Профессор Е.Е. Аракелян, профессор Ю.Н. Примаков, кандидат педагогических наук, доцент В.В. Тюпа, кандидаты педагогических наук Д.Д. Умаров, Ф.Д. Гусейнов, Российская государственная академия физической культуры
Обширный материал, накопленный по биомеханике бега, позволяет достоверно оценивать связь скорости передвижения с рядом показателей техники бега в неутомленном состоянии. В то же время почти не изучен вопрос влияния утомления на изменение техники бега на финише. В частности, здесь актуально выявление специфического влияния утомления. До настоящего времени выделен пока один такой признак - постановка более выпрямленной ноги на опору при беге на 200 и 400 м [4, 7, 13-15]. Изменение других показателей техники бега под влиянием утомления специфическим не является и соответствует общим закономерностям бега. Например, при беге на финише, когда скорость передвижения снижена, растут вертикальные колебания общего центра масс тела (ОЦМ), уменьшаются длина и частота шагов, понижается беговая посадка, нога ставится на опору дальше, а угол вылета ОЦМ растет [4, 7, 13-16]. Однако точно такие же изменения происходят и при беге в неутомленном состоянии, при переходе на более низкую скорость [4, 12-15]. Понятно, что факт повышенной вертикальной механической работы при беге на выносливость вряд ли правомерно считать признаком менее техничного бега у спортсменов низкой квалификации, бегущих с низкой скоростью [21].
Таблица 1. Скорость бега
| Показатели |
Дистанция, м |
| Усл. обозн. |
100 |
200 |
400 |
800 |
15000 |
| п=20 |
n=20 |
n=25 |
n=23 |
n=27 |
| Начало бега |
 |
8,93 |
8,80 |
7,42 |
6,75 |
5,50 |
| G |
0,81 |
0,73 |
0,85 |
0,49 |
0,40 |
| Финиш |
|
8,42 |
7,42 |
5,87 |
6,31 |
4,95 |
| G |
0,76 |
0.88 |
0,71 |
0,49 |
0,44 |
| Разность, % |
-5,7 |
-15,7 |
-20,9 |
-6,4 |
-9,9 |
Примечание: n - количество испытуемых, и G - среднеарифметическое и средиеквадратическое отклонения (м/с).
Очевидно, что указанные зависимости являются отражением общей закономерности бега независимо от того, в каком состоянии он проводится [4, 15]. Другими словами - кинематический рисунок бега соответствует лишь скорости передвижения, за редкими исключениями. Именно поэтому в нашей работе была поставлена задача: выявить специфичное влияние утомления на технику бега.
Методика. В исследовании приняли участие 115 бегунов квалификации от II разряда до мастера спорта, специализирующихся в беге на короткие, средние и длинные дистанции.
Регистрировались биомеханические показатели бега с установкой на максимально лучший результат на дистанциях 100, 200, 400, 800, 5000 и 15 000 м. Скорость бега определялась фотодиодными датчиками, динамические показатели - с помощью тензоплатформы ПД-3 с собственной частотой не менее 300 Гц, установленной за 5 м до финиша. По опорным реакциям методом двойного интегрирования определялись перемещение и скорость ОЦМ бегуна, по которым по общепринятой методике рассчитывались механическая энергия, работа и мощность [4, б, 18, 19]. Более подробно расчет этих данных приводится в [14].
Показатели техники бега на финише сравнивались с такими же, полученными при беге в неутомленном состоянии, перед забегами на дистанцию. Для спринтеров это был бег с ходу на 30 м с максимальной скоростью, для средневиков и стайеров - бег с индивидуальной "крейсерской" скоростью, характерной для начала дистанции. Таким способом моделировался бег в "начале" дистанции.
Основным показателем, анализируемым в нашей работе, является мощность взаимодействия ноги с опорой - отрицательная в фазе торможения и положительная в фазе отталкивания. При этом изменение механической энергии делится на время соответствующей фазы и на массу бегуна [18]. Рассчитанный таким образом показатель ценен тем, что дает опосредованное представление о мощности, развиваемой мышцами при их принудительном растяжении в фазе торможения и мощности сокращения в фазе разгона ОЦМ.
