Министерство образования Республики Беларусь
Учреждение образования
«Гомельский государственный университет
имени Франциска Скорины»
Биологический факультет
Кафедра химии
Допущено к защите
Зав. кафедрой __________ Шумилин В.А
« _____ » ______________ 2009г
ФЛУОРЕСЦЕНТНЫЙ МЕТОД ИССЛЕДОВАНИЯ КОНФОРМАЦИОННЫХ ИЗМЕНЕНИЙ АЛЬБУМИНА В РАСТВОРЕ
Дипломная работа
Исполнитель:
студент группы Б-51 ____________ Корноушенко Юрий Валерьевич
Научный руководитель:
к.х.н., доцент ____________ Дроздова Наталья Ивановна
Рецензент:
к.б.н., ассистент кафедры ____________ Галиновский Николай
зоологии и охраны природы Генадьевич
Гомель 2009
СОДЕРЖАНИЕ
| Введение……………………………………………………………………….. |
3 |
1 ОБЗОР ЛИТЕРАТУРЫ……………………………………………………...
|
4
|
| 1.1 Определение понятия «альбумин»………………………………………. |
4 |
| 1.2 Современные представления о структуре молекулы сывороточного альбумина……………………………………………………………………… |
4 |
| 1.2.1 Гетерогенность альбумина……………………………………………... |
9 |
| 1.2.2 Характеристика центров связывания лигандов в альбумине………... |
11 |
| 1.3 Влияние физико-химических факторов на альбумин………………….. |
13 |
| 1.3.1 Влияние химических модификаторов на альбумин………………….. |
14 |
| 1.4 Функции сывороточного альбумина в организме человека…………… |
17 |
| 1.5 Основные понятия и закономерности люминесценции……………….. |
19 |
| 1.6 Применение зондовой флуоресценции для изучения структуры молекулы альбумина…………………………………………………………. |
20
|
| 2 ОБЪЕКТ, ПРОГРАММА И МЕТОДИКА ИССЛЕДОВАНИЙ………….. |
24 |
| 3 РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ И ИХ ОБСУЖДЕНИЕ……………... |
27 |
| 3.1 Исследование влияния сверхвысоких концентраций мочевины на конформацию альбумина…………………………………………………….. |
27
|
| 3.2 Влияние свободных радикалов на структуру молекулы альбумина….. |
33 |
| ЗАКЛЮЧЕНИЕ……………………………………………………………….. |
37 |
| СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ………………………… |
38 |
ВВЕДЕНИЕ
Интерес к исследованию альбумина, неугасающий за последние 40 лет, связан с открытием всё новых его функций в организме. Уже одна цифра, указывающая его содержание в крови (60% - от всех белков плазмы), говорит о ключевом значении альбумина для выполнения кровью своих функций. С другой стороны его распространённость в различных физиологических жидкостях (интерстициальной, лимфе), а также различных тканях говорит о большой важности именно этого белка для организма, по сравнению с другими.
Для доказательства таких выводов, надо обратиться к строению альбумина. В 70-х – 80-х годах была расшифрована аминокислотная последовательность и в целом пространственная структура сывороточного альбумина. Оказалось, что уникальное строение альбумина, позволяет ему связывать различные по своей химической природе вещества (альдегиды, алканы, жирные кислоты, металлы постоянной и переменной валентности и м.д. низкомолекулярные вещества) и переносить их в крови. Таким образом, ключевая функция альбумина – транспортная. Позже было выяснено участие альбумина в антиокислительных процессах, в поддержании онкотического давления плазмы и даже в направленной локомоции нейтрофилов в очаги воспаления и т.д.
Из выше изложенного можно заключить, что значение альбумина в поддержании гомеостаза огромно. Однако известно, что при связывании альбумина с различными лигандами, его конформация определённым образом изменяется и это может отразиться на его связывании с другими лигандами. Флуоресцентным методом можно зарегистрировать многие тонкие конформационные изменения альбумина и судить о качественном состоянии белка, а по качеству структуры альбумина, можно оценить и степень патологического состояния организма.
Цель работы:
изучение конформационных изменений бычьего сывороточного альбумина под действием свободных радикалов и сверхвысоких концентраций мочевины.
Практическое значение
работы заключается в том, что полученные данные могут быть использованы в медицине, при расшифровке патогенеза таких заболеваний как почечная недостаточность, цирроз печени. Также, используя мочевину в качестве денатурирующего агента, можно смоделировать процесс взаимодействия альбумина с различными лигандами в условиях метаболитного стресса.
Научная значимость:
в силу высокой чувствительности флуоресцентного метода и уникальности молекулы альбумина как объекта для изучения данным методом его структуры, полученные результаты могут иметь большое значение для химии глобулярных белков схожего с сывороточным альбумином строения.
1 ОБЗОР ЛИТЕРАТУРЫ
1.1
Определение понятия «альбумин»
В 1838 году G. Mulder обнаружил, что в тканях животных и растениях содержатся вещества, напоминающие по своим свойствам яичный белок. Постепенно было установлено, что белки представляют собой обширный класс веществ, построенных по определённому плану. Выяснилось, что белки можно разделять, осаждая их из растворов с различной концентрацией сульфата аммония. Согласно одной из первых классификаций белков (P. Panum, 1852) альбумины – это белки, остающиеся в растворе полунасыщенного сульфата аммония.
Спустя почти век этот метод разделения белков уступил место электрофоретическому методу. В 1937 году А. Тизелиус разработал способ аналитического разделения белков электрофорезом в растворе, затем для борьбы с тепловой конвекцией стали использовать растворы с градиентом плотности сахарозы и, наконец, перешли к электрофорезу на различных носителях (бумага, крахмальный гель и т.д.) [1].
Согласно молекулярно-биологической классификации альбумин относится к мультигенному семейству белков, включающему в себя α-фетопротеин, группоспецифический компонент, витамин D-связывающий протеин.
В медицине понятие «альбумин» относят только к одному виду белков человека, который обычно называют «сывороточным альбумином» (ЧСА).
Сывороточный альбумин человека (ЧСА) – это глобулярный белок с молекулярной массой около 66,4 килодальтон (кДа), не осаждающийся при pH 6 – 7 в полунасыщенном растворе сульфата аммония.. Главным местом синтеза ЧСА в организме является печень, где одна молекула этого белка синтезируется примерно за 20 минут, а в сутки – около 10 – 15 грамм альбумина. В плазме крови концентрация ЧСА – около 35-50 г/л, что составляет 47 – 62 % всех белков плазмы, в лимфе – 15 – 36 г/л, в межклеточной жидкости – 3 г/л, в ликворе – 0,3 г/л. Всего в кровеносном русле человека содержится около 120 – 140 грамм альбумина, во внеклеточном пространстве – около 360 грамм. Время существования молекулы альбумина в организме человека – около 20 дней [2].
1.2 Современные представления о структуре молекулы сывороточного альбумина
В настоящее время расшифровка первичной структуры человеческого и бычьего сывороточного альбумина полностью завершена. Необходимо отметить, что структура ЧСА и БСА достаточно тесно согласуются друг с другом, отличаясь лишь в некоторых незначительных деталях положения отдельных аминокислот. ЧСА состоит из 585 аминокислотных остатков, БСА – 582.
При сравнении первичной структуры альбуминов различных видов, в частности ЧСА и БСА, наиболее примечательной является стабильность положения заряженных аминокислот (Lys, Glu, Asp). Возможно, что локализацией этих остатков определяются значительные особенности данного белка. Так, значительное число заряженных групп и характерная для альбумина низкая изоэлектрическая точка является важным фактором его онкотической функции. Кроме того, большое число полярных остатков удерживает в растворе обширные гидрофобные участки белковой молекулы. Последний факт имеет первостепенное значение в фиксации альбумином низкомолекулярных соединений [3].
Интересной особенностью первичной структуры ЧСА, является наличие единственного остатка триптофана, который локализован в положении 214. БСА имеет два триптофановых остатка в положениях 134 и 212. Данный факт важен с методической точки зрения. Он позволяет с успехом применять спектрофлюориметрию при изучении конформационных перестроек альбумина, а учитывая, что БСА и ЧСА практически идентичны и в БСА содержится два триптофанила, которые могут давать большую информацию о перестройках альбумина, чем один, можно с полной достоверностью переносить результаты, получившиеся с БСА в эксперименте, на ЧСА.
Наряду с заряженными группами уникальное значение имеет расположение дисульфидных связей, определяющие своеобразное свёртывание полипептидной цепи. Всего в молекуле альбумина имеется 35 остатков цистеина, причём их локализация в ЧСА и БСА абсолютно тождественна.
Основная заслуга в расшифровке аминокислотной последовательности БСА и ЧСА принадлежит J. Brown. Его модель трёхмерного строения альбумина признаётся наиболее обоснованной и тщательно разработанной.
J. Brown (1977) обратил внимание на тот факт, что характерной чертой первичной структуры альбуминов является многократное повторение спаренных остатков Cys-Cys. Всего в молекуле белка имеется 7 таких пар в положениях 90 – 91, 168 – 169, 245 – 246, 360 – 361, 437 – 438, 476 – 477 и 558 – 559 (рисунок 1).
Вправо (по направлению к С-концу цепи) от каждой такой пары через 7 – 10 остатков расположен одиночный остаток Cys. Влево от пяти из имеющихся семи пар через 43 – 45 остатков также расположен одиночный Cys. Локализованные описанным образом остатки Cys, связываясь дисульфидными мостами, образуют пять больших двойных петель (рисунок 2,а). В остальных двух случаях одиночный Cys лежит на 15 остатков левее пары Cys–Cys. При этом образуются две малые двойные петли (рисунок 2, б).
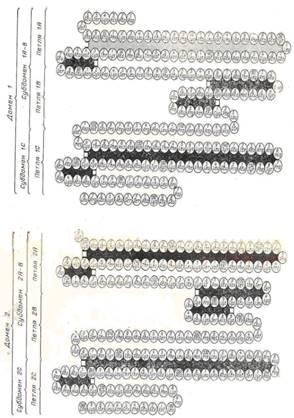
Рисунок 1 – Схематическое изображение структурной организации сывороточного альбумина человека [4]
 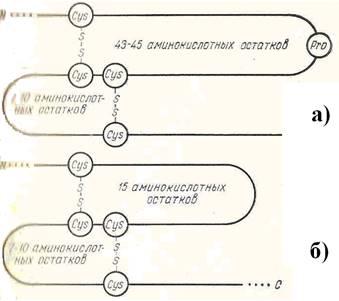
Рисунок 2 – Схематическое изображение образования больших (а) и малых (б) двойных петель молекулы альбумина [5]
Ещё одна малая двойная петля в середине цепи (256 – 289) имеет несколько иное строение. В центре два Cys лежат не рядом, как в остальных случаях, а разделены тремя аминокислотами (Pro – Glu – Lys). Кроме того вблизи N-конца имеется одна одинарная петля, включающая 10 аминокислот (53 – 62).
