зрелая культура грибов вследствие обволакивания частиц отрубей мицелием имеет вид плотной войлокообразной массы.
При поверхностном культивировании пшеничные отруби должны быть увлажнены и простерилизованы. В стерильных условиях готовят посевную культуру, но выращивают грибы в нестерильных условиях в кюветах, устанавливаемых в негерметичных растильных камерах. Теплоту, выделяющуюся в процессе роста грибов, удаляют продуванием через растильную камеру стерильного кондиционированного воздуха.
Поверхностный способ выращивания микроскопических грибов имеет ряд преимуществ. Так как во время роста гриба отруби не перемешиваются, посторонние микроорганизмы не распространяются по всей их массе и вызывают лишь незначительное местное инфицирование, которое, как правило, не влияет на активность ферментов. Это, однако, не исключает необходимости тщательной стерилизации воздуха, среды и оборудования. Культуру на отрубях высушивают до содержания влаги 10... 11 %. В таком виде она может храниться продолжительное время без значительной потери активности. Это позволяет организовать централизованное снабжение спиртовых заводов сухой культурой микроскопических грибов, что является одним из преимуществ поверхностного способа выращивания.
Недостаток поверхностного способа — необходимость устанавливать множество кювет, работу с которыми трудно механизировать. Себестоимость культуры гриба-продуцента высока,
причем в основном из-за затраты большого количества ручного труда. Механизация процесса выращивания возможна путем создания непрерывнодействующих установок или бескюветных аппаратов с вертикальным толстым слоем питательной среды и интенсивным продуванием воздуха через этот слой.
Глубинную культуру микроорганизмов выращивают на жидкой питательной среде при энергичной аэрации в герметически закрытых аппаратах и в стерильных условиях. Процесс полностью механизирован. Стерильность глубинной культуры микроорганизма — продуцента ферментов положительно отражается на результатах сбраживания сусла дрожжами.
4.3.1. Поверхностное культивирование микроскопических грибов
Питательные среды. При поверхностном культивировании твердой средой обычно служат пшеничные отруби, содержащие в достаточных количествах все вещества, необходимые для питания грибов. Лучшие результаты достигаются при содержании не менее 16 % крахмала в отрубях или в другой питательной среде.
Получение посевного материала. Для засева производственной питательной среды может быть использован посевной материал трех видов: споры, мицелий и спороносящая культура, выращенная на твердой питательной среде.
Споровый материал готовят поверхностным культивированием гриба на твердой или жидкой питательной среде в специальных аппаратах с кюветами. В первом случае гриб выращивают в пробирке на косом агаре до обильного образования спор, затем спорами засевают колбы со стерильными пшеничными отрубями, наконец, спороносящую культуру передают в специальный аппарат. По окончании спорообразования в этом аппарате отбирают споры посредством специального вибросепаратора при тщательной аспирации. Споры фасуют в полиэтиленовые мешочки, стеклянные или дюралюминиевые банки, в которых они могут сохраняться при температуре от 8 до 24 °С около 1,5 лет.
Во втором случае из спор, собранных в пробирке с косым агаром, готовят водную суспензию и ею засевают жидкую питательную среду, которую направляют в растильный аппарат. Из-под выросшей спороносящей пленки удаляют жидкую среду, пленку подсушивают, снимают с кювет, измельчают и упаковывают. Готовый споровый материал не содержит примеси среды и обладает высокой всхожестью (90...95 %), которая сохраняется свыше 3 лет.
Споры (конидии), полученные любым методом, обладают водоотталкивающей способностью и почти не смачиваются водой. Это свойство приводит к неравномерности засева среды и, следовательно, к неодинаковой скорости роста культуры. Смачиваемость спор можно улучшить, если добавить в их водную суспензию 25...50 мг поверхностно-активного вещества, например ал-килбензолсульфата на 1 г спорового материала. При этом всхожесть спор остается без изменения.
Для сокращения длительности культивирования микроскопических грибов в производственных условиях можно применять предварительное проращивание спор в течение 7 ч в жидкой питательной среде до появления ростовых трубочек. При этом продолжительность лаг-фазы сокращается на 8...9 ч и увеличивается оборачиваемость растильных камер.
