| Федеральное агентство по образованию
Пензенский государственный педагогический университет
им. В.Г. Белинского
Факультет
Естественно-географический
Кафедра
Биохимии
Дипломная работа
УРОВЕНЬ ВЕЩЕСТВА Р И активность ФЕРМЕНТОВ ОБМЕНА РЕГУЛЯТОРНЫХ ПЕПТИДОВ В СЫВОРОТКЕ крови спортсменов ПРИ ФИЗИЧЕСКОЙ РАБОТЕ
Студент
________________________ Дашкевич Д.В.
Руководитель
_________________________ Соловьев В.Б.
К защите допустить.
Протокол № от «____» ___________200_г.
Зав. кафедрой
__________________________ Генгин М.Т.
Пенза, 2009г.
СОДЕРЖАНИЕ
Список сокращений
ВВЕДЕНИЕ
ГЛАВА 1. ОБЗОР ЛИТЕРАТУРЫ
1.1 Вещество Р
1.2 Ферменты обмена регуляторных пептидов
1.2.1 Карбоксипептидаза N
1.2.2 Пептидил-дипептидаза А
1.2.3 Лейцинаминопептидаза
1.3 Механизм адаптации к физической работе. Роль регуляторных пептидов
ГЛАВА 2. МАТЕРИАЛЫ И МЕТОДЫ
2.1 Материалы исследования
2.2 Методы исследования
2.2.1 Моделирование физической работы
2.2.2 Метод определения концентрации вещества P
2.2.3 Метод определения активности КПN
2.2.4. Метод определения активности ангиотензинпревращающего фермента
2.2.5 Метод определения активности лейцинаминопептидазы
ГЛАВА 3. РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
ВЫВОДЫ
СПИСОК ЛИТЕРАТУРЫ
Список сокращений
АД – артериальное давление
АКТГ – адренокортикотропный гормон
АПФ – ангиотензинпревращающий фермент
БСА – бычий сывороточный альбумин
ГЭМЯК – гуанидиноэтилмеркаптоянтарная кислота
кбз- – карбобензокси-
КПN – карбоксипептидаза N
мРНК – матричная рибонуклеиновая кислота
ПВДС (ДСИП) - пептид, вызывающий дельта сон (дельта-сон индуцирующий пептид
ТХУ – трихлоруксусная кислота
ФМСФ – фенилметилсульфонилфторид
ЭБС – эмоционально-болевой стресс
ЭДТА – этилендиаминтетрауксусная кислота
ВВЕДЕНИЕ
В последнее время в биохимии спорта огромное внимание уделяется поиску тех ключевых факторов регуляции метаболизма, воздействие на которые позволит значительно улучшить спортивный результат. Наиболее перспективной мишенью для воздействия является пептидергическая система. Нейропептиды играют важную роль в адаптационных процессах, участвуют в формировании пищедобывательного и полового поведения, регулируют состояние иммунной системы [2]. Многие из этих веществ вовлекаются в регуляцию полового созревания, влияют на половую дифференциацию организма, на процессы внимания и памяти, на эмоциональное поведение, обладают анальгезирующим действием. Данные исследований последних лет свидетельствуют об участии некоторых регуляторных пептидов в реакциях адаптации при физической работе. Однако, ограничения, связанные с трудностью введения регуляторных пептидов в организм животных, снижают возможности их использования для регуляции функционального состояния организма и улучшения спортивного результата. Кроме того, изучение содержания уровня того или иного нейропептида в тканях и сыворотке крови не дает достаточно точных представлений о динамических процессах, происходящих в пептидергической системе при физической работе. Более информативным является изучение процессов синтеза и трансформации нейропептидов, поскольку концентрация биологически активных пептидов зависит от активности ферментов, участвующих в их обмене.
Еще одним существенным недостатком имеющихся работ по изучению содержания регуляторных пептидов при физической работе является отсутствие дифференциации объектов исследования по уровню тренированности. Высокий спортивный результат могут показать небольшое количество специализированных людей-спортсменов высокой квалификации, а обычные люди повторить его не в состоянии. Биохимические изменения, которые его сопровождают, являются очень значительными и превосходят изменения, которые наблюдаются у обычных людей, не занимающихся спортом, как в количественном, так и в качественном отношениях.
Знание путей образования и инактивации регуляторных пептидов, участвующих в ответе организма на физическую нагрузку, позволит, избирательно воздействуя на ферменты их метаболизма, регулировать уровень биологических форм тех нейропептидов, от которых зависит спортивный результат.
Таким образом, целью нашей работы
являлось выяснение роли пептидергической системы в адаптации к физической работе у спортсменов высокой квалификации.
При этом ставились следующие задачи
:
1. Изучить уровень вещества Р, регуляторного пептида контролирующего многие адаптационные перестройки и обладающего широким спектром физиологического действия, в сыворотке крови спортсменов и не спортсменов в норме и при физической работе.
2. Изучить активность ферментов обмена регуляторных пептидов - пептидил-дипептидазы А, карбоксипептидазы N и лейцинаминопептидазы в сыворотке крови спортсменов и не спортсменов в норме и при физической работе.
Научная новизна и практическая ценность работы
. Впервые исследован уровень вещества Р и активность ферментов его обмена при физической работе. Полученные данные позволят приступить к разработке методов направленного влияния на активность ферментов обмена пептидов, участвующих в адаптации к физической работе с целью улучшения спортивного результата.
Вещество P (Substance P) - один из наиболее известных пептидов, открытый в 1931 году [42].
Вещество Р - биологически активный пептид из 11 аминокислотных остатков. Вещество Р обладает широким спектром биологической активности: оказывает сосудорасширяющее действие, способствует дегрануляции тучных клеток, является хемоаттрактантом для лейкоцитов, активирует синтез и высвобождение медиаторов воспаления. Показано, что снижение концентрации вещества Р в синовиальной жидкости уменьшает тяжесть экспериментального артрита.
Обладает широким спектром физиологической активности: изменяет артериальное давление крови, капиллярную проницаемость, сокращение гладкой мускулатуры, обладает секретогенным действием, высвобождает пролактин и пищеварительные гормоны. В последнее время интенсивно изучается роль вещества Р и его аналогов в регуляции центральных процессов - порога болевого воздействия, обучения, сна, устойчивости к стрессу. В головном мозге вещество Р участвует в процессах, связанных с функцией другого нейрорегулятора – допамина. При повреждении допаминэргических волокон выявлено снижение экспресии мРНК, кодирующих образование вещества Р, энкефалинов, динорфина. На базе основной структуры вещества Р получено большое число химических производных, которые обладают свойствами сионистов или антагонистов тахикининовых рецепторов [57].
Вещество Р присутствует во многих специфических нейронных путях в головном мозгу, а также в первичных сенсорных волокнах периферических нервов. Некоторые из этих сенсорных нейронов, тела которых лежат в сенсорных ганглиях по обе стороны спинного мозга, содержат вещество Р и выделяют его из своих аксонных окончаний в синапсах со спинальными нейронами. Поскольку вещество Р возбуждает те спинальные нейроны, которые легче всего реагируют на болевые стимулы, было высказано предположение, что оно служит сенсорным медиатором, специфически связанным с передачей информации о боли от периферических болевых рецепторов в центральную нервную систему.