Важной особенностью первичной структуры ЧСА, является наличие единственного остатка триптофана, который локализован в положении 214 (рисунок 3).

Рисунок 3 – Расположение остатка триптофана в молекуле ЧСА [6]
БСА имеет два триптофановых остатка в положениях 134 и 212 (рисунок 2). Данный факт важен с методической точки зрения. Он позволяет с успехом применять спектрофлюориметрию при изучении конформационных перестроек альбумина, существенно облегчая интерпретацию полученных результатов.
Таким образом, согласно модели, предложенной J. Brown, молекула альбумина содержит 5 больших двойных петель, 3 малых и 1 одинарную, в образовании которых участвует 17 дисульфидных связей. Единственная свободная SH-группа принадлежит цистеину, локализованному в ближайшем к N-концу положении (Сys34
) (рисунок 1). Этой сульфгидрильной группе отводится особое место в плане регуляции конформационных переходов белковой молекулы.
Характерно, что только в двух из пяти больших двойных петель имеет место одинаковое сочетание из четырёх аминокислот: Arg-Arg-His-Pro. Кроме того, вблизи вершины каждой большой петли имеется остаток Pro (рисунок 1, 2, а).
С разнообразием аминокислотного состава петель может быть связана, в числе прочих факторов, универсальность комплексообразующих свойств альбумина, способного фиксировать низкомолекулярные соединения разнообразного химического строения и стерической структуры.
Если представить, как это полагает автор модели, что свободный N-концевой участок ранее в эволюции был также связан с ныне существующей единственной одиночной петлёй в большую двойную петлю, то всю молекулу альбумина легко разделить на три очень сходных домена: 1 – 190, 191 – 382 и 383 – 585. Каждый домен состоит из большой двойной петли, короткого соединительного сегмента, малой двойной петли, длинного соединительного сегмента, ещё одной большой двойной петли и соединительного сегмента со следующим доменом.
Каждый из трёх доменов J. Brown подразделил на два субдомена (А – В, объединяющего большую и малую петли, и C, включающего одну большую петлю) и считает, что трёхмерная структура всех субдоменов сходна. Учитывая данные о высоком проценте α-спиральности молекулы (50 – 60%) альбумина J. Brown пришёл к выводу, что каждая большая петля состоит из двух спиральных участков, соединённых с одной стороны дисульфидным мостиком, а с другой – полипептидным фрагментом. Спирали, начинаясь от цистина, закручиваются антипараллельно и образуют 6 туров, в которые вовлекаются 20 аминокислотных остатков. Расчётная длина каждого спирального участка равна 3,2 нм, а с учётом промежутков на каждом конце длина петли соответствует примерно 4 нм [7].
Остаток пролина, не имеющий водородной связи на атоме азота, ослабляет спиральную структуру, вызывая изгибы в полипептидной цепи. Сходные спирали, состоящие из 6 туров, образуются полипептидами, заключёнными между большой и малой двойными петлями (малый соединительный фрагмент), с вовлечением первой полупетли малой двойной петли. В этих участках также отсутствуют остатки пролина.
Исходя из изложенного выше, каждый домен сывороточного альбумина имеет пять спиралей: по две в каждой из больших петель и одну в малом соединительном сегменте. Предполагается наиболее вероятным, что эти спирали расположены в виде пятигранной призмы. При этом в пространство, ограниченное пятью спиралями, обращены гидрофобные группировки. У одного из оснований призмы сосредоточены концы спиралей, фиксированные дисульфидными связями. Противоположная сторона пятигранника остаётся открытой. Анализ аминокислотной последовательности свидетельствует о накоплении положительных зарядов в этих участках [8].
В настоящее время с полной определённостью доказано, что стабильность вторичной, третичной и четвертичной структур белка, определяется прежде всего физико-химическими взаимодействиями, обеспечивающими максимальный контакт изологических субъединиц. Было установлено, что укладка полипептидных цепей в нативную структуру начинается с формирования специфического «кармана», стабилизированного гидрофобными взаимодействиями, которые доминируют над остальными нековалентными связями. Кроме того, локальные гидрофобные взаимодействия стабилизируют вторичную структуру белка, что особенно важно для сохранения постоянства свойств описанных спиральных участков [9].
Описанная геометрическая чёткость расположения полипептидных цепей в пространстве – лишь одна из сторон структурной организации альбумина. Второй не менее важной чертой является высокая стерическая лабильность, лёгкость конформационных переходов.
Изложенная модель Брауна, а точнее – Брауна – Лозатти, допускает известную степень свободы. Это проявляется, во-первых, наличием вращательной подвижности доменов по отношению друг к другу; во-вторых, достаточно свободным положением аглобулярных цепей относительно глобулярного центра домена; в-третьих, возможностью объединения спиральных участков цепи не только в пятичленные сочетания, но и в трёхспиральные субъединицы. Последнее допущение связано с наличием двух дополнительных аминокислотных участков в положениях 176 – 197 и 368 – 389 (рисунок 1), объединяющих между собой три домена и состоящих из шести туров α-спирали, как и все другие спиральные образования альбумина. Вступая во временный контакт с двумя спиралями больших петель смежных доменов, эти участки могут формировать трёхгранные образования, имеющие строение, сходное с описанными пятигранниками.
Современные методы исследования пространственной структуры белка позволили построить трёхмерную модель структуры альбумина, которая представлена на рисунке 4 [10].

Рисунок 4 – Трехмерная структура альбумина
1.2.1 Гетерогенность альбумина
Гетерогенность альбумина разделяют на макрогетерогенность и микрогетерогенность. Первая связана с тем, что в организме молекулы альбумина не все являются идентичными и отличаются по сульфгидрильной группе, которая у одних молекул восстановлена, у других нет, по отношению к химическим реагентам. Также в пуле альбумина может наблюдаться наличие мономерной фракции, димерной и т.д., что также является фактором, обуславливающим гетерогенность молекулы альбумина .
Первоначально учёным удалось выделить две группы молекул альбумина: меркаптоальбумин и немеркаптоальбумин. Первый имеет восстановленные высокореакционноспособные SH-группы, во втором эти группы не были выявлены. L. Anderson (1966) выделил даже несколько форм меркаптоальбумина, по-разному чувствительных к действию β-меркапто -этанола. Во многих отношениях эти две формы альбумина идентичны [11].
Ещё одна форма гетерогенности альбумина связана с наличием наряду с мономерными олигомерных (дву-, три- и тетрамерных) форм, которые можно выявить методом электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия. Наиболее возможный путь димеризации заключается в формировании дисульфидных мостов в результате окисления SH-групп меркаптоальбумина. Было установлено, что такой процесс легко протекает в кислой среде при наличии катионов меди. Ионы других металлов (Zn2+
, Cd2+
, Co2+
, Ni2+
,Cr3+
) не катализируют реакцию полимеризации [12].
Однако механизм образования олигомерных фракций полностью не выяснен. Димеры альбумина в свою очередь гетерогенны. Только 30 % фракции димера переходит в мономер при восстановлении SH-групп β-меркаптоэтанолом в нейтральной среде. Часть димерных фракции расщепляется при низких значениях pH в присутствии диоксана, додецилсульфата натрия или гуанидингидрохлорида. Примеси димера и других олигомеров в препарате альбумина в количественном отношении широко варьируют: от 5 – 10 до 50% и более. Количественное соотношение олигомеров в препарате альбумина во многом зависит от способа выделения, продолжительности и условий хранения материала.
Термин «микрогетерогенность» был введён J. Colvin и соавт. (1954) [13], считавшими, что каждый белок является популяцией молекул. Развивая это понятие применительно к альбумину, J. Foster и соавт (1965) [14] выявили, что между перечисленными выше разновидностями белка существует множество промежуточных переходных форм (N – F).
Микрогетерогенность альбумина проявляется также при изучении тепловой денатурации белка.
О гетерогенности альбумина также свидетельствуют опыты по обратимой химической денатурации, т.е. различные представители популяций молекул альбумина неодинаково реагируют на присутствие в среде денатурирующего агента.
Проявлением микрогетерогенности альбумина является также неодинаковая растворимость отдельных его субфракций при специально подобранных условиях.
Степень гетерогенности и микрогетерогенности, присущей альбумину в норме, возрастает при различных патологических состояниях [15].
Особое место в происхождении полиморфизма отводится единственной меркаптильной группе, принадлежащей, как было отмечено выше, Cys34
. По одним данным эта группа участвует в образовании полимерных форм альбумина. По мнению J.Foster (1977), SH-группа свободно перемещаясь на N-концевой цепи над поверхностью глобулы альбумина, катализирует изомерные превращения дисульфидных мостиков.
Несмотря на сравнительно большое количество дисульфидных сшивок, молекула ЧСА не является совершенно жесткой глобулой. В ней есть достаточно много гибких и подвижных мест. Под действием температуры, рН и даже в результате связывания органических молекул в отдельных местах ЧСА происходят конформационные перестройки. Если изменять величину рН раствора альбумина от 7.4 в кислую сторону, то в области рН 4-5 происходит так называемый переход N-F. Этот переход изменяет общую конформацию глобулы и поэтому регистрируется многими методами. Другой переход происходит при увеличении рН до 7-9 (переход N-B). Он почти не влияет на размеры глобулы ЧСА и был зарегистрирован лишь в результате использования красителей, связывающихся с альбумином [16].
1.2.2 Характеристика центров связывания лигандов в альбумине
Многие аминокислотные остатки в молекуле альбумина имеют гидрофобную цепочку. В белковой глобуле такие остатки группируются друг с другом, скрываясь от молекул окружающей воды. Образующиеся гидрофобные кластеры, и некоторые из них способны, в свою очередь, связывать плавающие в воде небольшие гидрофобные молекулы – например, бутан, гептан и более длинные углеводороды.
Подобные же гидрофобные углеводородные цепочки содержатся и в молекулах жирных кислот, которые всегда есть в крови. Однако жирная кислота содержит не только гидрофобную цепочку, но и гидрофильную группу СОО-
. Поэтому такая кислота может "спрятать" свою гидрофобную цепочку в гидрофобный кластер альбумина только в том случае, если кластер находится около поверхности глобулы, чтобы группа СОО-
оставалась в полярном окружении (в воде). Таким образом, для жирных кислот и других молекул, имеющих гидрофильные группы, годится далеко не каждый кластер альбумина.
Группа СОО-
жирной кислоты при физиологических рН несет отрицательный заряд. Поэтому жирная кислота особенно хорошо должна связываться с такими участками глобулы альбумина, где имеется кластер гидрофобных аминокислот. Этот кластер находится около поверхности глобулы, а на поверхности расположены положительно заряженные аминогруппы остатков лизина или аргинина. В настоящее время установлено расположение четырех таких участков. Они обозначаются как ЖК-1, 2, 3, 4.