При использовании спорового материала упрощается технологический процесс, появляется возможность более полно механизировать его, а также сократить площадь цеха чистой культуры. Централизованное производство спорового материала для группы предприятий, работающих с данным штаммом гриба, выгодно создать в одном специализированном цехе чистой культуры. Положительный опыт подобной организации имеется в производстве лимонной кислоты биотехнологическим способом.
Исследователями б. ВНИИПрБ было предложено засевать производственную питательную среду мицелием, полученным глубинным культивированием гриба или бактерий в колбах на качалке.
При высокой производительности предприятия целесообразно выращивать посевной мицелий в небольших ферментерах, как это было предложено А. П. Левчиком и осуществлялось на Мичуринском экспериментальном ферментно-спиртовом заводе.
На спиртовых заводах получило распространение приготовление посевного материала в виде поверхностной культуры гриба на пшеничных отрубях, которое складывается из следующих двух операций:
выращивание спороносной культуры гриба в пробирках на сусло-агаре;
засев спорами пшеничных отрубей в колбах и выращивание культуры гриба.
Культуру получают из лаборатории чистых культур ВНИИПрБ. Исходная — музейная чистая культура хранится в холодильнике при температуре от 2 до 4 °С. Один раз в месяц пересевают споры музейной культуры из пробирки в пробирку с сусло-агаром с соблюдением правил проведения микробиологических работ. При этом первая засеянная пробирка является музейной, а остальные две-три пробирки используют для дальнейшего размножения. Засеянные пробирки помещают в термостат при температуре 30 °С и выдерживают в нем в течение 4...6 сут.
При нормальном росте поверхность сусло-агара равномерно покрывается спороносящим мицелием. Культура гриба с плохим спороношением, медленно растущим мицелием, пушком вторичного роста в производство не допускается. Пробирки с вырос-
шей культурой вынимают из термостата и хранят в холодильнике. Перед посевом в колбы проверяют чистоту культуры высевом в чашки Петри и микроскопированием.
Чистая культура на сусло-агаре служит исходным материалом для размножения грибов на пшеничных отрубях в колбах. Отруби смешивают с водопроводной водой в соотношении 1:0,7 и переносят в несколько колб по 10...20 г в каждую. Колбы закрывают ватными пробками и стерилизуют в автоклаве при избыточном давлении 0,15 МПа в течение 1 ч. После охлаждения до комнатной температуры в колбы вносят суспензию спор гриба, содержащую 150...200 тыс. спор в 1 мл. Для этого в пробирку с культурой гриба на сусло-агаре прибавляют 10 мл стерильной водопроводной воды и при помощи стерильной пипетки над огнем соскабливают конидии гриба. Полученную суспензию используют для посева (2...2,5 % массы засеваемой среды или 0,4...0,5 мл на 10 г отрубей).
Выращивание ведут в термостате при температуре 30 °С в течение 4...5 сут. Одну из колб предназначают для дальнейшего размножения чистой культуры, остальные колбы хранят в холодильнике до последующего цикла. В колбу, используемую для размножения, вливают 50...60 мл стерильной водопроводной воды и над пламенем стерильной пипеткой тщательно перемешивают.
Расход чистой культуры при пересеве из колбы в колбу или банку составляет 5... 10 % массы засеваемой среды.
Размножение посевного материала. Пшеничные отруби подготавливают так же, как и при получении чистой культуры. Отруби, стерилизованные в бюксах под избыточным давлением 0,15 МПа в течение 1,5 ч, переносят на специальный стол, предварительно вымытый и продезинфицированный. Охлаждение отрубей до температуры 40...42 "С проводят перемешиванием с соблюдением правил микробиологической чистоты. Затем их засевают спорами из колбы с чистой культурой (расход культуры 0,5... 1 % массы отрубей). Суспензию спор готовят из расчета 1 часть культуры на 5 частей воды. Засеянные отруби распределяют по кюветам, предварительно простерилизованным при 120 °С в течение 2 ч, слоем толщиной 2...2,5 см. Кюветы сверху закрывают крышкой, под которую подкладывают стерильную бумагу; на отверстие по центру крышки накладывают стерилизованную ватную подушку, после чего кюветы помещают в термостат на 18...24 ч при температуре 30...32 "С. В дальнейшем кюветы устанавливают на стеллажи, расположенные в самой растильной камере, и ведут выращивание при 24...28 °С в течение 3...4 сут. Выросшую культуру затем помещают в комнату подсушки и в воздушно-сухом состоянии передают на хранение в кладовую, температура в которой не должна превышать +8 °С, но не ниже 0°С.