Морфиноподобный пептид энкефалин тоже в изобилии содержится в мелких нейронах в той части спинного мозга, куда приходят волокна, содержащие вещество Р. Энкефалин и препараты опия способны подавлять выделение вещества Р из сенсорных волокон. Поэтому нейроны, содержащие энкефалин, могут регулировать поступление болевых сигналов в головной мозг, модулируя выделение вещества Р на уровне первого переключения в центральной нервной системе. Подобные же тормозные взаимодействия возможны и на более высоких уровнях мозга. Вещество Р - не единственный предполагаемый медиатор, локализующийся, как показано, в сенсорных нейронах; к таким же идентифицированным к настоящему времени веществам относятся ангиотензин, холецистокинин, соматостатин и глутаминовая кислота. Таким образом, по мере того как все больше становится известно о сенсорных медиаторах и их модуляционных механизмах в спинном мозгу, начинает возникать картина поразительной химической сложности.
Гипотетический воротный механизм в первом синаптическом переключении, возможно, регулирует передачу информации о боли от периферических болевых рецепторов к головному мозгу. В задних рогах спинного мозга вставочные нейроны, содержащие пептидный медиатор энкефалин, образуют синапсы на аксонных окончаниях болевых нейронов, которые в качестве медиатора используют вещество Р. Выделяемый вставочными нейронами энкефалин тормозит выход этого вещества, из-за чего воспринимающий нейрон в спинном мозгу получает меньше возбуждающей стимуляции и поэтому посылает в головной мозг меньше связанных с болью импульсов. Такие опийные препараты, как морфий, по-видимому, связываются с незанятыми рецепторами энкефалина, имитируя подавление боли, производимое энкефалиновой системой.[62]
По многим функциональным признакам вещество Р следует отнести к группе тахикининов: сходный спектр физиологических функций, общая система рецепторов, родственные признаки структуры предшественников. Однако тщательное исследование функционального профиля вещества Р и успехи в множественном синтезе его аналогов позволяют условно определить его как отдельную группу [59,44,68].
В группу тахикининов (Tachykinins) входят пептиды, имеющие, как и вещество P, сходную С-концевую последовательность [-(Phe)-Gly-Leu-Met]. Сюда относятся соединения, обозначенные как нейрокинины, нейромедины, бомбезины, а также оригинальные тахикинины, обнаруживаемые у амфибий, рыб и беспозвоночных животных. Другое сходство относится к наличию общего предшественника - бета-препротахикинина.
Распределение тахикининов в тканях одного и того же организма (в первую очередь, высших позвоночных) весьма разнообразно. Поэтому столь же широк спектр их физиологической активности: сокращение гладкой мускулатуры кишечника, бронхов, зрачка глаза, медиация центральных (поведенческих) и гормональных процессов, высвобождение других физиологически активных веществ, периферическая ноцицепция.
Функциональное разнообразие тахикининов связано с их взаимодействием с другими физиологически активными веществами, как пептидной (бета-эндорфин, нейропептид Y, ген-кальцитониновый пептид), так и непептидной природы - допамином, интерлейкинами и др. Существенным в рассмотрении этой группы пептидов оказалось выявление сложной системы тахикининовых - нейрокининовых рецепторов (NK1, NK2, NK3) и их подтипов.
Современная фармакология тахикининов и вещества Р в значительной мере сосредоточена на исследовании тканевой и видовой специфичности этих рецепторов, а также поиске разнообразных антагонистов, нивелирующих эффекты тахикининов.[63,69].
1.2 Ферменты обмена регуляторных пептидов
Нейропептиды – это полифункциональные высокоактивные вещества пептидной природы, которые играют важную роль в реализации и регуляции различных физиологических и поведенческих реакций организма.
Нейропептиды играют важную роль в адаптационных процессах, проявляют анальгетические эффекты, участвуют в формировании пищедобывательного и полового поведения [20,22,24,28]. Многие из этих веществ вовлекаются в регуляцию полового созревания [2,5,29,33]. Нейропептиды влияют на половую дифференциацию организма (гонадотропин-рилизинг-фактор, ЛГ, ФСГ, пролактин [19,21,34], на процессы внимания и памяти (например, АКТГ и α-меланотропин – стимуляторы запоминания и внимания), на эмоциональное поведение (тиролиберин, меланостатин, кортиколиберин – стимуляторы эмоционального поведения), обладают анальгезирующим действием (нейротензин, опиоидные пептиды).
Уровень нейропептидов определяется соотношением скоростей их синтеза и деградации.
Нейропептиды синтезируются в организме на рибосомах гранулярного эндоплазматического ретикулума в виде высокомолекулярных неактивных предшественников (препропептидов) [11,17]. В состав последних могут входить аминокислотная последовательность как одного, так и нескольких нейропептидов. Известно много белков, содержащих в своей структуре последовательности нейропептидов: предшественник гонадотропин-рилизинг-фактора, проопиомеланокортин, препроэнкефалин А, продинорфин (препроэнкефалин В) и другие [26,1].
Все препропептиды содержат на N-конце сигнальную последовательность из 15-20 остатков гидрофобных аминокислот. Нейропептиды, входящие в состав предшественника, как правило, ограничены с C- и N-концов парами остатков основных аминокислот – аргинина и лизина.
Сигнальная последовательность препропептидов необходима для взаимодействия с рецепторами эндоплазматического ретикулума и переноса предшественника нейропептида в просвет ретикулума. В цистернах эндоплазматического ретикулума под действием сигнальной эндопептидазы происходит отщепление сигнальной последовательности, а также N-гликозилирование и формирование характерной для полипептида третичной структуры, которая препятствует обратному выходу белка в цитоплазму. Посттрансляционная модификация, включающая гликозилирование, амидирование, ацетилирование или сульфирование, предотвращает нарушение процессинга и образование нетипичных пептидов.
Для получения активных форм, полипептиды подвергаются посттрансляционному процессингу, одним из основных механизмов которого является ограниченный протеолиз [3,6,14].
Процессинг биологически активных пептидов [6,7] осуществляется при передвижении молекул пропептидов по гранулярному эндоплазматическому ретикулуму, комплексу Гольджи и в секреторных везикулах [55]. Секреторные везикулы содержат полный набор ферментов, необходимых для процессинга и специальные системы поддержания pH внутри везикул [15].
Процессинг нейропептидов [64] внутри секреторных везикул включает в себя эндо- и экзопротеолитические реакции. Эндопротеолиз осуществляется при действии трипсиноподобных протеиназ (проопиомеланокортин-превращающего фермента [65,66], продинорфин-превращающего фермента [47], тиоловой прогормонконвертазы [39,40], субтилизиновых эндопептидаз семейства фурина, PC1, PC2, PC3 и PC4 [73]. В результате происходит расщепление пропептидов по парам остатков основных аминокислот [15,77].
Продукт, образовавшийся после действия эндопептидаз, далее подвергается экзопротеолизу с участием аминопептидазо-В- и/или карбоксипептидазо-В-подобных ферментов [7,9]. В результате происходит удаление ”лишних” N- и/или С-концевых остатков основных аминокислот.
Известно, что в различных тканях из одного белкового предшественника образуются различные нейропептиды. Так из проопиомеланокортина в аденогипофизе образуются преимущественно АКТГ, β-липотропин и β-эндорфин. В промежуточной доле гипофиза они подвергаются дальнейшему расщеплению с образованием α-меланоцитстимулирующего гормона и фрагментов β-эндорфина [3]. Тканевая специфичность, по-видимому, может быть связана с различным набором ферментов в разных тканях и/или с различными способами регуляции их активности. Поэтому представляет интерес изучение ферментов процессинга со сходной (но не идентичной) субстратной специфичностью. Такие исследования интересны не только для выяснения вопросов, связанных с функционированием данных ферментов, но и для понимания механизмов образования различных нейропептидов из одних и тех же предшественников в разных тканях.