Молекула билирубина по размеру и по гидрофобности значительно превосходит молекулу жирной кислоты. Связывание билирубина — одна из самых важных функций альбумина. Имеется один участок в глобуле альбумина, связывающий билирубин наиболее прочно. Кроме того, альбуминовая молекула может связывать еще несколько молекул билирубина, но слабее [17].
Множество различных органических молекул, имеющих гидрофобные группы, способны связываться с альбумином. Одни из них по своей структуре лучше соответствуют одним участкам альбумина, другие – другим. Для количественной характеристики такого соответствия обычно используются два параметра: концентрация участков (центров) связывания (N) и константа связывания (К). Если в раствор альбумина добавить органические молекулы, способные с ним связываться (лиганды), то часть этих молекул свяжется с альбумином, а часть останется в растворе. Жирные кислоты, билирубин, многие метаболиты взаимодействуют с альбумином обратимо: связавшись с альбумином, такая молекула какое-то время находится в центре связывания, а затем уходит из альбумина. Обычно время пребывания молекулы в альбуминовом центре – менее 1 секунды [18].
Жирные кислоты и билирубин имеют самые высокие значения константы связывания. Другие лиганды, в том числе лекарства и зонды, характеризуются величинами pК, ниже 107
М-1
.
Молекула органического лиганда, связавшись с глобулой альбумина, изменяет свойства глобулы. Это регистрируется самыми разными методами, и можно только удивляться, как маленькая молекула массой около 300 Да может так повлиять на глобулу массой около 66.000 Да. В частности, изменяются свойства оставшихся незанятыми центров связывания. В результате константа связывания одного центра зависит от того, какие лиганды связаны с другими центрами той же молекулы альбумина. Эти влияния центров друг на друга называются кооперативностью связывания лигандов. Поэтому нет возможности точно описать свойства центров, и, как правило, данные разных авторов заметно различаются в зависимости от условий проведения опытов и подбора лигандов.
Глобула альбумина столь лабильна и чувствительна даже к слабым влияниям потому, что каждый центр связывания лигандов формируется благодаря контакту разных участков полипептидной цепи, которые притягиваются друг к другу сравнительно слабыми гидрофобными и Ван-Дер-Ваальсовыми взаимодействиями. Такое взаимодействие легко нарушить, что сразу отразится на большом участке глобулы.
Наиболее удобным средством исследования центров связывания в молекуле альбумина оказалось использование красителей. Вначале это были абсорбционные красители, но затем все большую роль стали играть флуоресцентные красители – так называемые флуоресцентные зонды. В дальнейшем стали применять, кроме того, спиновые зонды.
Самыми чувствительными индикаторами структурных перестроек в альбумине являются флуоресцентные зонды [19].
1.3 Влияние физическо-химических факторов на альбумин
Как было сказано выше, микрогетерогенность альбумина также проявляется при изучении тепловой денатурации белка. S. Stokrova и Sponar установили, что молекулы ЧСА по-разному чувствительны к нагреванию. Боле подробное исследование этого процесса показало, что часть молекул альбумина под влиянием тепла переходит в особое конформационное состояние, приобретая при этом способность защищать исходный нативный альбумин от температурного воздействия. Полагается, что образование такой формы является эволюционным регуляторно-защитным механизмом [20].
Влияние рН.
Как известно, почти все глобулярные белки подвергаются, воздействию ионов водорода при их достаточных концентрациях. При этом белки могут увеличиваться в объеме, диссоциировать, быстро денатурировать при рК, не очень удаленных от изотонической точки, а некоторые белки, в зависимости от рН среды, могут существовать в нескольких изомерных формах. В этом случае другие изомерные формы не обязательно более беспорядочны, чем структура нативной молекулы. Как правило, все изомерные формы обладают хорошо определенной структурой. Альбумин относится к числу последних белков.
Было установлено, что при снижении величины рН (<3,5), происходит увеличение объема молекул БСА. Это объясняется явлением кулоновского отталкивания в результате аномального связывания H+
и накоплением положительных зарядов в кислой среде. В 0,15 М растворе NaCl кислотные изменения значительно менее выражаются, хотя в этом случае незначительно увеличивается объём молекулы. При более высоких значениях рН (=4,0) в 0,15 М растворе NaCl происходит некоторое повышение вязкости раствора. Такое состояние было названо экспендабельным, т.е. способным к расширению [21].
Учёные M. Sogami и J. Foster разделили кислотные переходы альбумина на ряд стадий: N → F`→ F → E (N – normal, F – fast, эта форма быстрее перемещается в геле сефадекса, E – expanded, т.е. полностью растворённая). Причём стадии N → F`→ F являются обратимыми, а E – необратимой. Разделение стадий на F`и F обусловлено неравномерным ходом процесса деспирализации альбумина в кислой среде. В интервале величин pH от 4,5 до 3,7 α-спиральность снижается с 51 до 44%. Затем процесс несколько стабилизируется (F`). Далее при pH 3,5 – 3,0 спиральность вновь снижается до 35%. При этом изменяются физико-химические свойства альбумина. В частности, F-форма значительно хуже растворима в концентрированных солевых растворах, чем N-форма. Предполагается, что N – F переход сопровождается раскрытием гидрофильных поверхностей белковой молекулы. Затем (F`→ F) – открываются гидрофобные зоны [21].
По данным A. Wishnia и T. Pinder, F-форма лишена присущей N-альбумину способности связывать алканы. Это положение свидельствует о необходимости наличия гидрофобных щелей в молекуле, в которые внедряется молекула лиганда. Исчезновение таких полостей при раскрытии гидрофобных зон в F-форме приводит к утрате способности белка фиксировать неполярные молекулы.
В диапазоне pH 6,0 – 8,0 конформационные переходы альбумина сопровождаются изменением микроокружения аминокислотных остатков триптофана, тирозина, гистидина. При повышении рН в дипазоне от 7,0 до 9,0 происходит N – B-переход молекулы, в результате которого повышается доступность имидазольных остатков, скрытых в N-форме. N – B-переход молекулы альбумина сопровождает pH-зависимое связывание нестероидных противовоспалительных соединений, гепариновых антикоагулянтов и т.д. [22].
1.3.1
Влияние химических модификаторов на альбумин
В организме молекула альбумина подвергается различным модификациям, причем при патологии степень модификации может возрастать во много раз. Один из видов модификации - это присоединение остатка глюкозы к лизину (No 525). У здоровых людей гликозилировано в среднем несколько процентов всех молекул альбумина, а при диабете – в 1.6 – 2.5 раза больше[23].
Одним из химических веществ, наиболее часто применяемых для денатурации глобулярных белков, является водный раствор мочевины. До сих пор точно не установлены ни механизм ее денатурирующего действия, ни природа происходящих в белке изменений структуры. В течение длительного времени денатурирующее влияние мочевины связывали с ее способностью образовывать водородные связи, разрывающие внутримолекулярные водородные связи белка. Однако после, было доказано, что метилзамещенная мочевина или аммониевые соли являются менее эффективными денатурантами, несмотря на их большую способность образовывать водородные связи. К тому же, если это было бы так, то сама вода могла бы быть эффективным денатурирующим агентом [24].
Другие исследования, имеющие дело с растворимостью углеводородов и аминокислот в растворах мочевины, показали, что влияние мочевины на нативную конформацию белков может быть приписано дестабилизации гидрофобных связей. Это объясняется тем, что в растворах мочевины неполярные группы находятся в энергетически менее выгодном окружении, чем в чисто водных растворах. Денатурирующее действие мочевины было объяснено также на основе предположения о ее катионной структуре. Наиболее удивительным в денатурирующем действии мочевины является то, что, несмотря на полное, как считается, разрушение пространственной структуры, белки в водной среде остаются в растворенном состоянии [25].
Для большинства белков действие мочевины проявляется в увеличении размеров молекулы и уменьшении количества вторичных структур: α-спиралей и β-структур. В этом плане сывороточный альбумин не является исключением. Обнаружено, что в растворе мочевины при концентрации больше, чем 2 М, увеличивается левовращение и вязкость сывороточного альбумина. При этом также изменяется УФ-спектр поглощения белка. Все эти изменения обратимы, что объясняется специфическим расположением дисульфидных мостиков в молекуле альбумина. Левовращение молекулы альбумина в растворе 8М мочевины (при 20°С, рН 6,3) увеличивается, степень β-спиральности ее уменьшается почти в 2 раза, что указывает на обширную дезорганизацию нативной структуры белка. При этом характеристическая вязкость этого белка возрастает от 4 до 17 мл/г, а радиус инерции молекулы, определенный рентгенографическим исследованием увеличивается. Кроме того, если в нативной форме альбумин имеет 3 доступных к восстановлению дисульфидных мостика, то в присутствии 8М мочевины восстанавливаются все дисульфидные связи [26].
Интересно отметить, что рН системы сильно влияет на процесс денатурации. Например, характеристическая вязкость альбумина в растворе 8 М мочевины при рН 10,5 почти в 2 раза больше, чем при рН 7,4. Кроме того величина изменения как оптического вращения так и характеристической вязкости зависит от температуры системы. В интервале температур от 0 до 40°С с повышением температуры действие, вызванное мочевиной подавляется. Ниже 40°С процесс денатурации является обратимым по отношению к изменению температуры и концентрации мочевины, выше 40°С происходит необратимое изменение структуры альбумина которое имеет место при более низких концентрациях денатуранта [27].
Несмотря на явное увеличение размеров молекулы, о чем свидетельствует четырехкратное увеличение вязкости и доступности его дисульфидных связей в растворах 8 М мочевины, белок не только остается в растворенном состоянии, но, как показали Ноли, Маркус и другие, не образует агрегированных форм. Последнее указывает, что тиольная группа, которая, как правило, является причиной образования агрегатов в растворах альбумина, остается стерически недоступной, как и в нативной форме. Из этого следует, что полного разрушения структуры молекулы альбумина в 8М мочевине не происходит, о чем свидетельствует также и сохранение половины спиралей и небольшое, всего в 1,5 раза, увеличение радиуса инерции молекулы [28].
Обобщая все эти факты, можно прийти к выводу, что природа изменений структуры альбумина в растворе 8 М мочевины до последнего времени остается непонятной. Однако использование метода тритиевой метки, позволяющего определить, как качественно, так и количественно, доступную поверхность молекулы белка в разных его структурных состояниях, дало более детальную информацию о происходящих изменениях в структуре альбумина при его взаимодействии с 8 М мочевиной[29].
В растворе 8 М мочевины распределение тритиевой метки по аминокислотным остаткам ЧСА отличается от нативного, обнаруживая увеличение доступности (радиоактивности) для целого ряда остатков: Arg, Asp, Ser, Pro, Gly, Ala. Наибольшими изменениями, почти в 3 раза, характеризуются остатки Arg и Gly [30].
Увеличение доступности аминокислотных остатков вполне закономерное явление, которого и следовало ожидать исходя из совокупности данных по влиянию мочевины. Известно, что даже в тех случаях, когда в присутствии 8 М мочевины не наблюдается заметных изменений структуры, фиксируемых по вязкости и оптическому вращению. Наблюдается, однако, уменьшение доступности дисульдных связей для меркаптэтанола[31].