Посевной материал, приготовленный описанным способом, представляет собой спороносную культуру гриба, содержащую не менее 0,7 млрд спор в 1 г, из которых 86...90 % способны к прорастанию. Посевной материал не должен содержать посторонней микрофлоры, подвергаться замораживанию, оттаиванию и увлажнению.
Производственное культивирование. Технологическая схема получения культуры микроскопических грибов в растильных камерах на кюветах приведена на рис, 4. Этот способ связан с большими затратами ручного труда, однако пока применяется на заводах.
Пшеничные отруби ковшовым элеватором / подают на автоматические весы 2,
отсюда они поступают в загрузочный бункер 3,
а из него — в стерилизатор 5.
В этом аппарате отруби увлажняются водой, подаваемой через форсунки из бака 4
для стерильной воды. На 1 кг отрубей добавляют 0,2 л воды и 7...8 мл соляной кислоты относительной плотностью 1,19 или 2,7...3,0 мл серной кислоты плотностью 1,84. Отруби стерилизуют острым паром при температуре 103... 105 "С (давление 0,07 МПа) в течение 1...1,5 ч, периодически включая мешалку. По окончании стерилизации влажность отрубей доводят до 58...60 %, охлаждают их до 40...42 °С и затем вносят культуру гриба, полученную в отделении чистых культур.
Расход посевного подсушенного материала составляет 0,5...0,6 % массы отрубей. Посевной материал вводят в виде водной суспензии, приготовляемой из культуры гриба в 5... 10-кратном количестве кипяченой воды.
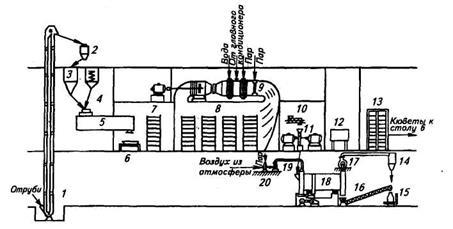
Рис. 4. Технологическая схема получения культуры микроскопических грибов в камерах на кюветах
Засеянную среду тщательно перемешивают в течение 15...20 мин, выгружают на раскладочный стол 6
и распределяют по кюветам. Наполненные кюветы ставят в этажерки 7, перевозят их на тележках в растильную камеру 8,
в которой поддерживают температуру 30...32 °С в течение 12... 16 ч (до начала интенсивного роста). В дальнейшем из кондиционера 9
в камеру подают вентилятором кондиционированный воздух температурой 26...28 °С и относительной влажностью 96...100 %. К концу выращивания культуры температуру снижают до 24...26 °С. Общая продолжительность выращивания 36...42 ч. В тех случаях, когда использование готовой культуры для осахаривания разваренной массы задерживается более чем на 2 ч, ее высушивают до влажности 18...20 %.
При необходимости транспортировки на дальние расстояния культуру гриба дробят на шнеке-дробилке 10
и дезинтеграторе 11
до получения частиц размером не более 10 мм и высушивают в сушилке 18
теплым воздухом до содержания влаги не более 10... 12 %.
Воздух нагревают в калорифере 20
и подают в сушилку вентилятором 19.
Отработавший воздух удаляют вентилятором 17
и выбрасывают в атмосферу через циклон 14.
Высушенную культуру шнеком 16
подают на весы 75 для фасования. Сюда же подаются уловленные в циклоне частички культуры, унесенные воздухом. Освободившиеся кюветы проходят через мойку 12,
обеспложиваются в стерилизаторе 13
и возвращаются под загрузку.