Поскольку карбоксипептидазо-В-подобные ферменты, то есть отщепляющие остатки основных аминокислот (аргинина и лизина) с карбоксильного конца пептидов, участвуют в конечной стадии процессинга биологически активных пептидов, то их изучение представляет особый интерес. Также эти ферменты участвуют в инактивации нейропептидов. В сыворотке крови такими ферментами являются карбоксипептидаза N (КПN) и ангиотензин-превращающий фермент (АПФ).
1.2.1 Карбоксипептидаза
N
Карбоксипептидаза N (КПN, энкефалинконвертаза, карбоксипептидаза Е, КФ 3.4.17.10) выделена и охарактеризована из мозга, гипофиза, хромаффинных гранул надпочечников, эндокринных клеток поджелудочной железы. Во всех органах и тканях КПN представлена двумя формами: растворимой и мембраносвязанной. Обе формы являются гликопротеинами, имеют идентичную субстратную специфичность и чувствительность к групп-специфичным реагентам. Молекулярная масса мембранной формы 55-57 кДа, растворимой формы – 53-57 кДа [45,52,53].
Карбоксипептидаза N является тиолзависимым металлоферментом, в активном центре которого находится Zn2+
.[54] Фермент имеет оптимум рН 5,5-6,0, активируется ионами Co2+
в 5-10 раз, ионами Ni2+
в 2-3 раза, ингибируется CuCl2
, HgCl2
, аминопропилмеркаптоянтарной кислотой, 2-меркаптометил-3-гуанидилэтилтиопропановой кислотой, ЭДТА и 1,10-фенантролином. Сульфат цинка, хлорид кальция, N-этилмалеимид и ФМСФ не влияют на активность КПN [78]. Наиболее эффективными ингибиторами являются ГЭМЯК и гуанидинопропилянтарная кислота с Ki
8,8 нМ и 7,5 нМ, соответственно. КПN ингибируется Met- и Leu-энкефалинами, веществом Р, вазопрессином, окситоцином, тиреотропин-рилизинг-фактором [59].
Уровни мРНК КПN и активности КПN в мозге и тканях в целом коррелируют между собой. Наивысшие уровни мРНК КПN обнаружены в пирамидальных клетках гиппокампа, в передней и промежуточной долях гипофиза, эпендимных клетках боковых желудочков мозга, базолатеральной миндалине, супраоптическом и паравентрикулярном ядрах. Средние уровни мРНК КПN у крыс обнаружены в таламусе, медиальном коленчатом ядре, коре мозжечка и промежуточной оливе. Наименьшие уровни выявлены в гранулярном клеточном слое гиппокампа, латеральном гипоталамусе, бледном шаре и в ретикулярной формации ножки мозга.
В клетке КПN локализована, в основном, в секреторных гранулах, причем часто совместно с биологически активными пептидами – инсулином [45], энкефалинами [58], вазопрессином [71], окситоцином [70], веществом P [44], атриальным натрийуретическим фактором.[67].
Она обладает, практически, абсолютной специфичностью по отношению к пептидным субстратам с С-концевыми остатками основных аминокислот. КПN хорошо отщепляет остатки лизина и аргинина от Arg8
-вазопрессин-Gly-Lys-Arg [71], а также превращает 125
I-Met-энкефалин-Arg6
и 125
I-Met-энкефалин-Lys6
в 125
I-Met-энкефалин. Фермент отщепляет остатки -Lys15
-Lys16
-Arg17
с карбоксильного конца фрагмента АКТГ (АКТГ1-14
) и остатки аргинина с С-конца гиппурил-L-Arg и Leu-энкефалин-Arg [58]. КПN с очень низким сродством отщепляет остаток гистидина с карбоксильного конца проокситоцина [76].
Следует отметить, что уровень мРНК КПN способен быстро изменяться в ответ на внешние воздействия, вызывающие изменения уровня биологически активных пептидов или уровня их мРНК. Кроме того, в случае синтеза дефектной (неактивной КПN) (мутация в гене, кодирующем КПN) наблюдается нарушение синтеза и секреции многих биологически активных пептидов.
Физико-химические свойства, субстратная специфичность, тканевая, клеточная и субклеточная локализация, особенности изменения активности фермента при различных фармакологических воздействиях на организм и культуры клеток [13], нарушение синтеза нейропептидов у мышей с дефектной КПN свидетельствуют о том, что исследуемый фермент вовлекается в процессинг многих биологически активных пептидов, таких как энкефалины, АКТГ, b-эндорфин, вазопрессин, окситоцин, нейротензин, меланоцитстимулирующий горомон, вещество Р и др. [6], а также участвует в (алкоголизм, стресс, болезнь Альцгеймера, в общем, в адаптационных процессах. [31]
1.2.2 Пептидилдипептидаза А
Ангиотензинпревращающий фермент (АПФ, дипептидил-карбоксипептидаза A, кининаза II, карбоксикатепсин, пептидилдипептидаза А, КФ 3.4.15.1) обладает пептидилдипептидазной и слабыми трипептидилкарбоксипептидазной и эндопептидазной активностями [50]. Он выделен и очищен из разных тканей, в том числе из мозга, различных видов животных[46]. Фермент из всех тканей (эндотелиальная форма), за исключением семенников (тестикулярная форма), имеет очень близкие физико-химические и иммунологические свойства, тогда как фермент из семенников отличается от ангиотензинпревращающего фермента из других источников по молекулярной массе и иммунологическим свойствам.
В мозге обнаружена только эндотелиальная форма. Она состоит из одной полипептидной цепи, имеет молекулярную массу 170-180 кДа и содержит около 13% остатков нейтральных сахаров. Эндотелиальная форма состоит из двух гомологичных доменов, каждый из которых имеет активный центр и центр связывания Zn2+
[10].
Отличия в степени гликозилирования приводят к образованию двух иммунологически идентичных форм, одна из которых (эндотелиальная) с молекулярной массой 180 кДа присутствует, исключая семенники, во всех тканях, в том числе и мозге, а вторая (нейрональная) с молекулярной массой 170 кДа – обнаруживается только в мозге, и не присутствует в других тканях.[41]
Имеются единичные сообщения об обнаружении в различных органах и тканях более высокомолекулярных каталитически активных форм АПФ с молекулярной массой 600 кДа, 430 кДа, 330 кДа, 240 кДа, иммунологически полностью идентичных эндотелиальной форме. В настоящее время неясно, являются ли эти формы предшественниками АПФ, или образуются при гель-фильтрации благодаря склонности молекул фермента, как и других мембраносвязанных белков, к агрегации. pI фермента из различных органов и тканей колеблется в пределах от 4,6 до 5,1, что обусловливает дополнительную гетерогенность фермента при электрофоретическом разделении. Полагают, что гетерогенность АПФ может иметь важное значение для избирательности регуляции его активности в различных тканях. [25].
АПФ из различных источников имеет оптимум pH 7,6-8,2, для проявления максимальной активности необходимы ионы Cl-
.[30] АПФ содержит 1 прочно связанный ион Zn2+
на каждый активный центр и сильно ингибируется хелатирующими агентами, такими как ЭДТА и о-фенантролином [61]. Фермент ингибируется брадикининпотенциирующим фактором (Ki
40 нМ), дитиотреитолом, 2-меркаптоэтанолом и додецилсульфатом натрия. N-этилмалеимид, бацитрацин, пуромицин, фосфорамидон, тиорфан, ингибитор металл-зависимых основных карбоксипептидаз ГЭМЯК не влияют на его активность [10]. Создан целый класс пептидных аналогов субстратов АПФ с Ki
порядка 10-50 нМ, наиболее известные из них каптоприл (IC50
» 20 нМ), еналаприл (IC50
25-35 нМ), лизиноприл (IC50
3-10 нМ) [49].