Применяя предлагаемый подход определения площади полной доступной поверхности белковой глобулы, можно оценить и полную доступную поверхность молекулы ЧСА в растворе 8 М мочевины, которая составляет 57300 А [32].
Таким образом, в растворе 8М мочевины полная доступная поверхность молекулы ЧСА увеличивается от 39000 (для нативного белка) до 57300 А. Известно, что характер изменения доступности остатков и полной доступной поверхности молекулы ЧСА не согласуется с четырехкратным увеличением вязкости раствора этого белка в присутствии 8 М мочевины. В самом деле, хорошо видно, что, во-первых масштаб изменений не велик, так как полная доступная поверхность молекулы увеличивается всего в 1,45 раза, а, во-вторых, доступность гидрофобных остатков увеличивается очень не существенно, от 19900 А до 24900 А – всего в 1,25 раза, тогда как признаком развертывания является превалирующее экспонирование растворителю гидрофобных остатков[33].
Можно прийти к выводу, что увеличение вязкости может быть следствием изменения формы молекулы, то есть увеличение асимметрии, и появления у нее дополнительной гибкости в результате разрушения гидрофобных связей. Слабое увеличение доступности гидрофобных остатков связано, в первую очередь, с тем, что в нативной структуре, они уже обладают повышенной доступностью тритию [34].
Слабое влияние мочевины на структуру альбумина хорошо согласуется с фактом устойчивости этого белка к действию мочевины. Эта закономерность прослеживается даже при нагревании ЧСА до 44°С. Интересно, что в растворе 8 М мочевины совсем по другому ведут себя другие глобулярные белки. Например, по данным, полученным Волынской методом тритиевой метки, изменение структуры лизоцима в растворе 8М мочевины характеризуется четырехкратным увеличением суммарной доступности остатков. Причем гидрофобные остатки увеличивают доступность в 7 раз при неизменности гидрофильных остатков. Это означает, что характер конформационных изменений заметно различается для сывороточного альбумина и лизоцима. По-видимому, наблюдаемое влияние мочевины на структуру альбумина обусловлено особенностями пространственной организации этого белка. Действительно, было доказано, что альбумин, по сравнению с лизоцимом, характеризуется низким коэффициентом свернутости нативной структуры и высоким коэффициентом шероховатости поверхности. По физическому смыслу коэффициент свернутости означает долю поверхности всей полипептидной цепочки, которая исчезает при свертывании ее в нативную структуру. Коэффициент шероховатости дает представление о рельефности поверхности, и очевидно, что большая величина его свидетельствует о большой изрезанности поверхности, наличием на ней складок, щелей карманов и петель. Поэтому влияние мочевины, по всей вероятности, обусловлено именно наличием таких неплотно упакованных участков в структуре белка, которые, вероятно, локализованы в интерфазах субъединиц. Возможно, что действие мочевины в существенной степени сводится к расширению молекулы в результате разъединения субъединиц. Поэтому, в отличие от лизоцима, структурные изменения в альбумине могут носить локальный характер и не затрагивать в значительной степени всю молекулу. Другими словами, деформация молекулярной структуры при не очень больших воздействиях может локализоваться именно в неплотноупакованных участках, не приводя к полному разрушению структуры [35].
Значительным структурным изменениям молекулы альбумина, по-видимому, препятствует также система S-S-связей, расположенных регулярно по всей длине полипептидной цепи[36].
Надо отметить, что на денатурацию альбумина мочевиной немало влияют и связанные с ним жирные кислоты, Катцом и Денисом показано, что обезжиренный альбумин более чувствителен к такой денатурации, чем альбумин с жирными кислотами. Удаление жирных кислот увеличивает скорость денатурации так, если денатурация обезжиренного белка происходит в течение 5-10 минут, то для необработанного альбумина требуется 30-60 минут. По-видимому, присутствие в альбумине жирных кислот не только уменьшает скорость денатурации белка при его взаимодействии с 8М мочевиной, но и препятствует обширным изменениям его структуры [37].
1.4
Функции сывороточного альбумина в организме человека
Альбумин выполняет в организме ряд функций, имеющих важное значение для поддержания гомеостаза внутренней среды всего организма. Их нарушение играет роль в патогенезе многих критических состояний организма человека[38].
В норме молекулы альбумина обеспечивают 80—85% онкотического давления плазмы. Это происходит вследствие низкой проницаемости стенки капилляров для молекул альбумина, которые содержат большое количество заряженных групп, удерживающих воду. При критических состояниях проницаемость эндотелия увеличивается и онкотическое давление падает. При некоторых заболеваниях (острой почечной недостаточности, циррозе печени, гемолитической болезни и др.) способность молекул альбумина связывать воду изменяется, и это тоже может оказывать влияние на онкотическое давление плазмы [39].
Альбумин обратимо связывает и переносит самые разнообразные низкомолекулярные вещества: метаболиты, среди которых жирные кислоты, желчные пигменты, окись азота, холестерин, металлы как постоянной (Zn2+
, Са2+
), так и переменной валентности (Cu2+
, Fe2+
, Ni2+
) и очень многие лекарственные препараты. Тестирование лекарственных средств на способность связываться с ЧСА является обязательным этапом их испытаний. Число молекул лигандов, которое одновременно способна удержать 1 молекула ЧСА, может достигать 10 и более. Примерно 1 литр плазмы может нековалентно связать более 1 грамма лиганда с молекулярной массой около 350 Да [40].
При связывании лиганда с альбумином химическая и биологическая активность этого лиганда меняется. Многие вещества, токсичные в свободной, несвязанной форме значительно снижают свою активность при связывании с ЧСА. Это особенно важно при таких состояниях, как полиорганная недостаточность, циркуляторный шок и т. п., когда функции печени и почек ослаблены. Таким образом, альбумин является важным звеном в системе детоксикации организма.
Многие метаболиты и лекарственные вещества конкурируют между собой за связывающие центры альбумина. Это обстоятельство должно обязательно учитываться при назначении лекарственных препаратов. Например, цефалоспорины и другие антибиотики, связываясь с ЧСА, могут вытеснять из него билирубин [41].
Альбумин вносит значительный вклад в защиту организма от вредного действия свободных радикалов. Это достигается, в частности, связыванием металлов переменной валентности, в результате чего их радикалобразуюшая активность уменьшается в 100 раз. Кроме этого, в норме молекула альбумина содержит одну восстановленную SH-группy, которая легко окисляется, в том числе в реакциях с участием радикалов. При этом происходит образование стабильных продуктов (дисульфидов, сульфоновых кислот). Это значит, что альбумин, таким образом, обрывает цепи радикального окисления и является антиоксидантом крови. В физиологических условиях около 80% всех определяемых тиолов плазмы составляют именно тиолы альбумина. Некоторые исследователи частично связывают положительный эффект препаратов альбумина при критических состояниях с увеличением концентрации активных тиолов. По-видимому, и другие группы альбумина (например, аминогруппы) способны вступать в реакции с радикалами, защищая организм от окислительного стресса [42].
Считается, что альбумин вовлечен в реакции с участием радикалов не специфически, а в силу того, что его концентрация во внеклеточном пространстве относительно высока, и обмен его происходит относительно быстро, около 20 дней.
1.5 Основное понятия и закономерности люминесценции
Понятие «люминесценция» относят не к отдельным атомам и молекулам, а к совокупностям – телам и тесно связывают с температурой среды. Последнее вызвано тем, что элементарные акты возбуждения молекул и испускания света могут быть одинаковыми в случае теплового излучения и люминесценции. Согласно определению Видемана – Вавилова, люминесценция – излучение, представляющее собой избыток над тепловым излучением тела при данной температуре и имеющее длительность, существенно превышающую период световых колебаний.
В зависимости от источников, вызвающих люминесценцию, она подразделяется на триболюминесценцию (механические деформации некоторых кристаллов), фотолюминесценцию (свет), сонолюминесценцию (звуковая волна – индуктор) и т.д.
Спектральная область, обычно используемая для люминесценных измерений (200 – 800 нм), соответствует электронным переходам в молекуле. Поглощение молекулой кванта света в этой области спектра приводит к переходу электрона на более высокий энергетический уровень.
Поглощение света происходит за время порядка 10-15
с, что соответствует периоду колебаний световой волны. В течение этого времени заметно не изменяются ни положение, ни импульсы тяжёлых ядер и их относительное движение до акта поглощения света и сразу же после него одинаковы.
Молекула, попавшая на верхние колебательные уровни любого возбуждённого состояния, быстро теряет избыток колебательной энергии при столкновениях с окружающими молекулами. Это процесс колебательной релаксации. Безызлучательный переход между электронными состояниями одинаковой мультиплетности называется внутренней конверсией, аналогичный переход между состояниями разной мультиплетности – интеркомбинационной конверсией [43].
Возможны два вида интеркомбинационной конверсии: переход S1
– S0
(из первого синглетного возбуждённого в основное синглетное состояние соответственно) и переход Т1
– S0
(из триплетного возбуждённого состояния в основное синглетное соответственно).
Помимо безызлучательного перехода молекула может попасть в основное состояние S0
, испустив квант люминесценции. Если квант был испущен с синглетного возбуждённого состояния, то такой тип люминесценции называется флуоресценцией
. Длительность флуоресценции составляет 10-8
– 10-10
сек.
Возможен и другой путь излучательной релаксации возбуждённого синглетного состояния. Возбуждённая молекула может безызлучательным путём перейти в триплетный возбуждённый уровень с изменением спина.
Это так называемое метастабильное состояние. Перейдя на триплетный уровень, молекула может находится на нём примерно 10-3
– 10 сек, что значительно больше длительности синглетного возбуждённого состояния.
Свечение, испускаемое при переходе молекулы с триплетного в основное синглетное состояние называется фосфоресценцией
.
Оcновными характеристиками, параметрами люминесценции являются её спектры, выход, длительность и поляризация [44].
Люминесценция белков может быть обусловлена аминокислотами, входящими в состав полипепидной цепи макромолекул (собственная люминесценция), простетическими группами (восстановленные пиридиннуклеотиды и флавины), связанными с белковой частью сложных ферментов, химически присоединённой меткой (красителем), а также зондом (зондовая люминесценция).
Собственная
люминесценция белка обусловлена в целом аминокислотами, имеющими ароматический радикал, а значит это – триптофан, тирозин, гистидин, фенилаланин. Поглощение этих аминокислот в области 250 – 300 нм довольно значительно. Из других аминокислот только цистин имеет в этой области поглощение, сравнимое с фенилаланином ε240 = 300
, ε290 = 50
; пептидная связь поглощает в области более 220 нм. Включение ароматических аминокислот в состав белков сопровождается небольшим сдвигом в «красную» сторону, при этом слегка увеличивается поглощательная способность хромофоров[45].