Готовую культуру гриба упаковывают в бумажные крафт-мешки. Каждый мешок снабжают этикеткой или биркой с указанием завода-изготовителя, наименования продукта, даты выпуска, номера партии, активности ферментов и массы. Хранят культуру в сухом помещении на деревянных стеллажах.
Предложены технологические схемы производства поверхностных культур плесневых грибов с использованием механизированных растильных установок: с вертикальными каналами (установка была смонтирована на Мичуринском экспериментальном заводе и может быть использована на предприятиях средней мощности — до 2 т культуры гриба в сутки); непрерывнодейст-вующей конвейерно-пластинчатой, предназначенной для специализированных ферментных предприятий мощностью более 2 т культуры в сутки.
4.3.2. Глубинное культивирование микроорганизмов
При глубинном культивировании микроорганизмы развиваются во всем объеме жидкой питательной среды. Так как подавляющее большинство продуцентов ферментов — строгие аэробы, среду интенсивно аэрируют. В микроорганизмах протекают два связанных процесса — синтез биомассы и синтез ферментов.
Питательные среды. Для глубинного культивирования используют жидкие среды, содержащие твердые компоненты. При работе с комплексными средами, основанными на естественном сырье с добавлением отрубей, ростков, кукурузного жмыха, глю-тена, свекловичного жома, спиртовой барды, следят за тем, чтобы не было крупных комочков, так как они затрудняют стерилизацию и могут привести к закупорке коммуникаций. Поэтому перед смешением твердых компонентов необходимо проводить их просеивание или грубую фильтрацию (например, зерно-картофельной барды).
Жидкую часть питательной среды (воду или фильтрат барды) обогащают питательными солями, гидролизатами белков, аминокислотами, источниками витаминов, различными углеводами. Содержание сухих веществ в жидких средах может колебаться от 1,5 до 20 % в зависимости от продуцента и принятого режима культивирования.
В научно-исследовательских организациях постоянно проводится работа по улучшению эффективности производства ферментных препаратов, в основном по повышению активности, зрелой культуры. Это достигается не только селекцией штаммов микроорганизмов, но и совершенствованием условий культивирования, в том числе и изменением состава питательной среды, поэтому приведенные среды могут подвергаться значительным изменениям при корректировании аэрации и других условий культивирования.
Получение и размножение посевного материала. Для засева производственной питательной среды при глубинном культивировании посевной материал готовят глубинным или поверхностным способом. Вид посевного материала зависит от продуцента:
для грибов это вегетативная мицелиальная масса или споронося-щая поверхностная, для бактерий — молодая растущая культура на начальной стадии спорообразования.
Посевной материал получают постадийным увеличением массы культуры продуцента. При небольшой производительности цеха это сводится к одной или двум операциям, а для заводов большой производительности представляет собой многоступенчатый процесс.
В связи с вводом штаммов микроскопического гриба Asp. awamori 466 или ВУД Т-2 усовершенствована и значительно упрощена схема приготовления спорового взамен вегетативного посевного материала. В простейшем виде она выглядит следующим образом. Пробирку с исходной культурой на агаризованной питательной среде пересевают водной суспензией в колбы со стерильными отрубями, которые выдерживают в термостате до полного созревания культуры. Из колб посевной материал в виде водной суспензии спор пересевается непосредственно в производственный ферментер, минуя инокулятор. В зависимости от объема ферментера количество посевных колб увеличивают в 2..4 раза. Спорового посевного материала вполне хватает для того, чтобы вырастить достаточно активную культуру при обычной продолжительности процесса. При этом получаются большой выигрыш в трудоемкости операций и более надежная защита от инфекции, так как в 2 раза сокращаются число операций и продолжительность выращивания посевного материала.
Культивирование на всех стадиях должно проводиться при оптимальной температуре, аэрации и строго определенное время. Если возникают непредвиденные задержки в использовании, то посевной материал охлаждают до 8... 10 °С и хранят не дольше 4 ч, иначе качество его может резко ухудшиться.