Фермент in vitro катализирует расщепление в общей сложности более 30 биологически активных пептидов и их предшественников [75]. Он превращает ангиотензин I в ангиотензин II (Km 4-70 мкМ), последовательно отщепляет два дипептида с C-конца брадикинина, расщепляет неокиоторфин с образованием киоторфина (Km
0,58 мМ), Met-энкефалин-Arg6
-Phe7
с образованием Met-энкефалина (Km
0,30 мМ), вещество P и вещество K, холецистокинин и гастрин, энкефалин, нейротензин, нейрокинины A и B, рилизинг-фактор лютеинизирующего гормона. АПФ катализирует образование Met-энкефалин-Arg6
из Met-энкефалин-Arg6
-Gly7
-Leu8
, отщепляет последовательно два дипептида с C-конца динорфина A 1-8, расщепляет натрийуретический фактор из мозга и предсердий, вазопрессин, окситоцин.
Активность АПФ in vitro ингибируется многими биологически активными пептидами и их предшественниками: неокиоторфином, Met-энкефалин-Arg6
-Phe7
, b-липотропином, веществом P, брадикинином, Leu-энкефалин-Arg6
и некоторыми дипептидами [10].
АПФ широко распространён в тканях человека и животных. Наиболее высокий уровень АПФ в периферических тканях обнаружен в лёгких и семенниках, в почках активность АПФ примерно в 25 раз меньше, чем в лёгких. В сыворотке крови активность АПФ примерно в 200 раз ниже, чем в лёгких [18]. Активность фермента в мозге сопоставима с активностью в почках. Наиболее высокое содержание АПФ обнаружено в гипофизе и стриатонигральном тракте. АПФ локализован преимущественно в синаптосомах [43]. В стриатонигральном тракте фермент связан с фракцией мембран, содержащей мускариновые рецепторы, причём в этой фракции обнаруживается только нейрональная, но не эндотелиальная форма АПФ. Таким образом, на мембранах дендритов локализована только нейрональная форма АПФ.[41]
Хорошо известно, что АПФ является компонентом ренин-ангиотензиновой системы. В периферических органах и тканях он вовлекается в обмен ангиотензина и брадикинина и является важным звеном регуляции артериального давления, водно-солевого баланса и воспалительных процессов.[60] В последнее время обнаружено, что АПФ вовлекается в определение эмоционального статуса животных, устойчивости к эмоциональному стрессу [12], агрессивности [16], предрасположенности к потреблению этанола, мозговой АПФ вовлекается в формирование наследственно обусловленной гипертонии, в ответ на воздействие стрессирующих факторов [27] и потребление этанола [17]. Ингибиторы АПФ влияют на потребление воды и этанола, усиливают анальгетическую активность Met-энкефалин-Arg6
-Leu7
и подавляют его деградацию в мозге. Каптоприл, кроме того, обладает центральным действием, является антидепрессантом, его антидепрессивный эффект блокируется налоксоном, он пролонгирует и усиливает анальгетические эффекты Met- и Leu-энкефалинов, причём это усиление также блокируется налоксоном. Участие АПФ в некоторых из перечисленных выше физиологических и патологических процессов, а также многие из перечисленных эффектов, вызванных введением его ингибиторов, невозможно объяснить исходя из представления, что единственной функцией АПФ является участие в обмене ангиотензинов и брадикинина. Приведенные факты позволяют предположить, что АПФ вовлекается в обмен и других регуляторных пептидов [10], что подтверждается и достаточно высокой активностью фермента в мозге.
Лейцинаминопептидаза (КФ 3.4.11.1) проявляет широкую субстратную специфичность и отщепляет практически любые N-концевые аминокислоты, за исключением аланина и цистеина [56]. Она гидролизует амид лейцина лучше, чем ариламид лейцина, с меньшей скоростью расщепляет лизин- и аргининамид. Фермент имеет по данным Хрусталёвой [36] и Fraticante и соавт. [51] молекулярную массу около 62 кДа, по данным Shimamura и соавт. [74] – 53000, по данным Gibson и соавт [56] – 270 кДа, что позволяет предположить возможность существования субъединичной структуры. Он проявляет максимальную активность при нейтральных и слабощелочных значениях pH, активируется ионами Mn2+
, ингибируется ПХМФС, бестатином и амастатином; пуромицин и арфаменин A не влияют на его активность [72]. Фермент отщепляет остаток тирозина от Tyr-Gly-Gly, Tyr-Gly-Gly-Phe, Leu-энкефалина, динорфина-(1-8), динорфина-(1-10) и динорфина-(1-13), максимальная скорость расщепления наблюдается в случае Leu-энкефалина, с удлинением цепи скорость расщепления субстрата резко уменьшается [56]. Активность фермента равномерно рапределена между серым и белым веществом коры полушарий. Он обнаружен как в цитозольной так и мембранной фракциях головного мозга, особенно высокая активность найдена в миелине. Считают, что лейцинаминопептидазе принадлежит важная роль в инактивации энкефалинов, вещества Р и, возможно, большинства других регуляторных пептидов [36].
1.3 Механизм адаптации к физической работе. Роль регуляторных пептидов
Одним из эндогенных пептидов, широко представленных в разных отделах мозга, в том числе и в эмоциогенных зонах гипоталамуса, является вещество Р. Известно, что вещество Р способно оказывать непосредственное влияние на активность центральных нейронов, в большинстве случаев возбуждая их. Вместе с тем была отмечена способность вещества Р изменять реакции нейронов на нейромедиаторы. Доказано, что вещество Р способно снижать степень выраженности невротических состояний, нормализовать сон, улучшать память и процессы обучения, что позволяет рассматривать его как модулятор физиологических и патологических процессов.
Кроме того, вещество Р, содержащееся в нейронах задних рогов спинного мозга, способно передавать сигналы от периферических болевых рецепторов в центральные отделы нервной системы. Исследования на крысах позволили рассматривать вещество Р, синтезирующееся в гипоталамусе, как один из возможных пептидных факторов устойчивости к эмоциональному стрессу[37]. Вещество Р оказывает также модулярное влияние на метаболизм катехоламинов мозга при эмоциональном стрессе, выражающееся в способности вызвать долговременные изменения содержания норадреналина и дофамина в гипоталамусе и среднем мозге в сторону повышения, что расценивается как проявление центральных нейрохимических механизмов адаптации к эмоциональному стрессу[37].
Экспериментальные данные показывают, что содержание вещества Р в гипоталамусе коррелирует с устойчивостью к эмоциональному стрессу [48]. У больных неврозом отмечены снижение содержания вещества Р в крови и расстройства сна. У экспериментальных животных, подвергнутых эмоциональному стрессу, также нарушается электроэнцефалографическая структура сна. Сон является антистрессовым фактором. Во время бодрствования содержание вещества Р в крови снижается. При полноценном сне его уровень вновь повышается. Введение в организм животных, подвергнутых эмоциональному стрессу, вещества Р восстанавливает нормальную структуру сна. Таким образом, эмоциональный стресс, нарушая сон, лишает организм одного из защитных механизмов, с помощью которого повышается содержание вещества Р и обеспечивается устойчивость к стрессу. Учитывая то, что организм способен сам синтезировать вещество Р в большей степени во время сна, с одной стороны, и то, что содержание этого вещества в гипоталамусе коррелирует с устойчивостью к эмоциональному стрессу, с другой стороны, встает вопрос: каким путем, кроме полноценного сна, можно добиться ускоренного и полного восстановления в организме уровня вещества Р.
