Введение.
Математическое моделирование биосинтеза продуктов метаболизма
Реферат на сдачу экзамена по кандидатскому минимуму по специальности 03.00.23 – Биотехнология.
Аспирант: Осипов Д.С.
Московский Государственный Университет Инженерной Экологии, Кафедра «Экологическая и промышленная биотехнология»
Москва – 2002
Введение.
МЕТАБОЛИЗМ – греческое слово metabole, означающее перемена, превращение.
В физиологическом смысле метаболизм – это промежуточный обмен, т. е. превращение определенных веществ внутри клеток с момента их поступления до образования конечных продуктов (напр., метаболизм белков, метаболизм глюкозы, метаболизм лекарственных препаратов).
С точки зрения промышленной биотехнологии метаболизм – это образование в процессе роста и развития клеток ценных биохимических продуктов – некоторые из них выделяются в среду (внеклеточные продукты), некоторые накапливаются в биомассе (внутриклеточные продукты). С помощью метаболизма получают антибиотики, молочную и лимонную кислоту, пищевые консерванты и многие другие продукты.
В естественных условиях метаболизм настроен так, чтобы производить минимальное количество необходимых метаболитов. Промышленное производство, направленное на получение максимальной прибыли, такая ситуация никак не устраивает. Поэтому, для максимизации прибыли необходимо произвести оптимизацию следующих технологических параметров:
– выхода продукта в расчете на потребленный субстрат;
концентрация продукта;
скорость образования продукта.
Оптимизация технологии биосинтеза метаболитов состоит из следующих основных этапов [5]:
1) Первоначальная селекция штамма микроорганизмов;
2) Определение оптимальных значений температуры, рН, тоничности и потребности в кислороде;
3) Определение оптимального режима питания и накопления биомассы;
4) Изменение генетической структуры организма для увеличения образования продукта.
Разработка 3-его этапа, непосредственно связана с биосинтезом. Для нахождения режима питания и накопления биомассы, оптимального для биосинтеза метаболитов необходимо математическое описание процесса [1,2,3,4]. Кроме нахождения оптимальных условий проведения процесса, математическая модель используется для автоматизации биосинтеза, что в современной биотехнологической промышленности не менее важно.
Прежде чем приступить, к описанию моделей приведем принятые обозначения основных количественных характеристик процесса биосинтеза.
Кинетические характеристики процесса биосинтеза.
Обычно состояние процесса определяется следующими основными параметрами:
– концентрация биомассы микроорганизмов – Х, г/л;
– концентрация питательной среды – субстрата (или его основного компонента) – S, г/л.
концентрация продукта – P, г/л.
Кинетические характеристики процесса отражают скорость протекания биохимических превращений. Эти превращения, естественно, отражаются на всех указанных выше параметрах процесса – биомассе, продукте и субстрате.
Важным показателем процесса является скорость роста биомассы. Для описания скорости роста используется такая характеристика, как общая скорость роста – QX:
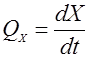 (1). (1).
Больший интерес для характеристики интенсивности роста представляет не величина QX, а удельная скорость роста в пересчете на единицу биомассы (ведь рост биомассы пропорционален концентрации клеток). Она обозначается буквой m:
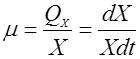 (2). (2).
Размерность величины m – [1/ч].
Рассмотрим теперь второй параметр процесса ферментации – концентрацию субстрата S. По аналогии с ростом биомассы, можно ввести кинетическую характеристику – скорость потребления субстрата QS:
 (3). (3).
Знак ‘–‘ обозначает, что скорость потребления положительна, когда концентрация субстрата в среде падает (т.е. скорость изменения концентрации отрицательна).
Аналогично, удельная скорость потребления субстрата, которую обозначим малой буквой qS, равна:
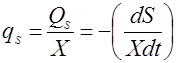 (4). (4).
При биосинтезе метаболитов, наряду с ростом биомассы, происходит накопление в среде продукта метаболизма (его текущая концентрация – Р).
Общая скорость биосинтеза продукта метаболизма QР в периодическом процессе равна:
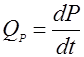 (5). (5).
Удельная скорость биосинтеза продукта единицей биомассы обозначается qР и равна:
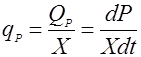 (6). (6).