Посевной материал как на отдельных стадиях, так и готовый подвергают тщательному микробиологическому контролю. Он не должен быть инфицирован посторонней микрофлорой, в нем должно содержаться определенное количество спор или клеток на единицу массы, генетически заложенные в нем свойства продуцировать ферменты должны стойко сохраняться.
Производственное культивирование. Известны следующие способы глубинного культивирования: периодический, непрерывно-циклический и непрерывно-проточный.
Периодический способ
характеризуется несменяемостью питательной среды в ферментере, состав которой в процессе развития постепенно изменяется. Процесс протекает по-стадийно: в ферментер набирают питательную среду и задают посевной материал; после размножения микроорганизмов и накопления продуктов их обмена зрелую культуру выгружают, а все оборудование, коммуникации и запорные устройства промывают, а затем стерилизуют паром.
При непрерывноциклическом способе
микроорганизмы, расположенные на неподвижной насадке в ферментере, омываются средой, протекающей в замкнутом контуре, до полного потребления ими питательных веществ. После этого зрелую культуру выгружают, начиная с последнего, аппараты промывают, стерилизуют и цикл повторяют с последнего головного ферментера. Богатая питательными веществами среда в ходе такой циклической ферментации постепенно истощается; по времени пребывания среды в зоне реакции этот процесс более продолжителен, чем периодический.
Непрерывно-проточный
способ культивирования микроорганизмов более совершенен. Суть его заключается в том, что микробная популяция развивается в проточной питательной среде. Способ имеет две разновидности: гомогенно-непрерывный и градиентно-непрерывный. В первом случае выращивание ведут в одном ферментере; при тщательном перемешивании среды и аэрации обеспечивается одинаковое состояние культуры во всем объеме жидкости. В ферментер при этом непрерывно поступает свежая среда, а из него также непрерывно вытекает избыток зрелой культуральной жидкости.
Градиентно-непрерывное культивирование осуществляют в батарее ферментеров, соединенных переточными трубами. Засеянная среда с большим содержанием углеводов и других ингредиентов непрерывно перетекает из одного ферментера в другой и также непрерывно вытекает в виде готовой культуры.
Непрерывным культивированием в проточных средах можно выращивать микроорганизмы в условиях, оптимальных для их стадии развития. При этом такие важные факторы, как концентрация питательных веществ, количество продуктов обмена, рН, содержание растворенного кислорода, резко изменяющиеся при периодическом способе культивирования, поддерживаются постоянными на заданном уровне или изменяются по разработанной программе.
Способы непрерывного культивирования в производстве ферментных препаратов для спиртового производства еще находятся в стадии производственных испытаний. Поэтому, к сожалению, в настоящее время на заводах до сих пор применяют периодическое культивирование микроорганизмов.
4.4. концентрирование ферментных растворов
Глубинные культуры микроскопических грибов в жидком виде не могут сохраняться длительный срок без потери активности, перевозить их на большие расстояния вследствие значительного содержания в них воды (до 85...90 %) также экономически не всегда оправдано. Поэтому при организации снабжения культурой спиртовых заводов целесообразно ее предварительно концентрировать.
Концентрирование культур можно осуществлять на вакуум-выпарных установках с получением сиропов при температурах, обеспечивающих сохранение ферментов, или на распылительных сушилках с получением порошкообразного ферментного препарата. Однако эти способы сопровождаются значительными потерями активности (до 50 %) и поэтому не нашли распространения на спиртовых заводах. Начиная с семидесятых годов во ВНИИПрБТ был разработан и внедрен на Мичуринском экспе-риментальном заводе способ концентрирования глубинных культур ультрафильтрацией.
Способ основан на проведении процесса фильтрации через ряд полупроницаемых мембран, в результате чего ферменты и другие высокомолекулярные вещества задерживаются или выводятся из аппарата в
виде ультраконцентрата (концентрированного сиропа). Вода и часть низкомолекулярных веществ (пермеат) проходят через мембраны и также удаляются.
Потери активности при ультрафильтрации составляют до 15 % в пересчете на первоначальную активность исходной осветленной культуральной жидкости.