Одним из путей достижения этой цели, на наш взгляд, является использование рефлексотерапевтических методов (игло-, электро- и прессотерапии). Воздействуя на определенные биологически активные точки организма, связанные с гипоталамусом и средним мозгом, можно "запустить" выработку данными структурами мозга вещества Р. Физиологический эффект вещества Р как одного из эндогенных пептидов проявляется в модуляторном действии, выражающемся в способности влиять на нейрохимические свойства и катехо-ламиновый метаболизм нейронов, участвующих в формировании эмоциональных реакций, и тем самым прекращать нейромедиаторную интеграцию отрицательного эмоционального возбуждения, от которой зависит продолжительность отрицательной эмоциональной реакции, а значит и возможность развития эмоционального стресса.
В работах М.А. Звягинцевой (1988) [23], К.В. Судакова (1989) [32] разрабатываются способы коррекции и купирования стрессовых реакций и вызванных ими нарушений кровообращения с помощью эндогенных нейропептидов, а именно субстанции Р и пептида дельта-сна. Как показали исследования, субстанция Р обладает антистрессовым действием, улучшает функциональные состояния головного мозга и нормализует АД. Синтез в мозге эндогенных пептидов (вещества Р и, возможно, других эндогенных пептидов) может быть одним из факторов, определяющих генетические и индивидуальные различия в устойчивости к острому и хроническому эмоциональному стрессу и особенность метаболизма биогенных аминов в различных структурах мозга у устойчивых и предрасположенных к эмоциональному стрессу особей. Другими эндогенными пептидами, регулирующими эмоциональные реакции, в том числе и стрессовые, помимо вещества Р, являются эндорфины и энкефалины. Они так же, как и вещество Р, широко представлены в различных отделах мозга, в том числе в эмоциогенных зонах лимбической системы и в промежуточной доле гипофиза[4].
Эндорфины и энкефалины обладают необычайной способностью подобно морфину и героину снимать болевые ощущения. Это так называемые естественные опиаты. Тот факт, что морфин, героин и эндорфины связываются в одних и тех же местах, позволяет предположить, что эндорфины играют роль и в тех разновидностях эмоций, которые не имеют прямого отношения к боли. Ф. Блум и соавт. (1988) [4] показали, что у экспериментальных животных и человека при стрессе происходит выработка и высвобождение эндорфинов в нервных сетях. Высвободившиеся эндорфины, по всей вероятности, действуют двояко. С одной стороны, как опиаты, с другой - как регуляторы эмоциональных (стрессовых) реакций. Как опиаты, они блокируют высвобождение в синапсах задних рогов спинного мозга вещества Р, выделяемого из медленных (безмиелиновых) волокон, проводящих болевые импульсы от болевых рецепторов. В результате этого постсинаптический нейрон подвергается более слабой стимуляции веществом Р и головной мозг получает меньше болевых импульсов. Как регуляторы эмоциональных реакций, они каким-то образом, по всей вероятности через лимбическую систему, регулируют возбуждение, страх и другие стрессовые состояния в соответствии с ситуацией [35].
В настоящее время протеолиз рассматривается не только как процесс катаболической утилизации биологически активных пептидов, но и как регуляторный фактор, функция которого состоит в запуске и прерывании ряда биохимических и физиологических процессов при различных функциональных состояниях организма. Практически неизученным остается вопрос об изменениях в функции ферментов обмена нейропептидов при физической работе, в то время как именно активность этих ферментов определяет уровень биологически активных пептидов в организме и, следовательно, степень адаптации организма к физической работе. Физическую работу можно отнести к факторам стресса.
Обнаружено, что у устойчивых к стрессу животных в гипоталамусе и стриатуме активность КПN при воздействии стресса повышается. Авторами высказано предположение, что такой эффект наблюдается в связи с активацией синтеза в исследованных отделах нейропептидов (энкефалинов, вещества Р и др.), играющих ключевую роль при адаптации к стрессу [38].
Особый интерес представляют исследования, касающиеся влияния различных веществ на ферменты обмена нейропептидов при стрессе. Известно, что этанол ослабляет некоторые физиологические проявления стресса, усиливает секрецию стресс-пептидов, а так же активирует энкефалинэргическую систему. Поскольку уровень опиоидных и стресс-пептидов в организме контролируется КПN, то представляется возможным, что КПN определяет характер влияния этанола на организм при стрессе. Характер влияния этанола на физиологические проявления стресса связан с особенностями стрессирующего фактора. Сведения о гиперактивации КПN при совместном действии этанола иммобилизационного или хронического ЭБС в отделах мозга, где синтезируются опиоидные пептиды, вещество Р, подтверждают данные об адаптогенном действии этанола при стрессе. Однако такая активация пептидэргических систем ведет к тяжелым последствиям для организма, так как вызывает более быстрое истощение этих систем.
АПФ участвует в деградации энкефалинов, вещества Р и ПВДС - биологически активных пептидов, основная роль которых заключается в адаптации организма к стрессу [8].
Таким образом, изменения в проявлении функциональной активности ферментов процессинга и инактивации биологически активных пептидов при стрессе свидетельствуют о важной роли этих ферментов в регуляции уровня активных нейропептидов, участвующих, как в развитии, так и в торможении размаха стресс-реакции.
В исследовании использовали сыворотку крови, полученную путем центрифугирования крови при 4000 об/мин в течении 30 мин. Забор крови осуществлялся из локтевой вены у спортсменов квалификации мастер спорта и мастер спорта международного класса, вид спорта: легкая атлетика, средний бег и триатлон (n=12). В качестве контрольного материала использовали сыворотку крови студентов и аспирантов ВУЗов города Пензы (n=12).
Каждая группа делилась на две подгруппы – до нагрузки и на субмаксимальной нагрузке. Физическую работу создавали с помощью программируемого тредбана Tunturi 90, начиная со скорости 3,5 м/с, повышая каждые две минуты на 0,5 м/с до скорости, характеризующейся подъемом пульса до 180 ударов в минуту, на которой испытуемый бежал до состояния полного утомления. Пробы крови отбирались из локтевой вены до нагрузки и непосредственно после остановки тредбана.
Концентрацию вещества Р определяли иммуноферментным методом ELISA [42] в сыворотке крови. Для получения сыворотки кровь центрифугировали 30 мин при 4000 об/мин.
2.2.3 Метод определения активности КПN
Сыворотку крови (40 мкл) смешивали с 20 мкл 35 мкМ СоSO4 в 100 мМ трис-НСl буфере, рН=7,6,и преинкубировали 8 мин при 37°С. Реакцию начинали прибавлением 10 мкл кбз-Gly-Arg в вышеуказанном буфере. Через 120 мин реакцию останавливали прибавлением 30 мкл 10% трихлоруксусной кислоты. Пробы центрифугировали 30 мин при 4000 об/мин. Отбирали 50 мкл надосадочной жидкости и определяли количество образовавшегося Arg нингидриновым методом (добавляли 1 мл нингидрина). Далее инкубация 12 мин на кипящей водяной бане. Затем пробы колориметрировали на КФК-2 при λ=590 нм. Активность КПN определяли по отщеплению Arg от кбз-Gly-Arg как Со²+
- активируемую. Активность фермента выражали в нмоль Arg, образовавшегося за 1 мин инкубации в пересчете на 1 мг белка. Количество белка в пробах определяли по методу Лоури.