Таблица 2. Отрицательная мощность при беге в утомленном состоянии
| Показатели |
Дистанция, м |
| 100 |
200 |
400 |
800 |
15000 |
Реальные 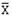 |
34,8 |
30.5 |
26,3 |
24,1 |
16,2 |
| Расчетные |
31,6 |
27,2 |
20,4 |
22,3 |
16,4 |
| Разность, Вт/кг |
3,2 |
3,3 |
5i9 |
1.8 |
-0,2 |
Результаты и их обсуждение.
Поиск специфики утомления проводился следующим образом. Если имелась высокая корреляционная связь показателей техники и скорости бега всех 115 испытуемых в "начале" забе гов, то выводились линейные уравнения регрессии. Затем в уравнение подстав лялись величины скорости бега на фи нише и находилась расчетная величина показателя. Сравнение расчетных и реальных величин показателя техники бега на финише позволяло выявить те, которые не укладывались в общую закономерность бега. В этом случае реальные показатели по величине должны быть больше или меньше тех, которые соответствовали бы бегу в неутомленном состоянии, но с той же скоростью бега, как на финише.
Для начала приведем данные о скорости бега на всех дистанциях, поскольку ее величины необходимы для расчетов (табл. 1).
Затем приведем уравнение:
Рторм. = -5,288 + 4,38V (±5,б2), г = 0,75, где Рторм. - продольная слагаемая отрицательной мощности (Вт/кг), V - скорость бега (м/с), r - коэффициент корреляции. Теперь, подставляя в уравнение значения скорости бега на финише каждой дистанции, получим расчетную отрицательную мощность (табл. 2).
Сопоставим расчетные и реальные значения. Видно, что на дистанции 400 м реальная мощность не соответствует скорости бега на финише, превышая ту мощность, которая должна бы быть показана (различие выходит за пределы стандартной ошибки оценки уравнения, равной 5,62). Это несколько неожиданный факт, поскольку утомленные мышцы смогли развить более значительную мощность в уступающем режиме работы. Видимо, это стало возможным при более жесткой постановке ноги, как было зафиксировано при беге на 200 и 800 м за счет ее выпрямления в дистапьных суставах [4, 7, 13, 15]. Такой способ позволяет снизить диссипативные потери механической энергии в фазе торможения, поскольку переход энергии по ноге к голеностопному суставу производится с большим использованием костей [5]. Кроме того, этот прием помогает несколько снизить потери скорости ОЦМ в фазе торможения. Однако радикально повлиять на мощность сокращения мышц опорной ноги в фазе отталкивания это уже не может, что и видно из анализа положительной мощности. Ее связь с отрицательной мощностью выражается следующим образом:
Рот. = 1,801 + 1,288 Рторм. (±9,06), г=0,76, где Рот. - продольная слагаемая положительной мощности (Вт/кг). Подставляя в уравнение значение отрицательной мощности, реально показанной на финише дистанции 400 м (табл. 1), находим, что положительная мощность соответственно должна быть равной 34,1 Вт/кг. На самом же деле реальная величина меньше на 38% и равна 21,2 ± 7,2 Вт/кг.
Из вышеизложенных фактов следует, что при беге на 400 м в фазе декомпенсированного утомления, когда скорость бега необратимо упала, нарушается зависимость между принудительным растяжением и последующим сокращением мышц. Мало того, что утомленная мышца сокращается с низкой мощностью, при таком нарушении неизбежно ухудшается и механизм рекуперации энергии, что дополнительно снижает мощность отталкивания и скорость бега. Известно, что этот механизм повышает эффективность движений [8, 5, 6, 19, 20]. Его проявление связано с биомеханическими свойствами мышц - чем выше жесткость и сила мышц и меньше время их растяжения, тем больше может утилизоваться накопленной механической энергии [2, 5, 6, 20]. Бег на высокой скорости с укороченной фазой торможения более выгоден для проявления механизма рекуперации с использованием других свойств мышц. Усиление этого механизма удачно демонстрируется с применением резиновых рекуператоров, повышающих упругие свойства мышц - подошвенных сгибателей стопы, основного движителя при беге [10]. И наоборот, при заземлении бега, когда увеличивается время растяжения мышц, накопленная механическая энергия в значительной мере рассеивается в мышцах.