При использовании концентрированных препаратов на станции осахаривания применяют обычное оборудование для дозирования ферментных осахаривающих материалов.
4.5. Оборудование для получения амилолитических ферментов

МОДУЛЬНОЕ БИОТЕХНОЛОГИЧЕСКОЕ ОБОРУДОВАНИЕ СЕРИИ "ОКА-01"
4.5.1Назначение
"ОКА-01" представляет собой гибкое модульное биотехнологическое оборудование, позволяющее производить в непрерывном режиме широкий спектр биотехнологических продуктов, причём переход с выпуска одного продукта к другому осуществляется по программе под управлением компьютера.
Модульное построение биотехнологических агрегатов и систем управления, а также их новый принцип действия, позволяют не только быстро менять рецептуру производимых продуктов, но и обеспечивают масштабирование создаваемых производств от опытно-экспериментального до промышленного уровня.
4.5.2.Области применения
Предлагаемый комплект биотехнологического оборудования был использован для разработки новейших высокопродуктивных непрерывных технологий производства биопродуктов (Рис. 1):

4.5.3.Устройство
Состав базового комплекта модульного биотехнологического оборудования серии "Ока-01" представлен на Рис. 2:

4.5.3.1Модуль для приготовления питательной среды (МПС)
Модуль предназначен для приготовления питательных сред из растительного сырья с последующим разделением полученной суспензии на жидкую и твёрдую фракции и представляет собой набор унифицированного оборудования для автоматизированного непрерывного производства питательных сред с заданными параметрами по концентрации, соотношению компонентов, pН и температуре. В состав "МПС", входят: бункер для исходного сырья (зерно, отруби и др.), обеспечивающий запас исходного сырья для непрерывного производства питательной среды; измельчитель твёрдых веществ с дозирующим устройством, обеспечивающий дробление и транспортировку расчётной дозы сырья в смеситель по заданной программе;
парогенератор для получения насыщенного водяного пара, используемого для варки растительного сырья, термостатирования и разбавления приготовляемой среды; смеситель с термостатируемой рубашкой, обепечивающий гомогенное перемешивание и дозированный отбор сгущенной среды по заданному алгоритму; устройство для фракционного разделения питательной среды, обеспечивающее выделение из среды жидкой и твёрдой фракций.
4.5.3.2.Модуль стерилизационный (МС)
Модуль предназначен для стерилизации насыщенным водяным паром ферментационного оборудования, питательных сред и титранта, воздуха и отходящих газов, а также для обеспечения асептических условий работы всех устройств базового модуля.
В состав "МС" входят: парогенератор, обеспечивающий насыщенным водяным паром работу насосов, дозаторов, паростерилизуемых разъёмов трубопроводов, стерилизаторов питательных сред, автоклава и термостатов;
автоклав, обеспечивающий ускоренную стерилизацию лабораторной посуды и титранта в условиях циклической принудительной конденсации пара;
стерилизатор питательной среды, обеспечивающий дозированную стерилизацию сгущенных питательных сред с их последующим разбавлением стерильной дистиллированной водой до заданных концентраций; устройство для слива стерильной среды, обеспечивающее охлаждение и дробную подачу приготовленной питательной среды в ферментёр по заданному алгоритму.
4.5.3.3.Модуль ферментационный (МФ)
Модуль предназначен для выращивания гетеротрофных микроорганизмов в периодическом или проточном режимах культивирования, проведения процессов биосинтеза и биокатализа веществ.
В состав "МФ" входят: ферментёр для проведения микробиологических процессов в асептических условиях: снабжён турбинной мешалкой, обеспечивающей гомогенное перемешивание культивационной среды с фиксированным пенообразованием и щадящим воздействием на биомассу и твёрдые включения, например, носители уплотнённой биомассы или сорбционные материалы; работает при коэффициенте загрузки 0,8-0,9, что позволяет увеличить выход продукта; парогенератор, обеспечивающий насыщенным водяным паром стерилизацию ферментационного оборудования, работу паростерилизуемых разъёмов, дозаторов и термостата;
ёмкость для осаждения биомассы, обеспечивающая накопление и охлаждение культуральной жидкости, вытекающей из ферментёра, в результате чего происходит её расслоение с образованием осадка;
термостат, обеспечивающий поддержание заданных температур в ферментёре и ёмкости для осаждения биомассы; дозаторы для слива продукта, обеспечивающие раздельный слив осаждённой биомассы и надосадочной культуральной жидкости в асептических условиях по заданным алгоритмам.