Для математического описания биосинтеза продуктов метаболизма различными исследователями было предложено много различных моделей. Все эти модели можно разделить на следующие группы:
1) Математические модели кинетики биосинтеза продуктов метаболизма как функция от удельной скорости роста;
2) Субстрат-зависимые модели кинетики биосинтеза продуктов метаболизма;
3) Модели, основанные на концепции возраста культуры микроорганизмов .
Во время биосинтеза происходит также процесс уменьшения количества метаболитов, для описания этого феномена были предложены модели деградации (инактивации) продуктов метаболизма.
Математические модели кинетики биосинтеза продуктов метаболизма как функции от удельной скорости роста.
Математический параметр – удельная скорость роста m – послужил основой составления многих математических моделей биосинтеза продуктов метаболизма. Процессы биосинтеза продуктов издавна делят на два больших класса – связанные с ростом и не связанные с ростом. В качестве примера первого класса можно назвать биосинтез конститутивных ферментов клетки, а второго класса - биосинтез многих антибиотиков, интенсивный синтез которых происходит после прекращения роста микроорганизмов.
Удельная скорость биосинтеза связанных с ростом продуктов может быть выражена простым соотношением [5]:
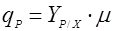 (7), (7),
где YP/X – выход единицы продукта с единицы биомассы: (dP/dX).
Более сложное выражение было предложено Людекингом и Пайри [6]:
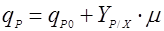 (8), (8),
где qP0 – эмпирическая константа.
В этом случае биосинтез продукта, с одной стороны, ассоциирован с ростом, а, с другой, осуществляется покоящейся клеткой [7]. Модель (8) впервые была предложена для описания синтеза молочной кислоты.
Есть ряд уравнений, учитывающих нелинейный характер связи qP и удельной скорости роста:
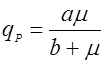 (9), (9),
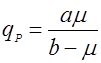 (10), (10),
где а и в – эмпирические константы.
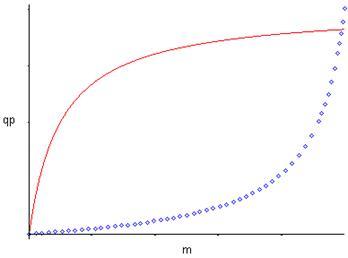
Рис. 1. Форма зависимости qР(m) для уравнений (9) и (10).На рис. 1 показаны графики функций (9) и (10), эти уравнения дают выпуклую (9) и вогнутую (10) кривые, выходящие из нуля, но эти функции могут иметь также дополнительный свободный член qP0:
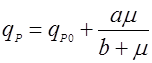 (11), (11),
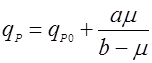 (12). (12).
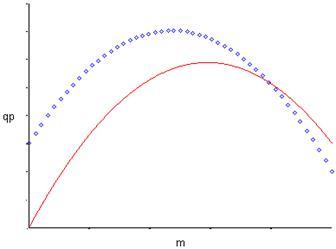
Тогда графики функций (11) и (12) в отличии от (9) и (10) выходят не из нуля, а из некоторой точки qP0 на оси y, что продемонстрировано на рис. 2.Рис. 2. Форма зависимости qР(m) для уравнений (11) и (12).
Возможны также эмпирические уравнения типа [8,9]:
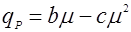 (13), (13),  (14). (14).где а, b, с – эмпирические константы.
Рис. 2. Форма зависимости qР(m) для уравнений (13) и (14).
По аналогии с уравнениями (11) и (12) уравнение (14) начинается, не из 0 по оси ординат, а из некоторой точки a, что означает начало синтеза продукта без роста биомассы.
Субстрат-зависимые модели кинетики биосинтеза продуктов метаболизма.
С точки зрения математики, уравнения, в которых в качестве аргумента выступает m, предполагают, что совсем неважно, каким образом формируется то или иное значение m. Например, величину m можно изменять путем уменьшения концентрации углеродного субстрата или азотного субстрата, или путем снижения температуры или повышения величины рН. Для процессов, связанных только с ростом, возможно скорость биосинтеза при этом будет одинакова. Для несвязанных с ростом процессов небезразлично, каким путем мы будем изменять величину m. Лимитирование углеродом, лимитирование азотом, повышение рН или снижение температуры, давая одно и то же значение скорости роста, могут давать совершенно различные скорости биосинтеза продукта метаболизма. Другими словами, связь между qP и m не имеет строго причинно-следственного характера, а обусловлена влиянием на обе эти кинетические характеристики одних и тех же факторов внешней среды. Для таких процессов необходимо использовать уравнения, которые в качестве аргументов содержат независимо влияющие первичные факторы: концентрация того или иного субстрата, температура или величина рН.