1.6 Применение зондовой флуоресценции для изучения структуры молекулы альбумина
Способность красителей связываться с белками и реагировать на состояние белковой молекулы, известна давно. Вначале о структуре и поведении альбумина судили по изменению поглощения света красителем. И.Клотц использовал метиловый оранжевый и другие красители – органические кислоты с отрицательным зарядом [46]. По мере появления приборов, регистрирующих слабые потоки люминесцентного света, стали использоваться и флуоресцирующие красители. Флуоресценция обычно гораздо более чувствительна к изменениям окружения молекулы красителя, чем поглощение света, и имеет ряд других особенностей, благодаря которым флуоресцентные красители приобрели большое распространение в исследовании белков.
Краситель может быть связан с белком ковалентно (необратимо) благодаря химической связи или нековалентно (обратимо) благодаря слабым гидрофобным и электростатическим взаимодействиям. Первую группу обычно называют метками, вторую - зондами. В 1952 году Г.Вебер впервые применил флуоресцентную метку, а Д.Лоуренс - флуоресцентный зонд для изучения структуры альбумина. Выяснилось, что флуоресценция зондов очень чувствительна к перестройкам в глобуле альбумина, чего нельзя сказать, используя флуоресцентные метки. При связывании зонда АНС (1-амино-N-фенил-8-сульфонафталин) с альбумином интенсивность флуоресценции зонда возрастает в сотни раз. Это свойство АНС почти сразу же попытались использовать для клинического измерения концентрации альбумина в сыворотке. Однако оказалось, что при патологиях, особенно пригипербилирубинемии, флуоресценция АНС в альбумине снижается, что мешает количественному его определению.
Активное использование флуоресцентных зондов для изучения структуры белков началось с 1965 года. Наиболее подробно исследовано взаимодействие с альбумином отрицательно заряженного зонда АНС. Имеется около 5 центров связывания АНС на молекуле альбумина. При этом 2-3 из них имеют более высокую константу связывания. Попадая в эти центры, молекула АНС оказывается в жестком окружении, а ее гидрофобная группа полностью скрыта от молекул воды. Примерно 90% флуоресценции АНС происходит именно из этих центров. Остальные 2-3 центра связывают АНС слабее, и в них зонд частично доступен воде. При переходе от рН 7 к рН 4 (N-F переход) константа связывания АНС сильными центрами снижается, однако число таких центров возрастает, за счет чего интенсивность флуоресценции АНС тоже возрастает. При этом сильные центры по-прежнему хорошо скрывают молекулу АНС от воды.
Благодаря тому, что флуоресцируют практически только те молекулы АНС, которые связаны с альбумином, легко измерить параметры связывания зонда с белком.
Незаряженный зонд ДМХ (4-диметиламинохалкон) связывается с 2-3 центрами альбумина. Подобно АНС, он попадает в жесткое окружение, что видно по появлению кругового дихроизма, и скрыт от молекул воды, которые теперь уже не тушат флуоресценцию ДМХ. Молекула ДМХ контактирует в этих центрах с двумя остатками тирозина, расположенными вблизи поверхности белковой глобулы. Такой контакт с фенольными группами тирозина приводит к частичному тушению флоуресценции ДМХ. При переходе от рН 7 к рН 9 (переход N-B) эти два остатка тирозина ионизируются: один становится отрицательно заряженным к рН 9, второй – к рН=10,5. Остальные 16 остатков тирозина ионизируются только при рН > 10,5. В результате ионизации первых двух остатков тирозина они выходят на поверхность белка, оставив ДМХ в глубине центра связывания, их тушащее влияние уменьшается, и интенсивность флуоресценции ДМХ возрастает. В свою очередь, ДМХ перестает влиять на круговой дихроизм этих остатков тирозина. Дальнейшее увеличение рН приводит к разрыхлению белковой молекулы, что видно по снижению флуоресценции ДМХ, молекула которого становится теперь доступной тушащему действию молекул воды.
При снижении рН от 7 к 3 (переход N-F) центры связывания ДМХ разрыхляются, в них проникает вода и тушит флуоресценцию ДМХ.
Отрицательно заряженный зонд К-35 был предложен специально для исследования состояния альбумина в сыворотке крови при патологии. К-35 имеет величину константы связывания с альбумином примерно такую же, как АНС, около 1,8 ∙ 10 5
М-1
, и занимает два типа центров. Примерно 23% связанных молекул К-35 находится в центрах одного типа, где аминогруппа зонда хорошо скрыта от молекул воды. Остальные 77% молекул – в центрах другого типа, где зонд частично доступен воде.
В результате исследования конкуренции флуоресцентных зондов с метаболитами и лекарственными веществами за центры связывания были найдены зонды-маркеры различных центров альбумина: ДАНСил-амид (5-диметиламинонафталин-1-сульфониламид), ДАНСил-саркозин, ДАНСил-ундеканоевая кислота и производные 7-аминокумарина.
Если к раствору АНС добавить альбумин, то часть молекул зонда связывается с альбумином, и интенсивность их флуоресценции возрастает в десятки-сотни раз. Однако при добавлении билирубина интенсивность флуоресценции АНС снижается, по-видимому, в результате конкуренции между билирубином и АНС. Изучение этой смеси показало, что билирубин не влияет на константу связывания АНС с альбумином, а уменьшает число центров связывания.
Зонды конкурируют и с лекарственными веществами. Так, при добавлении ряда лекарственных веществ интенсивность флуоресценции АНС и К-35 в растворе альбумина снижается. При этом зонд К-35 гораздо чувствительнее к присутствию лекарств, чем АНС, хотя оба зонда имеют близкие величины констант связывания.
После многочисленных исследований конкуренции подобных соединений за центры альбумина сложились следующие, достаточно упрощенные представления. Считают, что для связывания билирубина, лекарственных соединений и флуоресцентных зондов наиболее важны два центра:
Центр I расположен во втором домене альбуминовой глобулы. В его формировании участвует область полипептидной цепи, включающей лизин-199 и триптофан-214. Маркерами этого центра являются фенилбутазон и незаряженный флуоресцентный зонд ДАНСил-амид.
Центр II формируется участком полипептидной цепи первого домена, включающим аргинин-145 и лизин-194. Его маркерами являются иопановая кислота и отрицательно заряженный флуоресцентный зонд ДАНСил-саркозин. Как следует, связывание отрицательно заряженного зонда К-35 с альбумином нарушается маркерами обоих типов центров. Возможно, К-35 связывается как с центром II, так и с центром I. Однако пример с билирубином, рассмотренный выше, показывает, что вполне возможны влияния одних центров на другие, а не только прямая конкуренция вещества и зонда за один и тот же центр [47].
Обобщая обзор литературы можно прийти к выводу о многообразии функции, выполняемых сывороточным альбумином. Такое многообразие определяется уникальным строением данного белка, причём структура сывороточного альбумина настолько лабильна, что даже при небольшом изменении интенсивности физико-химических факторов (рН, температура, денатурирующие агенты), в белке происходят конформационные изменения, которые в свою очередь отражаются на функциях, выполняемых альбумином.
Наличие таких остатков аминокислот как: тирозин, фенилаланин и, в особенности, триптофан позволяет с успехом использовать очень чувствительный физико-химический метод в аналитической химии – флуоресцентная спектроскопия.
Благодаря открытию зондов (красители, нековалентно связывающиеся с белком), например АНС, который в связанном с белком состоянии в сотни раз интенсивнее флуоресцирует, чем в несвязанном состоянии, стало возможным изучать очень тонкие конформационные изменения, происходящие в альбумине и в других белках в целом.
2 ОБЪЕКТ, ПРОГРАММА И МЕТОДИКА ИССЛЕДОВАНИЙ
Объектом исследования являлся бычий сывороточный альбумин фирмы «Sigma-Aldrich».
Целесообразность использования бычьего сывороточного альбумина для эксперимента по сравнению с человеческим сывороточным альбумином (ЧСА) заключается в том, что БСА имеет два триптофанила, а ЧСА только один, находящийся в поверхностной гидрофобной складке молекулы. Именно остатками триптофана обусловлен основной вклад в собственную флуоресценцию белка. Причём в молекуле БСА один триптофанил располагается там же, где и триптофанил молекулы ЧСА, а второй спрятан внутри белковой глобулы. Если произойдёт разворачивание белка под действием каких-либо факторов, то внутренний триптофанил выйдет на поверхность и флуоресценция белка увеличится. В случае ЧСА увеличение интенсивности флуоресценции наблюдаться не будет.
Полученные результаты исследований можно будет с полной достоверностью перенести на молекулу ЧСА, т.к. БСА и ЧСА отличаются друг от друга на три аминокислоты и положением некоторых аминокислот, а в целом структуры этих молекул идентичны.
Предметом исследования явилось изучение конформационных изменений и структуры альбумина в целом под действием физико-химических факторов.
Программа исследований включала постановку и решение следующих задач:
1) Исследование влияния сверхвысоких концентраций мочевины на конформацию альбумина.
2) Влияние свободных радикалов на структуру молекулы альбумина.
Методом изучения влияния сверхвысоких концентраций мочевины на конформацию альбумина была флуоресцентная спектроскопия. Параметр флуоресценции – интенсивность.
Влияние свободных радикалов на структуру молекулы альбумина исследовали методом гель-хроматографии и флуоресцентной спектроскопии.
Флюоресценцию растворов альбумина исследовали на спектрофлюориметре марки СМ 2203 Solar. Хроматографию проводили на TSK - HW55 геле.
Эксцинцию для собственной флуоресценции устанавливали на приборе при длине волны 280, 290 нм, эмиссию при длине волны 350 нм. Эксцинцию зондовой флуоресценции устанавливали при длинах волн 280, 290 и 320 нм, эмиссию – при длине волны 450 нм.
Для изучения влияния различных концентраций мочевины на бычий сывороточный альбумин (БСА) использовали концентрацию белка, равную 0,66 г/л, или 10-5
моль/л. Раствор БСА с концентрацией 10-5
М готовили в трёх различных буферах: фосфатный с рН 7,43, трис-буфер с рН 9,08, ацетатный с рН 4,54.
Концентрация БСА 0,66 г/л является оптимальной, т.е. флуоресценция белка связана прямопропорциональной зависимостью с концентрацией. Для нахождения данной концентрации строили калибровочный график зависимости интенсивности флуоресценции от концентрации белка. На графике нашли отрезок, характеризующийся прямой пропорциональной зависимостью и, поделив его пополам, получили значение оптимальной концентрации альбумина.
Концентрации мочевины, которые брались для эксперемента находились в интервале от 2М до 10 М.
По сравнению с соотношением содержания мочевины и сывороточного альбумина в организме (10 : 1), для эксперимента бралось соотношение мочевины и альбумина 1 : 10000 – 100000 соответственно. Т.о концентрации мочевины были взяты сверхвысокими.
Помимо собственной флуоресценции белка исследовали зондовую флуоресценцию. В качестве зонда использовали N-фенил-1-амино-8-сульфонафталин (АНС). Количество зонда, необходимое для реакции с альбумином, рассчитывали, исходя из того, что количество центров на альбумине для АНС равно 5 – 6. Поэтому концентрация зонда для эксперимента взяли равной 6 ∙ 10-5
моль/л.