4.5.3.4.Модуль интерактивного управления (МУ)
Модуль предназначен для автоматического управления всеми устройствами и агрегатами, а также для измерения и регистрации текущих параметров процесса по pН, температуре, растворенному кислороду и оборотам мешалки.
В состав "МУ" входят: блоки питания, обеспечивающие стабилизированное питание электронных приборов; контроллер, обеспечивающий прохождение и обработку аналоговых и дискретных сигналов, ввод и обработку сигналов с датчиков, вычисление и выдачу на исполнительные устройства значения регулирующих воздействий: формирует программно-временную последовательность команд для управления клапанами и осуществляет обмен информацией с пультом оперативного управления и компьютером верхнего уровня; компьютер, обеспечивающий интерактивное программное управление многостадийными процессами.
Ряд агрегатов и приборов, входящих в состав модульного биотехнологического оборудования серии "Ока-01", могут использоваться самостоятельно и иметь широкое применение. К ним относятся:
Парогенератор непрерывного действия:
Объём испарителя от 5 до 30 л
Температура водяного пара от 105 до 130°С
Производительность от 3 до 10 кг/час.
|
Автоклав:
Объём камеры 20 л
Температура пара от 105 до 120°С.
|
Паростерилизуемый разъём:
Время стерилизации стыков 15 мин
Проходное сечение трубопроводов du=4-8 мм
Давление от 0,02 до 0,2 МПа.
|
Прибор интерактивного управления многостадийным процессом:
IBM совместимый компьютер
Контроль и регистрация текущих параметров
Графическое отображение процессов.
|
В зависимости от задачи эксперимента или видов исходного сырья и конечного продукта, который необходимо получить в результате биотехнологического процесса, базовый комплект трансформируется и дополняется специфическими техническими средствами.
4.5.3.5.Выполняемые функции
Базовый комплект позволяет обеспечить:
· комплексное технологическое оснащение целевого процесса;
· сокращение времени стерилизации и сохранение качества среды за счёт нового метода стерилизации;
· дозирование жидкостей и доставку стерильных сред в асептических условиях по заданному алгоритму;
· недопущение накипи в стерилизационной камере;
· возможность реализации периодических, объёмно-доливных и непрерывных процессов биосинтеза, биокатализа и биотрансформации веществ в асептических условиях;
· увеличение загрузки ферментёров до 80% без использования пеногасителей;
· разделение осаженной и охлаждённой биомассы от нативной жидкости в процессе ферментации с возможностью дозированного слива или рециркуляции внутри системы в асептических условиях;
· возможность использования блоков оборудования в качестве биотехнологического конструктора для формирования различных многостадийных процессов с единым интерактивным управлением;
· повышенный выход продуктов в 1,5-2 раза при значительной экономии расходуемого сырья.
4.5.3.6.Примеры использования ОКА-01
С применением модульного биотехнологического оборудования реализованы следующие процессы:
1. Производство мицелиальной биомассы грибов
.
Биомасса используется в качестве инокулюма для выращивания плодовых тел вешенки, опёнка и других высших грибов. Предлагаемая технология позволяет значительно снизить себестоимость процесса выращивания высших грибов за счёт исключения импорта и создания собственного высокопродуктивного производства инокулюма.
2. Производство пекарских дрожжей из крахмалсодержащего сырья.
Внедрение новой технологии на хлебозаводах позволяет использовать отходы производства мучных и крупяных продуктов для производства пекарских дрожжей, что снижает себестоимость целевых продуктов, транспортные расходы и позволяет решить экологические проблемы.
3. Производство растворов амилолитических ферментов
.