Активность ангиотензинпревращающего фермента определяли по образованию Gly-Arg из кбз-Gly-Gly-Arg при рН=8,2, как активность, ингибируемую каптоприлом. Сыворотку крови (40 мкл) смешивали с 20 мкл 35 мкМ каптоприла в 100 мМ трис-НСI буфере,рН=8,2, или 20 мкл буфера и преинкубировали 8 мин при 37°С. Реакцию начинали прибавлением 10 мкл раствора кбз-Gly-Gly-Arg в вышеуказанном буфере. Через 120 мин реакцию останавливали прибавлением 30 мкл 10% трихлоруксусной кислоты. Пробы центрифугировали 30 мин при 4000 об/мин. Отбирали 50 мкл надосадочной жидкости и определяли количество образовавшегося Gly-Arg нингидриновым методом (добавляли 1 мл нингидрина). Далее инкубация 12 мин на кипящей водяной бане. Пробы колориметрировали на КФК-2 при λ=590 нм. Активность ангиотензинпревращающего фермента определяли как разность в оптической плотности проб не содержащих и содержащих каптоприл. Активность фермента выражали в нмоль Gly-Arg, образовавшегося за 1 мин инкубации в пересчете на 1 мг белка. Количество белка в пробах определяли по методу Лоури.
Для определения активности лейцинаминопептидазы 90 мкл сыворотки крови смешивали с 10 мкл раствора пуромицина, приготовленного на соответствующем буфере (в случае опытной пробы (I)) и к 10 мкл 20 мМ фосфатного буфера (рН 7,4) (в случае опытной пробы (II) и контрольной пробы). Далее проводили преинкубацию 8 мин при 37˚С. Реакцию начинали прибавлением в опытные пробы 100 мкл 310 мкМ раствора лей-b-нафтиламина, приготовленного на буфере. Через 30 мин реакцию останавливали прибавлением 40 мкл 10% раствора ТХУ. В контрольные пробы добавляли 100 мкл субстрата. Пробы центрифугировали 30 мин при 4000 об/мин. Отбирали 100 мкл надосадочной жидкости, добавляли по 1,5 мл натрий-ацетатного буфера (рН 4,2). Измерение флюоресценции образовавшегося b-нафтиламина проводили на флюориметре ФМЦ-2 при λex
=360 нм и λem
=420 нм в кювете толщиной 1 см. Активность фермента определяли как разность между опытной пробой (I) и контрольной пробой и выражали в мкмоль b-нафтиламина, образовавшегося за 1 мин инкубации в пересчете на 1 мг белка. Белок определяли методом Лоури.
Полученные результаты обрабатывали статистически с использованием t-критерия Стьюдента. Полученные отличия в значениях между опытной и контрольной группами считали достоверными при р <0,05.
ГЛАВА 3. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
И ИХ ОБУЖДЕНИЕ
Содержание вещества Р в сыворотке крови спортсменов в покое было на 300% выше, чем у неспортсменов. При физической работе у спортсменов не наблюдалось изменений в его концентрации, в то время как у неспортсменов происходило повышение уровня вещества Р на 60%.

Рис. 1 Влияние максимальной физической нагрузки на уровень вещества Р в крови спортсменов и неспортсменов в норме и при физической работе. Условные обозначения:  - спортсмены, - спортсмены,  - неспортсмены. - неспортсмены.
*** - Р < 0,001 по сравнению с физиологическим состоянием, +++ - Р < 0,001 по сравнению с неспортсменами.
Активность пептидил-дипептидазы А, карбоксипептидазы N и лейцинаминопептидазы у спортсменов в физиологическом состоянии была выше, чем у людей, не занимающихся спортом. После анаэробной работы у спортсменов не наблюдалось изменений в активности исследуемых ферментов, в то время как у неспортсменов происходило повышение активности данных пептидгидролаз.

Рис. 2 Влияние максимальной физической нагрузки на активность пептидилдипептидазы А в сыворотке крови спортсменов. Условные обозначения:
 - спортсмены, - неспортсмены. - спортсмены, - неспортсмены.
** - Р < 0,01 по сравнению с физиологическим состоянием,
++ - Р < 0,01 по сравнению с неспортсменами.
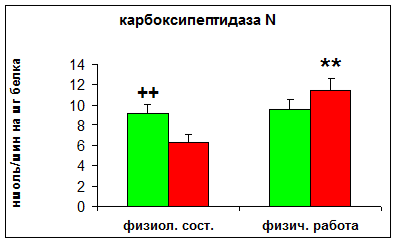
Рис. 3 Влияние максимальной физической нагрузки на активность карбоксипептидазы N в сыворотке крови спортсменов. Условные обозначения:
- спортсмены, - неспортсмены.
** - Р < 0,01 по сравнению с физиологическим состоянием,
++ - Р < 0,01 по сравнению с неспортсменами.

Рис. 3 Влияние максимальной физической нагрузки на активность лейцинаминопептидазы в сыворотке крови спортсменов. Условные обозначения: - спортсмены, - неспортсмены.
** - Р < 0,01 по сравнению с физиологическим состоянием,
+++ - Р < 0,001 по сравнению с неспортсменами.
Результаты нашего исследования показывают, что существенную роль в адаптации к физической работе у спортсменов высокой квалификации играет вещество Р, обладающее широким спектром действия – модулирующее болевую чувствительность, повышающее устойчивость к стрессу. Одним из механизмов регуляции уровня вещества Р при физической работе является изменение активности ферментов его обмена. Адаптационные перестройки функционирования пептидергической системы приводят к значительной активизации и стабилизации ее работы – в физиологическом состоянии у спортсменов все ее компоненты работают интенсивнее, чем у неспортсменов, и ее функционирование не изменяется при совершении физической работы у спортсменов высокой квалификации.
Таким образом, пептидергическая система активно вовлечена в адаптивные процессы к физической работе и перестройки в ее функционировании являются одними из тех факторов, от которых зависит спортивный результат.
ВЫВОДЫ
1. Физическая работа вызывает существенные адаптационные изменения в функционировании пептидергической системы. У спортсменов высокой квалификации в физиологическом состоянии по сравнению с неспортсменами все компоненты системы работают интенсивнее.
2. Существенную роль в адаптации к физической работе у спортсменов высокой квалификации играет вещество Р, обладающее широким спектром действия – модулирующее болевую чувствительность, повышающее устойчивость к стрессу.
3. Одним из механизмов регуляции уровня вещества Р при физической работе является изменение активности ферментов его обмена – пептидил-дипептидазы А, карбоксипептидазы N и лейцинаминопептидазы.
Список литературы
1. Ашмарин И.П., Каразеева Е.П. Нейропептиды // в кн. “Нейрохимия” под ред. Ашмарина И.П., Стукалова П.В.- М.: Издательство института биомедицинской химии РАМН.-1996.- С.296-333.
2. Бабичев В.Н. Нейроэндокринный контроль процессов пубертации // Усп. совр. биол. – 1994. – 114, № 3. – С. 330-344.
3. Бабичев В.Н., Миронов С.Ф. Нейропептиды мозга и их нейроэндокринные эффекты // Пробл. эндокринол. – 1981. – № 3. – С. 78-85.
4. Блум Ф., Лейзерсон А., Хофстедтер Л. Мозг, разум и поведение / Пер. с англ. - М.: Мир, 1988. - 248 с.
5. Бэйрд Д.Т. Яичник / Гормональная регуляция размножения у млекопитающих. – М.: Мир. – 1987. – С. 118-144.
6. Вернигора А.Н., Генгин М.Т. Механизм регуляции активности и биологическая роль карбоксипептидазы H – фермента процессинга нейропептидов // Биохимия - 1995. - 60, № 12. - С. 1491-1497.
7. Вернигора А.Н., Генгин М.Т. Протеолитические ферменты: субклеточная локализация, свойства и участие в обмене нейропептидов // Биохимия. – 1996. - 61, № 5. - С. 771-785.
8. Вернигора А.Н., Генгин М.Т., Никишин Н.Н. Об участии некоторых ферментов нейропептидов в механизмах эмоционального стресса // Физиол. журн.-1995.- т.81.- №5.- С.1025-1028
9. Вернигора А.Н., Генгин М.Т., Никишин Н.Н. Множественность молекулярных форм растворимых карбоксипептидазо-В-подобных ферментов головного мозга кошки // Укр. биохим. журн. – 1993.– 65, № 4. – С. 17-21.