Данные зондовой флуоресценции наряду с данными о собственной флуоресценции могут дать дополнительную информацию о конформационных изменениях белка, т.к. зондовая флуоресценция более чувствительна к изменениям структуры белка, чем собственная флуоресценция.
Для изучения влияния свободных радикалов на бычий сывороточный альбумин использовали концентрацию альбумина 10-5
М. Раствор альбумина готовили в фосфатном буфере (Na2
HPO4
– KH2
PO4
(1/15M)) с pH = 7,44, близкой к физиологическому значению рН крови. Свободные радикалы образовывались в растворе альбумина, по реакции Вейсса – Габера, путём добавления смеси пероксида водорода с концентрацией 3 · 10-5
и сульфата меди (II) – 12 · 10-4
моль/л.
В организме человека пероксид водорода и ионы меди (II) содержатся в количествах, примерно равных 30 мкмоль/л и 12 мкмоль/л соответственно.
При таком соотношении в организме свободные радикалы практически не образуются, но при повышении концентрации ионов меди (II), вследствие их чрезмерного поступления извне, происходит образование свободных радикалов.
Ход эксперимента выглядел следующим образом. Вначале приготовили контрольную пробу, в которой находился раствор альбумина с пероксидом водорода. Затем готовили опытный раствор альбумина, куда добавили вначале пероксид водорода, а затем, через два часа – сульфат меди (II) выше указанных концентраций соответственно. Далее опытную и контрольную пробу нанесли на колонки, содержащие гель TSK - HW55. Собрали элюированные пробы и получили профиль элюции белков.
Затем проводили флуоресцентную спектроскопию белковых фракций, полученных элюированием сывороточного альбумина на геле TSK - HW55. Длина волны возбуждения ультрафиолетом (УФ) равнялась 290 нм, длина волны регистрации флуоресценции составляла 350 нм.
При возбуждении фракции УФ излучением с длиной волны 290 нм, наблюдали появление, кроме полосы флуоресценции триптофана, дополнительной полосы флуоресценции, обладающей длинноволновым сдвигом, с длиной волны 360 нм.
Предположили, что данная полоса свечения возникла при окислении свободными радикалами остатка гистидина. Для проверки данной гипотезы провели химическую реакцию взаимодействия гистидина со смесью сульфата меди и пероксида водорода. Затем производили измерение флуоресценции продуктов окисления гистидина. Максимум интенсивности флуоресценции соответствовал длине волны 360 нм, что совпадало с флуоресцентной характеристикой дополнительной полосы свечения белковой фракции.
Далее изменили длину волны возбуждения на 360 нм и регистрировали появление нового спектра флуоресцеции, обладающего длинноволновым сдвигом.
3 РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
3.1 Исследование влияния сверхвысоких концентраций мочевины на конформацию альбумина
В результате проведения эксперимента были получены результаты, отражённые на графиках зависимости интенсивности собственной и зондовой флуоресценции от концентрации мочевины и длины волны возбуждающего и эмиссионного света.
Для интерпретации полученных результатов нужно упомянуть, что в нативном белке в водных растворах собственная флуоресценция, обусловленная триптофаном наблюдается при значении длины волны экстинции на 280 -290 нм и при значении длины волны эмиссии на 350 нм. Экстинция зонда АНС = 320 нм, а эмиссия – 450 нм.
Из полученных результатов следует, что сверхвысокие концентрации мочевины уменьшают интенсивность как собственной так и зондовой флуоресценции. Это можно увидеть на рисунках 5 – 10. Однако падение интенсивности собственной флуоресценции белка носит сложный нелинейный характер. В некоторых участках кривых отмечается даже незначительное увеличение интенсивности. Например, на графиках зависимости собственной флуоресценции от концентрации мочевины показано, что в кислой среде (рисунок 5) незначительное увеличение интенсивности флуоресценции (ИФ) наблюдается при концентрации мочевины 8М, в нейтральной среде (рисунок 6) – при концентрации мочевины 5М и 8М, в щелочной среде (рисунок 7) – при концентрации мочевины 5М. Очевидно, что результатом такого характера кривых является различие в рН среды, в которой находится альбумин, что подтверждает литературные данные о существовании разных форм альбумина в зависимости от рН среды.
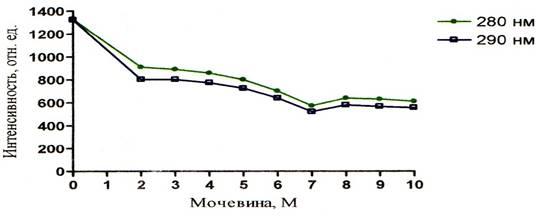
Рисунок 5 – График зависимости интенсивности флуоресценции альбумина от концентрации мочевины при рН = 4,54
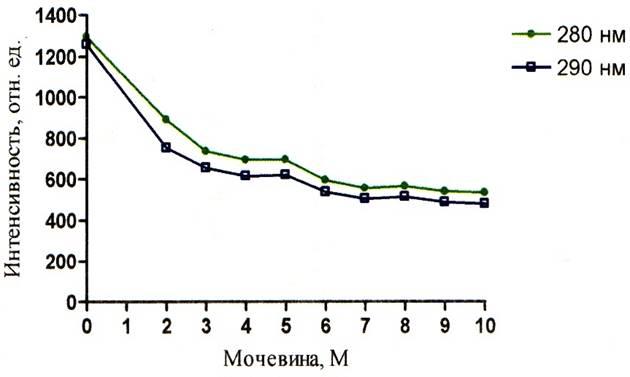
Рисунок 6 – График зависимости интенсивности флуоресценции альбумина от концентрации мочевины при рН = 7,43
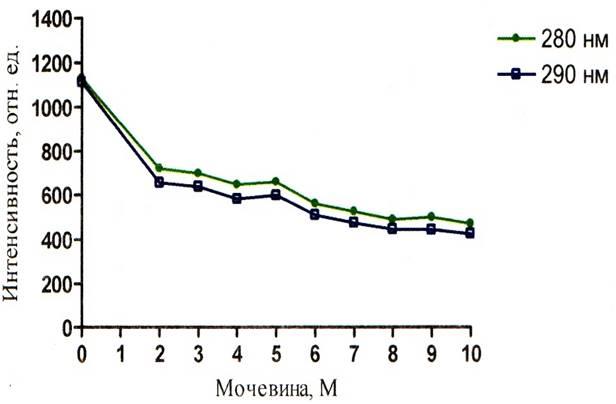
Рисунок 7 – График зависимости интенсивности флуоресценции альбумина от концентрации мочевины при рН = 9,08
Падение интенсивности зондовой флуоресценции (рисунки 8, 9, 10) при возбуждении длиной волны 320 нм имеет более спокойный и плавный характер.
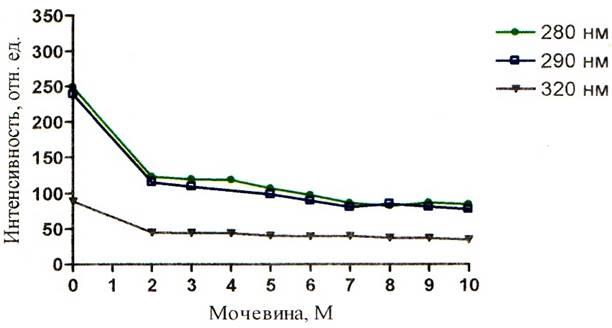
Рисунок 8 – График зависимости интенсивности зондовой флуоресценции альбумина от концентрации мочевины при рН = 4,54
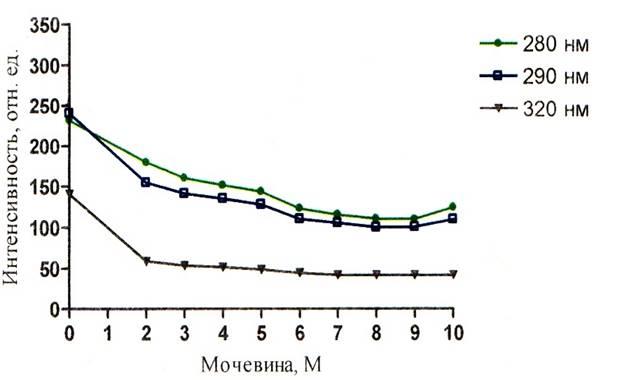
Рисунок 9 – График зависимости интенсивности флуоресценции альбумина от концентрации мочевины при рН = 7,43
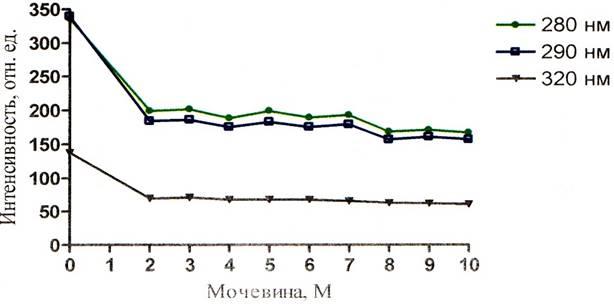
Рисунок 10 – График зависимости интенсивности флуоресценции альбумина от концентрации мочевины при рН = 9,08
Обычно зондовая флуоресценция альбумина, обусловленная зондом АНС, появляется при возбуждающем свете с длиной волны равной 320 нм и регистрируется флуориметром на длине волны равной 450 нм. Исходя из данных, что АНС в альбумине располагается рядом с триптофанилом, обуславливающим собственную флуоресценцию белка, следует, что возможна миграция энергии с триптофана на зонд АНС. Если возбуждающий свет на спектрофлуориметре поставить на длину волны 290 – 280 нм (что характерно для спектра поглощения триптофана), а регистрировать флуоресценцию на 450 нм, то можно наблюдать миграцию энергии. Такой приём отражён на рисунках 5 – 7 (кривые на графиках данных рисунков отмечены квадратами (λвозб.
290 нм) и кружками (λвозб.
280 нм).
На рисунках 11 – 13 изображены графики, показывающие как изменяется спектр поглощения триптофана и зонда в длинах волн, при различных рН средах.
Суммируя выше изложенное можно заключить, что падение интенсивности собственной и зондовой флуоресценции в присутствии мочевины обусловлено её денатурирующим действием. Так, известно, что в растворах мочевины неполярные находятся в менее энергетически выгодном окружении, чем в чисто водных растворах. Это ведёт к дестабилизации гидрофобных связей. В результате третичная структура начинает нарушаться, а триптофанил стремится уйти в более гидрофобную часть белковой глобулы. Свидетельством тому является наблюдаемое на графиках (рисунков 11 – 13) коротковолновое смещение спектров флуоресценции триптофана. Обычно коротковолновые сдвиги флуоресценции триптофана при попадании в неполярное сопровождаются и увеличением ИФ, однако в эксперименте взяты настолько высокие концентрации мочевины, что происходит обширная дезорганизация нативной структуры альбумина. При таком разрушении третичной структуры триптофан возможно лишается того микроокружения в котором он был в нативном белке, что и уменьшает интенсивность флуоресценции. Так же возможны и процессы тушения флуоресценции триптофана соседними группами, в результате конформационного изменения альбумина.