Тогда встает вопрос: может быть, нарушение зависимости "скорость - растяжение" мышц при беге на 400 м связано с чрезмерным ростом фазы торможения? Обратимся к фактам. На финише время торможения выросло на 31%, до 80 ± 15 мс. Однако сравнение расчетных и реальных величин показывает, что они не имеют существенных различий и соответственно равны 0,073 и 0,080 с. Это рассчитано по уравнению:
t0торм. = 0,126 - 0,009V (±0,009), r = -0.83,
где tropм. - время торможения (с). Таким образом, время торможения соответствовало скорости бега на финише и не могло быть причиной нарушения в зависимости "скорость - растяжение" мышц. Видимо, это нарушение связано с ухудшением биомеханических свойств мышц при утомлении [б]. И то, что оно проявилось при беге на 400 м, соответствует остроте утомления именно на этой дистанции, так как здесь зарегистрирован наибольший сдвиг кислотно-щелочного равновесия крови [З]. А это приводит к подавлению функциональной активности нервных клеток [3, 11, 17], резкому падению содержания АТФ и Крф и увеличению АДФ [1].
Итак, выявленные зависимости позволяют количественно оценить соотношение мощности растяжения и сокращения мышц опорной ноги при беге в состоянии острого утомления. Но, что нам представляется не менее существенным, эти данные подтверждают идею о необходимости повышения. локальной мышечной выносливости. То, что она имеет высокую связь с результатом в беге на средние дистанции, установлено экспериментально [8]. Повышение этого качества неизбежно приведет к улучшению биомеханических свойств мышц, что позволит на более качественном уровне использовать их упругие свойства и успешнее противодействовать падению скорости бега на финише в состоянии острого утомления.
Выводы
1. Специфика утомления в беге проявляется на дистанции 400 м, выражаясь в нарушении соотношения мощности растяжения и сокращения мышц.
2. Подтверждается идея о повышении локальной мышечной выносливости, что позволит увеличить эффективность бега в состоянии острого утомления.
Список литературы
1. Батунвр Л.С. "Физиологический журнал СССР". 1979, т. 65, № 1, с. 128.
2. Верхошанский Ю.В. //Теор. и практ. физ. культ. 1970, № 12, с. 8.
3. Волков Н.И. Физиологические основы современных методов развития выносливости. М., 1962.
4. Гусейнов Ф.А. Канд. дисс. М., 1982.
5. Зациорский В.М., Аруин А.С., Селуянов В.Н. Биомеханика двигательного аппарата человека. - М.: ФиС, 1981.
6. Зациорский В.М., Алешинский С.Ю., Якунин Н.А. Биомеханические основы выносливости. - М.: ФиС, 1982.
7. Комаров А.И. Автореф. канд. дисс. М., 1974.
8. Нурмекиви А., Лемберг X. //Легкая атлетика. 1990, № 7, с. 27.
9. Озолин Н.Г., Чхаидзе Л.В. //Теор. и практ. физ. культ. 1951, № 5, с. 339.
10. Попов Г., Чапайкин В. //Легкая атлетика. 1991, № 6, с. 15.
11. Розенблат В.В. //Теор. и практ. физ. культ. 1958, т. 21, вып. 3,с.195.
12. Степанов В.В. Автореф. канд. дисс. М., 1982.
13. Тюпа В., Чистяков В., Алешинский С. и др. //Легкая атлетика. 1981, № 9, с. 10.
14. Тюпа В.В., Травин Ю.Г., Гусейнов Ф.А. и др. //Теор. и практ. физ. культ. 1982, № 4, с. 11.
15. Тюпа В.В., Гусейнов Ф.А., Мироненко И.Н. //Теор. и практ. физ. культ. 1989, № 2, с. 33.
16. Чхаидзе Л.В. //Теор. и практ. физ. культ. 1948, № 10, с. 457.
17. Яковлев Н.Н., Александрова Г.В, Батунер Л.С. и др. //Физиологический журнал СССР. 1978, т. 64, № 11, с. 1510.
18. Fukunaga Т., Matsuo A., Yuasa К. а.о. //Ergonomics, 1980, vol. 23, N 2, р. 123.
19. Cavagna G.A. //J. Physiol., Paris, Sept. 1969, N 61, р. 4.
20. Cavagna G.A., Thys H., Zamboni A. //J. Physiol. 1976, 262, р. 639.
21. Miura M., Kobayashi К., Miyashita М. а.о. In: Review of our researches. 1970-1973 (ed. H. Matsui). Univ. of Nagoya, 1973, p.
|