Внедрение новой технологии на спиртовых заводах позволит решить важнейшую проблему утилизации барды и исключить импорт амилаз, расходуемых для ферментативного гидролиза зерна. Возможна реализация новой технологии для промышленного непрерывного производства амилолитических ферментов в России.
4. Производство осахаренного сиропа (подсластителей) из зерновых.
Использование подсластителей в пищевой промышленности значительно снизит себестоимость выпускаемых продуктов, в рецептуру которых входит импортируемый сахар. Возможно использование новой технологии для промышленного производства глюкозы в России. Сопутствующим продуктом при получении подсластителей является белково-витаминная кормовая добавка, содержащая все незаменимые аминокислоты, витамины, белки и углеводы.
5. Средства защиты раст
ений.
Предлагается ферментационный модуль и технологии для высокопродуктивного производства субстанций биопрепаратов для защиты растений. Субстанции биопрепаратов обладают высокой активностью, не загрязняют окружающую среду и могут производиться в хозяйствах в необходимом ассортименте, приуроченном к срокам обработки растений.
6. Производство пищевых добавок на основе мицелиальных грибов
.
Пищевая добавка - экологически чистый белковый, витаминный продукт, представляющий собой мицелий высших грибов в виде монокультуры, выращенной в асептических условиях. Пищевой белок предназначен для использования в качестве вкусовой добавки в пищевые продукты - майонез, пищевые концентраты, соусы, мясной фарш и др.
7. Производство новых целебных
напитков.
Все напитки, полученные на оборудовании серии “Ока-1”, - экологически чистые белковые, витаминные препараты в виде суспензии винных дрожжей, выращенных на пивном сусле в асептических условиях.
8. Производство лечебно-профилактических препаратов
с использованием дрожжей Saccharomyces vini Т-8
, обладающих ярко выраженным эффектом нейтрализации токсичных метаболитов возбудителей желудочно-кишечных заболеваний и неспецифическим иммунностимулирующим действием. Новый препарат в отличие от имеющихся на рынке аналогов может производиться по замкнутой безотходной автоматизированной технологии в асептических условиях непрерывного процесса.
ЛИТЕРАТУРА
1. Большая медицинская энциклопедия. Гл.ред. Б.В.Петровский. – 3-е изд. – М.: Советская энциклопедия. – Т. 5, 1986, 544 с.
2. Большая медицинская энциклопедия. Гл.ред. Б.В.Петровский. – 3-е изд. – М.: Советская энциклопедия. – Т. 26, 1986, 568 с.
3. Большая медицинская энциклопедия. Гл.ред. Б.В.Петровский. – 3-е изд. – М.: Советская энциклопедия. – Т. 28, 1986, 543 с.
4. Диксон, Уэбб. Ферменты. Ред. М.Г.Дунина. – М.: 726 с., 1961
5. Биологическая химия Д.Г.Кнорре, С.Д.Мызина. – 3-е изд. – М.: Высш.шк. 2002. – 479 с.: ил.
6.Попов Е.М. Структурно-функциональная организация белков. – М.: Наука, 1992.–358 с.
7.Галич И.П. Амилазы микроорганизмов. – Киев: Наук. думка, 1987. – 192 с.
8.Жеребцов Н.А. Амилолитические ферменты в пищевой промышленности. – М.:Лег. и пищ. пром-сть, 1984. – 160 с.
9.Кочетов Г.А. Практическое руководство по энзимологии. – М.:Высш. шк., 1980. – 272 с.
10.Ковалева Т.А., Селеменев В.Ф., Гречкина В.Р. Некоторые свойства глюкоамилазы // Труды III Всесоюзной межуниверситетской конференции по физико-химической биологии. – Тбилиси, 1982. – С. 240-241.
11.Ковалева Т.А., Башарина О.В., Селеменев В.Ф. Исследование структуры глюкоамилазы методом ИК спектроскопии // Тезисы докладов VII Всесоюзной конференции по спектроскопии биополимеров. – Харьков, 1991. – С. 134-135.
12.Ковалева Т.А. Кинетико-термодинамические аспекты катализа полисахаридов свободными и иммобилизованными амилазами // Биофизика. - Т.45, N3, - 2000 - C. 439-444.
|