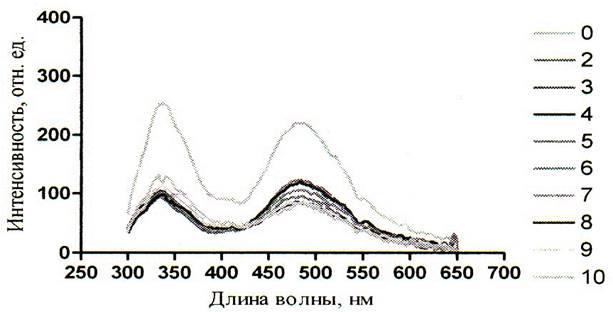
Рисунок 11 – Спектры зондовой флуоресценции альбумина (280 нм) при действии на него мочевиной (2 – 10М) при рН 4,54

Рисунок 12 – Спектры зондовой флуоресценции альбумина (280 нм) при действии на него мочевиной (2 – 10М) при рН 7,43
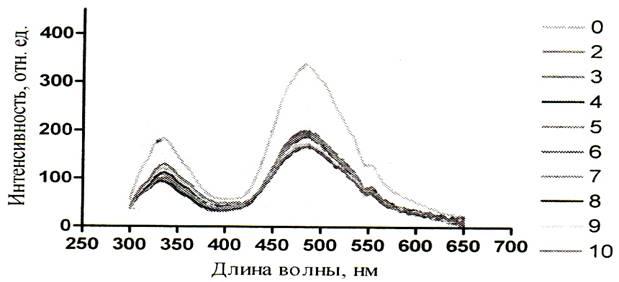
Рисунок 13 – Спектры зондовой флуоресценции альбумина (280 нм) при действии на него мочевиной (2 – 10М) при рН 9,08 [48]
Обращает на себя внимание и тот факт, что даже при концентрации мочевины, превышающей физиологическое соотношение белка и мочевины в организме в 100000 раз, мочевина не разрушает первичную и вторичную структуру альбумина. Об этом свидетельствуют графики рисунков 5 – 7 (собственная флуоресценция) и 8 – 10 (зондовая (кривые с треугольниками)). На этих графиках ИФ хотя и падает, но далеко неравна нулю.
Доказательством неполного разрушения третичной структуры может служить тот факт, что интенсивность зондовой флуоресценции неравна нулю, даже при концентрации мочевины 10М. Это значит что зонд – АНС располагается на своём центре в альбумине. Если бы третичная структура была полностью нарушена, то зондовая флуоресценция отсутствовала, что выразилось бы значением зондовой ИФ, равной нулю. Однако тот факт, что ИФ зонда падает, говорит о частичном разрушении центров для АНС.
Ценность зондовой флуоресценции состоит не только в её большей чувствительности к структурным изменениям молекулы белка, но и в том, что поведение зонда в белке является моделью поведения различных метаболитов, которые взаимодействуют с альбумином. Так, например, по результатам зондовой флуоресценции можно спрогнозировать, что при действии высоких концентраций мочевины некоторые метаболиты, переносимые альбумином останутся в связанном с ним состоянии, хотя транспортная функция альбумина будет значительно нарушена.
Интересным является и тот факт, что при значительной дестабилизации третичной структуры, происходит миграция энергии с триптофана на АНС (рисунки 8 – 13). На этих графиках наблюдается значение ИФ, неравное нулю. ИФ с ростом концентрации мочевины постепенно падает и при концентрации 10М падение прекращается, это говорит о существовании определённого денатурирующего предела у мочевины по отношению к альбумину. Т.о. данный факт ещё раз подтверждает то, что мочевина не полностью разрушает конформацию белка. Рисунки 8 – 10 наглядно отражают процесс миграции энергии с триптофана на АНС, где видно падение триптофановой флуоресценции на фоне возрастания зондовой флуоресценции.
При изучении зависимости ИФ собственной от рН среды, взглянув на графики рисунков 5 – 7, можно сделать вывод о том, что ИФ при концентрации мочевины, равной нулю, в щелочной среде намного ниже, чем значения ИФ для кислой и нейтральной среды. Этот факт обусловлен тушением флуоресценции триптофана. Причиной тушения флуоресценции в щелочной среде может являться миграция энергии с триптофанила на ионизированный тирозинил.
Учитывая литературные данные о том, что молекула альбумина обладает высокой степенью шероховатости поверхности, т.е. её поверхность изрезанна, содержит складки, щели карманов и петель, можно сделать вывод, что влияние мочевины в первую очередь направленно на неплотно упакованные участки в структуре белка.
Значительным структурным изменениям, даже при сверхвысоких концентрациях мочевины, возможно, препятствует система S – S – связей, расположенных регулярно по всей длине полипептидной цепи.
3.2 Влияние свободных радикалов на структуру молекулы альбумина
Профиль элюции сывороточного альбумина на TSK -HW55 геле отражен на рисунке 14.
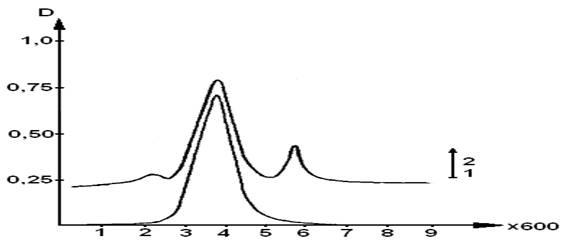
Pиcунок 14 – Профиль элюции сывороточного альбумина на TSK -HW55 геле после действия перекисиводорода с сульфатом меди (II)
1 - исходная мономерная фракция сывороточного альбумина; 2 - мономерная фракция альбумина с СuSO4
через 2 часа после добавленияH2
O2
Флуоресценция белковых фракций после действия пероксида водорода с сульфатом меди (II) представлена на рисунке 15.
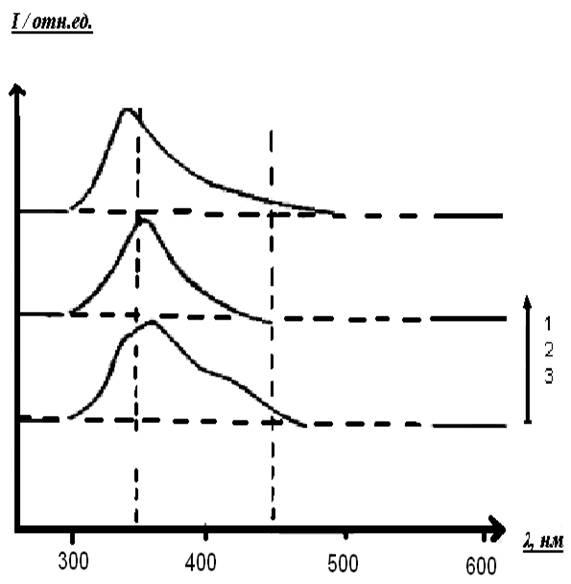
Рисунок 15 – Спектры флуоресценции белковых фракций, после действия CuSO4
через 2 часа после добавления Н2
О2
[49]
1 - димер, 2 - мономер (исходный белок), 3 - белковая фракция с массой меньшей, чем исходный мономер, длина волны возбуждения 290 нм.
При возбуждении белковой фракции УФ излучением волной 290 нм наблюдали появление кроме полосы флуоресценции триптофана дополнительной полосы флуоресценции, которая не наблюдается в исходном белке. Можно предположить, что данное свечение появляется в белке вследствие образования новых флуорофоров при окислении свободными радикалами остатков аминокислот белка. Длинноволновый максимум в спектре флуоресценции совпадает с максимумом полосы свечения, возникающего при действии гидроксильных радикалов на гистидин (рисунок 16).

Рисунок 16 – Спектры флуоресценции триптофана (l) и гистидина(2)
1;1’ и 2;2’ до и после действия СuSO4
иH2
O2
(λ возбуждения 290нм), 2;2’’ до и после действия СuSO4
и H2
O2
(λ возбуждения 360 нм), рН – 6,8
Свободные радикалы в эксперементе образовывались по реакции Вейсса – Габера, суть которой заключается в том, при, что при действии металла переменной валентности, являющегося одноэлектронным донором, на пероксид водорода, образуются свободные радикалы. Реакция Вейсса – Габера состоит из нескольких стадий, показанных на рисунке 17.
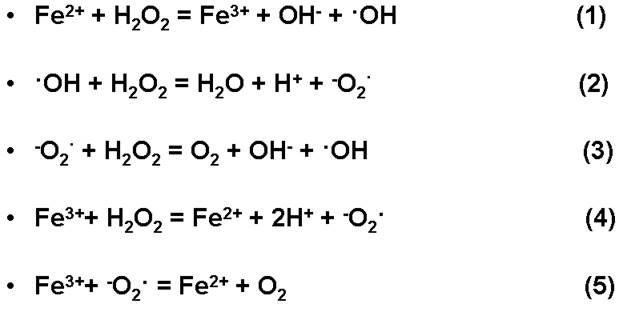
Рисунок 17 – Реакция Вейсса – Габера
Обращает на себя внимание тот факт, что при воздействии СuSO4
через 2 часа после добавленияH2
O2
на белки тиольные соединения практически не образуются. Этот эффект связан с образованием очень реакционных свободных радикалов ОН∙, которые с высокой скоростью взаимодействуют с S-S связями:
∙ОН+RS-SR→RSOH+RS∙
Радикалы ОН∙, реагируя с дисульфидами, дают сульфоновуюкислоту и другие продукты окисления, вплоть до цистеиновой кислоты.
Образовавшиеся радикалы RS∙ взаимодействуют с высокой скоростью с радикалами ОН∙. Так как скорость реакции радикалов ОН∙ с тиолами выше, нежели с цистином, то реакции:
RSН+ОН∙→ RS∙+H2
О и RS∙+ОН∙ → RSОH
приобретают основное значение, и тиольные продукты не образуются.
Образовавшиеся радикальные продукты R, взаимодействуя с ∙ОН, дают также аланин, серин, глицин, но только в очень малых количествах. Образовавшиеся анионы сероводорода окисляются радикалами ОН.
Однако в присутствии кислорода очень эффективно протекает реакция с образованием супероксидного радикала, дисмутация которого дает в конечном результате перекись водорода Н2
О2
. Пероксид водорода, в свою очередь в результате реакции с супероксидным радикалом генерирует чрезвычайно активный окислитель – гидроксильный радикал.
ЗАКЛЮЧЕНИЕ
В результате проведённого эксперимента по влиянию сверхвысоких концентраций мочевины на сывороточный альбумин, были получены данные, исходя из которых, можно сделать вывод, что концентрации мочевины 2 – 10 М не способны полностью разрушить третичную структуру белка, не говоря уже о первичной и вторичной структурах альбумина. Данный факт ещё раз подтверждает уникальность строения молекулы альбумина.