10. Вернигора А.Н., Никишин Н.Н., Генгин М.Т. О взаимосвязи между активностью карбоксипептидазы Н и ангиотензинпревращающего фермента // Биохимия. – 1995. – 60, N 1. – С. 144-149.
11. Генгин М.Т. Особенности структурно-функциональной организации и физико-химические свойства нелизосомальных пептидгидролаз мозга животных: Автореф. дис. … док. биол. наук. – Пенза, 2002.
12. Генгин М.Т., Вернигора А.Н., Никишин Н.Н., Щетинина Н.В. Активность карбоксипептидазы N и ангиотензинпревращающего фермента в сыворотке крови крыс в норме и при эмоциональном стрессе // Укр. биохим. журн. – 1994. – 66, № 2. – С. 139-142.
13. Генгин М.Т., Соловьев В.Б., Сметанин В.А., Пазялова А.А., Симкина О.В., Коновалов А.Н., Кузичкин Д.С., Шашкина Н.К., Генгина Н.М., Субботина К.Б. Влияние атропина на активность ангиотензин-превращающего фермента и карбоксипептидазы N в сыворотке крови крыс // Украинский биохимический журнал. – 2005. – Т. 77, № 6. – С. 78-80.
14. Гомазков О.А. Функциональная биохимия регуляторных пептидов.– М.: Наука. 1993.
15. Гомазков О.А. Энзимологические основы физиологического действия регуляторных пептидов // Биологические науки. – 1986. – № 2. – С. 13-23.
16. Гомазков О.А., Комиссарова Н.В., Петрий О.П., Панфилов А.Д. Возрастные и регионарные изменения ангиотензинпревращающей и кининдеградирующей активности в мозге агрессивных крыс // Бюлл. эксперим. биол. мед. – 1987. – 104, № 7. – С. 18-20.
17. Гомазков О.А., Панфилов А.Д., Комиссарова Н.В., Ростовцев А.П. Влияние длительного потребления этанола на физиологическое состояние и изменения активности пептидаз мозга у мурицидных (агрессивных) крыс // Журн. высш. нервн. деят. – 1992. – 42, № 4. – P. 771-777.
18. Гомазков О.А., Панфилов А.Д., Ростовцев А.П., Комиссарова Н.В., Фомин В.В., Григорьянц О.О. Регионарная активность энкефалин- и ангиотензин II-образующих пептидаз мозга и периферических тканей у крыс с различным влечением к этанолу // Вопр. мед. химии. – 1991. – 37, N 4. – С. 33-37.
19. Гормонотерапия: Пер. с нем. / Под ред. Шамбаха Х., Кнаппе Г., Карола В. – М.: Медицина, 1988, 416 с.
20. Дмитриев А.Д. Биосинтез нейропептидов // Итоги науки и техники ВИНИТИ. Фармакол. химиотерапевт. средства. – 1982. – 43. – С. 7-49.
21. Држевецкая И.А. Основы физиологии обмена веществ и эндокринной системы. – М.: Высш. шк., 1994, 256 с.
22. Замятнин А.А. Общие функциональные особенности зндогенных регуляторных олигопептидов // Физиол. ж. – 1992. – 78, № 9. – С. 39-49.
23. Звягинцева М.А. Роль пептида дельта-сна в электрической стабильности сердца // Кардиология. - 1988. - №3. - С. 89-91.
24. Корнева Е.А. Регуляторные пептиды как модуляторы защитных функций организма // Физиол. ж. – 1985. – 75, № 5. – С. 656-665.
25. Кост О.А., Ламзина Н.А., Казанская Н.Ф. Микрогетерогенность ангиотензин-превращающего фермента, как фактор регуляции его в организме // Механизмы регуляции клеточной активности: (Ташкент, 18 – 22 сент. 1989 г.): Тез. докл. – Москва, 1989. – С. 40.
26. Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека: В 2-х томах. Пер. с англ. / Под ред. Гинодмана Л.М. – М.: Мир, 1993.
27. Никишин Н.Н., Вернигора А.Н., Генгин М.Т. Активность ангиотензинпревращающего фермента в отделах головного мозга крыс, обладающих различной устойчивостью к эмоциональному стрессу в норме и при стрессе // Укр. биохим. журн. – 1994. – 66, № 5. – С. 53-57.
28. Панченко Л.Ф., Брусов О.С., Беляев Н.А. Исследование механизма действия этанола на активность энкефалиназы A мозга крыс // Бюлл. эксперим. биол. мед.– 1984.– 97, № 6.– С. 691-692.
29. Реминяк В.И. Некоторые нейрохимические критерии прогноза эффективности терапии атропиновыми комами больных шизофренией // Тезисы докл. IX Всесоюз. конференции по биох. НС. – 1983. – Ереван.
30. Сахаров И.Ю., Данилов С.М., Духанина Е.А. Получение и молекулярно-кинетические свойства ангиотензинпревращающего фермента из сердца человека // Биохимия. – 1986. – 51, N 11. – С. 1836-1842.
31. Соловьев В.Б., Генгин М.Т. Активность пептидил-дипептидазы А и карбоксипептидазы N в сыворотке крови пациентов с болезнью Альцгеймера // Украинский биохимический журнал. – 2007. – Т. 79, № 6. – С. 103-105.
32. Судаков К. В. Олигопептиды в механизмах устойчивости к эмоциональному стрессу//Патол. физиология и эксперим. терапия.-1989.-Вып. 1.-С.3-11.
33. Сухоруков В.С., Тарабрин С.Б. Роль пролактина в регуляции функций мужской гонады // Усп. совр. биол. – 1993. – 113, № 3. – С. 366-376.
34. Теппермен Дж., Теппермен У. Физиология обмена веществ и эндокринной системы. Вводный курс: Пер. с англ. / Под ред. Ажипа Я.И. – М.: Мир, 1989.
35. Федоров Б. М. Стресс и система кровообращения. -М.: Медицина, 1991. - 320с
36. Хрусталева Н.А. Выделение, очистка, некоторые физико-химические свойства лейцинаминопептидазы миелина и диагностическое значение этого фермента при демиелинизирующих заболеваниях // Автореф. канд. дис. – Москва. – 1987.
37. Юматов Е.А. Пептидно-нейромедиаторные механизмы устойчивости к эмоциональному стрессу // Стресс и психологическая патология. - М.: Московский НИИ психиатрии, 1983. - С. 7-12.
38. Юматов Е.А., Гехт К., Скоцеляс Ю.Г. Субстанция Р как фактор устойчивости к эмоциональному стрессу // Журн. выс. нервн. деятельности.-1984.-34.-4.-С.771-777.
39. Azaryan A.V., Hook V.Y.H. Distinct properties of prohormone thiol protease (Ptp) compared to cathepsin B, cathepsin L, and cathepsin H - evidence for Ptp as a novel cysteine protease // Arch. Biochem. Biophys. – 1994. – 314, N 1. – P. 171‑177.
40. Azaryan A.V., Hook V.Y.H. Unique cleavage specificity of prohormone thiol protease related to proenkephalin processing // FEBS Lett. – 1994. – 341, N 2-3. – P. 197-202.
41. Barnes K., Turner A.J., Kenny A.J. Membrane localization of endopeptidase-24.11 and peptidyl dipeptidase A (angiotensin converting enzyme) in the pig brain: a study using subcellular fractionation and electron microscopic immunocytochemistry // J. Neurochem. – 1992. – 58, N 6. – P. 2088-2096.
42. Bouassida A. et al. // J. of Sports Science and Med. 2006, V 5, P. 172 – 181
43. Clemens D.L., Okamura T., Inagami T. Subcellular localization of angiotensin-converting enzyme in cultured neuroblastoma cells // J. Neurochem. – 1986. – 47, N 6. – P. 1837-1842.
44. Chesselet M.F., Hook V.Y. Carboxypeptidase H-like immunoreactivity in the striatum of cats and monkeys // Regul. Pept. – 1988. – 20, N 2. – P. 151-159.
45. Davidson H.W., Hutton J.C. The insulin-secretory-granule carboxypeptidase H: Purification and demonstration of involvement in proinsulin processing // Biochem. J. – 1987. – 245, N 2. – P. 575-582.
46. Demmer W., Brand K. A dipeptidyl carboxypeptidase in brain synaptic membranes active in the metabolism of enkephalin containing peptides // Biochem. Biophys. Res. Commun. – 1983. – 114, N 2. – P. 804-812.
47. Devi L. Tissue distribution of a dynorphin-processing endopeptidase // Endocrinology. – 1993. – 132, N 3. – P. 1139-1144.
48. Dupont A.. Sabord P., Merand G. el al. Age-related changes in central nervous system eukephalins substance P // Life Sci. - 1981. - Vol. 29. - №22. - P. 2317-2322.
49. Edwards C.R., Padfield P.L. Angiotensin-converting enzyme inhibitors: past, present and bright future // Lancet. – 1985. – 8419. – P. 30-34.
50. Erdos E.G., Skidgel R.A. Angiotensin-I-converting enzyme // Lab. Inuest. – 1987. – 56, – N 4. – P. 345-348.
51. Fraticante L.I., Rotrosen I., Siekiorski I., Fracer H., Gershon S. Enkephalin inactivation by N-terminal tyrosine cleavage: purification and partial characterisation of a highly specific enzyme from human brain // Life Sci. – 1980. – 26, N 20. – P. 1697-1706.
52. Fricker L.D., Plummer T.H., Snyder S.H. Enkephalin convertase: potent, selective and irreversible inhibitors // Biochem. and Biophys. Res. Commun. – 1983. – 11, N 3. – P. 994-1000.
53. Fricker L.D., Snyder S.H. Enkephalin convertase: purification and charasterization of a specific enkephalin-synthesizing carboxypeptidase localized to adrenall chromaffin granules // Proc. Natl. Acad. Sci. USA. – 1982. – 79. – P. 3886-3890.
54. Fricker L.D., Snyder S.H. Purification and characterization of enkephalin convertase, an enkephaline-synthesizing carboxypeptidase // J. Biol. Chem. – 1983. – 258, N 18. – P. 10950-10955.
55. Gainer H., Russel J.T., Loh Y.P. The enzymology and intracellular organization of peptide precursor processing: the secretory vesicle hypothesis // Neuroendocrinology. - 1985. – 40, N 1. – P. 171-184.1992. – 131, N 3.
56. Gibson A.M., Biggins J.A., Lauffart B., Mantle D., McDermott J.R. Human brain leucyl aminopeptidase: Isolation, characterization and specificity against some neuropeptides // Neuropeptides. – 1991. – 19, N 3. – P. 163-168.
57. Jukic D. Neurokinin receptors antagonists: old and new. // Life Sci. 1991, Vol. 49, P. 1463.
58. Hook V.Y. Carboxypeptidase B-like activity for the processing of enkephalin precursors in the membrane component of bovine adrenomedullary chromaffin granules // Neuropeptides. – 1984. – 4, N 2. – P. 117-126.
59. Hook V.Y.H., LaGamma E.F. Product inhibit of carboxypeptidase H // J. Biol. Chem. – 1987. – 262, N 26. – P. 12583-12588.
60. Hooper N.M. Angiotensin converting enzyme: implications from molecular biology for its physiological functions // Int. J. Biochem. – 1991. – 23, N 7/8. – P. 641-647.
61. Hooper N.M., Turner A.J. Charasterization of angiotensin-converting enzyme from pig brain // Biochem. Soc. Trans. – 1986. – 14, N 6. – P. 1249-1250.
62. Hughes J. (ed.). Centrally Acting Peptides, University Park Press, 1978. Iversen S.D., Iversen L.L. Behavioral pharmacology, Oxford University Press, second
63. Krause J. // PNAS USA 1987, Vol. 84, P. 881.
64. L.D. Peptide processing exopeptidases: amino- and carboxypeptidases involved with peptide biosynthesis // Peptide biosynthesis and processing (Fricker L.D. ed.), CRC Press, Boca Raton, Florida, 1991. – P. 199-230.
65. Loh Y.P., Birch N.P., Castro M.G. Pro-opiomelanocortin and pro-vasopressin converting enzyme in pituitary secretory vesicles // Biochimie. – 1988. – 70, N 1. – P. 11-16.
66. Loh Y.P., Parish D.C., Tuteja R. Purification and charasterization of a paired basic residue-specific pro-opiomelanocortin converting enzyme from bovine pituitary intermediate lobe secretory vesicles // J. Biol. Chem. – 1985. – 260, N 12.
67. Lynch D.R., Venable J.C., Snyder S.H. Enkephalin convertase in the heart: similar disposition to atrial natriuretic factor // Endocrinology.– 122, N 6.– P. 2683-2691.
68. Maggi C. , Manzini S. Tachykmin Receptor Antagonists. // Drugs and the Lung 1994, 507-520.
69. Nawa H. // Nature 1983, Vol. 306, Р. 32.
70. Norenberg U., Richter D. Processing of the oxytocin precursor: isolation of an exopeptidase from neurosecretory granules of bovine pituitaries // Biochem. Biophys. Res. Commun. – 1988. – 156, N 2. – P. 898-904.
71. Rouille Y., Chauvet J., Acher R. Partial conversion of vasopressinyl-Gly-Lys-Arg into pharmacologically active vasopressin through secretory granule carboxypeptidase E and alpha-amidating processing enzymes // Biochem. Int. – 1992. – 26, N 4. – P.739-746.
72. Schnebli H.P., Phillipps M.A., Barclay R.K. Isolation and characterization of an enkephalin-degrading aminopeptidase from rat brain // Biochim. Biophys. Acta. – 1979. – 569, N 1. – P. 89-98.
73. Seidah N.G., Chretien M. Proprotein and prohormone convertases of the subtilisin family - recent developments and future perspectives // Trends Endocrinol. Met. – 1992. – 3, N 4. – P. 133-140.
74. Shimamura M., Hazato T., Iwaguchi T. A new aminopeptidase in monkey cerebral membrane fraction: hydrolysis of enkephalin // Brain Res. – 1988. – 445. – P. 350-353.
75. Skidgel R.A., Erdos E.G. The broad substrate specificity of human angiotensin I converting enzyme // Clin. Exp. Hypertens. – 1987. – A9, N 2/3. – P. 243-259.
76. Smyth D.G., Maruthainar K., Darby N.J., Fricker L.D. Catalysis of slow C terminal processing reactions by carboxypeptidase H // J. Neurochem. – 1989. – 53, N 2. – P. 489-493.
77. Steiner D.F. The biosynthesis of biologically active peptides: a perspective // Peptide Biosynthesis and Processing (Fricker L.D., ed.).– CRC Press, Boca Raton, Florida, 1991.– P. 1-16.
78. Supattapone S., Fricker L.D., Snyder S.H. Purification and characterization of a membrane-bound enkephalin-forming carboxypeptidase, «enkephalin convertase» // Neurochem. – 1984. – 42, N 4. – P. 1017-1023.
|