Несмотря на не полное разрушение третичной структуры, данный факт сильно угнетает способность альбумина транспортировать различные низкомолекулярные метаболиты организма.
Полученные результаты можно с успехом перенести на человеческий сывороточный альбумин, что очень важно для медицины в изучении патогенеза таких заболеваний как почечная недостаточность, цирроз печени.
Также, исследуя качество структуры молекулы альбумина, флуоресцентным методом, можно спрогнозировать степень тяжести состояния больного в очень короткие сроки.
Влияние свободных радикалов на молекулу альбумина показало, что происходит разрушение альбумина на уровне первичной структуры. Причём главная роль в этом разрушении принадлежит сильно реакционно способному радикалу ∙ОН.
Гидроксильные радикалы могут образовываться при повышении содержания в организме тяжёлых металлов, которые при взаимодействии с пероксидом водорода, по реакции Вейсса – Габера, образуют различные агрессивные формы кислорода, окисляющие альбумин. В результате таких процессов альбумин не может выполнять ключевых функций для организма.
Данный эксперимент можно также использовать в качестве модели влияния ионизирующего излучения на альбумин, в силу того, что при действии ионизирующего излучения на водные растворы, происходит радиолиз воды, сопровождающийся образованием свободных радикалов.
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1 Граник, В. Г. Основы медицинской химии / В. Г. Граник. – М.:Вузовская книга, 2001. – 384 с.
2 Альбумин сыворотки крови в клинической медицине / Под ред. Ю. А. Грызунова, Г. Е. Добрецова. – М.: ИРИУС, 1994. – 226 с.
3 Чёгер, С.И. Транспортная функция сывороточного альбумина / С. И. Чёгер. – Бухарест.: Изд-во Академии СРР, 1975. – 183 с.
4 Луйк, А.И. Сывороточный альбумин и биотранспорт ядов / А. И. Луйк, В. Д. Лукьянчук. – М.: Медицина, 1984. – С. 12-29.
5 Биохимия человека / Р. Мари, Д Греннер, П Мейес, В Родуэлл. – М.: Мир, 1993. – С.256-273.
6 Альбумин сыворотки крови в клинической медицине. Книга 2 / Под ред. Ю. А. Грызунова, Г. Е. Добрецова. – М.: Медицина, 1998. - С. 28-51.
7 Кантор, Ч. Биофизическая химия: в 3 т. Т.2. Пер. с англ / Ч. Кантор, П. Шиммел,– М.: Мир, 1984. –– 496 с.
8 Практическая химия белка: Пер. с англ. / Под ред. А. Дарбре. – М.: Мир, 1989. – 623 с.
9 Кольман, Я. Наглядная биохимия: Пер. с нем. / Я. Кольман, К.-Г. Рем,– М.: Мир, 2000. – 469 с.
10 Березов, Т.Т. Биологическая химия: Учебник. – 3-е изд., перераб. и доп. / Т.Т. Березов, Б. Ф. Коровкин,– М.: Медицина, 1998. – 704 с.
11 Лабораторная гематология / С. А. Луговская, В. Т. Морозова, М. Е. Почтарь, В. В. Долгов – М.: ЮНИМЕД-пресс, 2002. - 116 с.
12 Гольдберг, Е.Д. Справочник по гематологии / Е.Д. Гольдберг. – Томск: Изд-во ТГУ, 1989. – 420 с.
13 Физиология кровообращения /Отв. ред. Б.И. Ткаченко. – Л.: Наука, 1984. – 652 с.
14 Физиологические показатели организма здорового человека: Морфологический состав и биохимические показатели крови / Е.К. Алимова и др. – Ростов н/Д., 1985. – 84 с.
15 Лебедева, М. И. Аналитическая химия и физико-химические методы анализа: учеб. пособие / М. И. Лебедева. – Тамбов: Изд-во Тамб. гос. техн. ун-та, 2005. – 216 с.
16 Бартенев, Г.М. Физика полимеров / Г. М. Бартенев, С. Я. Френкель; под ред. А.М. Ельяшевича. – Л.: Химия, 1990. – 432 с.
17 Браун, Д. Спектроскопия органических веществ: Пер. с англ. / Д. Браун, А. Флойд, М. Сейнзбери. – М.: Мир, 1992. – 300 с.
18 Сенов, П. Л. Фармацевтическая химия / П. Л. Сенов. – 8-е изд., перераб. и дополн. – М. : Медицина, 1978. – 480 с.
19 Черницкий, Е. А. Спектральный люминесцентный анализ в медицине / Е. А. Черницкий, Е. И. Слабожанин. – Мн.: Наука и техника, 1989. – 141 с.
20 Машковский, М. Д. Лекарственные средства. В двух частях. Ч.1. / М. Д. Машковский. – 12-е изд., перераб. и дополн. – М.: Медицина, 1998. – 736 с.
21 Добрецов Г. Е. Флуоресцентные зонды в исследовании клеточных мембран и липопротеинов / Г. Е. Добрецов. – М.: Наука, 1989. – 277 с.
22 Захаревский, А. С.. Фармакология с рецептурой: учебник / А. С Захаревский, Б. Б. Кузьмицский, Л. Д. Курлович. – Мн.: Высш. шк., 2004. – 304 с.
23 Руководство к лабораторным занятиям по биологической химии / Под ред. Т. Т. Березова. – М.: Медицина, 1976. – 294 с.
24 Миллер, Ю. И. Использование флюоресцентного зонда в оценке связывающей способности сывороточного альбумина человека при печеночной недостаточности / Ю. И. Миллер // Лаб. Дело. – 1989. - №7. – С. 20-23.
25 Тарусов, Б. Н. Биофизика / Б. Н. Тарусов, В. Ф. Антонов. – М. : Высшая школа, 1969. – 467 с.
26 Владимиров, Ю. А. Флуоресцентные зонды в исследовании биологических мембран / Ю. А Владимиров, Г. Е. Добрецов. – М.: Наука, 1980. -320 с.
27 Биохимические методы исследования в клинике / Под. Ред. А. А. Покровского. – М.: Медицина, 1969. – 652 с.
28 Казицына, С. Я. Биофизика / С. Я. Казицына, И. В. Сомова. – М.: Высшая школа, 1990. – С. 212-215.
29 Лакович Дж. Основы флуоресцентной спектроскопии / Дж Лакович. - М. : Мир, 1986. - 496 с.
30 Биофизика: учебник / Ю. А. Владимиров, Д. И. Рощупкин, А. Я. Потапенко, А. И. Деев,– М. : Медицина, 1983. – 272 с.
31 Маршелл Э. Биофизическая химия в 2 томах / Э. Маршелл. - М.: Мир, 1981. – 358 с.
32 Современные методы биофизических исследований: практикум по биофизике / Под ред. А. Б. Рубина. - М.: Высшая школа, 1988. – 359 с.
33 Фрайфелдер Д. Физическая химия / Д. Фрайфелдер. – М.: Мир, 1980. – 582 с.
34 Черницкий, Е. А. Спектральный люминесцентный анализ в медицине / Е. А. Черницкий, Е. И. Слабожанина. – Мн.: Наука и техника, 1989. – С. 142.
35 Пиккеринг У.Ф. Современная аналитическая химия / У. Ф. Пиккеринг. – М.: Химия, 1977. – 559 с.
36 Кузяков, Ю. Я. Методы спектрального анализа / Ю. Я. Кузяков, К. А. Семененко, Н. Б. Зоров. – М.: МГУ, 1990. 231 с.
37 Тагер А. А. Физико-химия полимеров / А. А. Тагер. – М.: Химия, 1978. – 544 с.
38 Практикум по биофизике: Учебн. Пособие для студ. высш. учеб. заведений / В. Ф. Антонов, А. М. Черныш, В. И. Пасечник, С. А. Вознесенский, Е. К. Козлова,– М.: Владос, 2001. – 352 с.
39 Ремизов, А. Н. Медицинская и биологическая физика: Учеб. для вузов. – 4-е изд., перераб. и дополн. / А. Н. Ремизов, А. Г. Максина, А. Я. Потапенко. – М.: Дрофа, 2003. – 560 с.
40 Красовицкий, Б. М. Органические люминофоры. – 2-е изд. перераб. / Б. М. Красовицкий, Б. М. Болотин,– М.: Химия, 1984. – 336 с.
41 Вилков, Л. В. Физические методы исследования в химии. Резонансные и электрооптические методы: Учеб. для хим. спец. вузов. / Л. В. Вилков, Ю. А. Пентин,– М.: Высшая школа, 1989. – 288 с.
42 Пиккеринг, У.Ф. Современная аналитическая химия / У.Ф. Пиккеринг. – М.: Химия, 1977. – 559 с.
43 Кузяков, Ю.Я. Методы спектрального анализа / Ю.Я. Кузяков, К.А. Семененко, Н.Б. Зоров. – М.: МГУ, 1990. – 231 с.
44 Практикум по биофизике: Учебн. пособие для студ. высш. учеб. заведений / В.Ф. Антонов, А.М. Черныш, В.И. Пасечник, С.А. Вознесенский, Е.К. Козлова. – М.: Владос, 2001. – 352 с.
45 Ремизов, А.Н. Медицинская и биологическая физика: Учеб. для вузов. – 4-е изд., перераб. и дополн. / А.Н. Ремизов, А.Г. Максина, А.Я. Потапенко. – М.: Дрофа, 2003. – 560 с.
46 Красовицкий, Б.М. Органические люминофоры. – 2-е изд. перераб. / Б.М. Красовицкий, Б.М. Болотин. – М.: Химия, 1984. – 336 с.
47 Вилков, Л.В. Физические методы исследования в химии. Резонансные и электрооптические методы: Учеб. для хим. спец. вузов. / Л.В. Вилков, Ю.А. Пентин. – М.: Высшая школа, 1989. – 288 с.
48 Корноушенко, Ю.В. Влияние различных концентраций мочевины на конформационные состояния сывороточного альбумина / П.А. Авдеев, Ю.В. Корноушенко // Проблемы и перспективы развития медицины в постчернобыльский период: сборник научных статей Республиканской научно-практической конференции студентов и молодых ученых: в 2 т./ УО «Гомельский государственный университет»; сост. А.Н. Лызиков [ и др.]. – Гомель, 2008. – Т.1. – С. 3-5.
49 Корноушенко, Ю.В. Изучение механизма влияния металла переменной валентности Сu2+
и Н2
О2
, на конформацию сывороточного альбумина / Ю.В. Корноушенко, П.А. Авдеев // Проблемы и перспективы развития медицины: сборник научных статей I республиканской научно-практической конференции с международным участием студентов и молодых ученых: в 3 т./ УО «Гомельский государственный университет»; сост. А.Н. Лызиков [ и др.]. – Гомель, 2008. – Т.1. – С. 164-167.
|