Биосинтез продукта может описываться однофакторными или многофакторными уравнениями. Кроме того, было установлено, что структуры зависимостей qP от S, P, температуры и величины рН аналогичны структурам таких же уравнений для роста биомассы, например: Моно, Андрюса, Перта, Хиншельвуда и т.д. Например, если субстрат влияет на qP по Андрюсу, то имеем [10]:
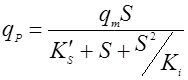 (15), (15),
где qm – максимальная удельная скорость биосинтеза продукта;
K’S – константа насыщения;
Ki – константа ингибирования продуктом.
Многофакторные зависимости здесь чаще бывают мультипликативными, чем аддитивными. Приведем зависимость мультипликативного [11] и аддитивного влияния концентрации субстрата по механизму Моно:
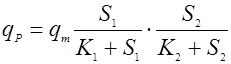 (16), (16),
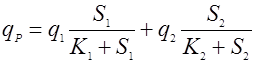 (17). (17).
Применяются также уравнения с не разделяющимися эффектами факторов, например, типа Контуа [10] или неконкурентного торможения продуктом [5]:
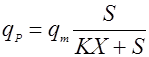 (18), (18),
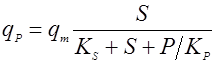 (19). (19).
К сожалению, невозможно изложить все кинетические зависимости биосинтеза продуктов от первичных факторов в столь короткой работе. Данный материал подробно изложен в монографии [14], в которой приведены не только несколько десятков уравнений, но и произведен их анализ.
Модели, основанные на концепции возраста культуры микроорганизмов.
Для биосинтеза продуктов метаболизма часто бывает недостаточно только благоприятных “внешних” факторов среды. Потому что в биосинтезе участвуют внутриклеточные ферменты микроорганизмов, промежуточные продукты, содержание которых в клетке зависит от предыстории развития культуры. Слишком быстро выросшая культура часто неэффективна с точки зрения биосинтеза продукта. У микробиологов есть выражение “культура ушла в ботву”, что означает биомассы – много, продукта – мало или вообще нет. Однако, учитывать эти внутриклеточные компоненты при моделировании очень проблемно – их трудно измерять и соответственно находить кинетические коэффициенты.
Вместо этого предложены некоторые феноменологические подходы к оценке физиологического состояния микробной биомассы, основанные на оценке возрастного состояния популяции клеток.
Есть несколько подходов для учета возраста культуры. Один из них заключается в определении распределения клеток микроорганизмов по возрастам [12]. Тогда значение удельной скорости биосинтеза продукта можно считать как бы суммой скоростей, даваемых разными возрастными фракциями биомассы:
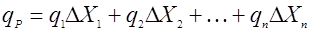 (20), (20),
где ΔXi – концентрация биомассы i-ой возрастной группы;
qi – удельная скорость биосинтеза биомассой i-ой возрастной группы.
При этом, вполне возможно, что значения q1, q2, ... ,qn не будут одинаковыми: “молодежь” не синтезирует нужный продукт, слишком старые клетки – тоже.
Японским ученым Аибой был предложен более простой подход, использовать для оценки возраста культуры так называемый средний возраст популяции 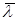 как параметр, определяющий биосинтетическую активность культуры [13]. Биологически термин вполне понятен – это сумма возрастов всех клеток, деленная на их количество: как параметр, определяющий биосинтетическую активность культуры [13]. Биологически термин вполне понятен – это сумма возрастов всех клеток, деленная на их количество:
 (21), (21),
где λi – возраст i-ой возрастной группы.
Если последовательно уменьшать поддиапазоны Δt и ΔΧ, доведя их до бесконечно малых dX и dt, то для среднего возраста можно получить интегральную формулу:
 (22), (22),
где Х0 – начальная концентрация биомассы;
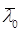 – средний возраст культуры в начальный момент культивирования. – средний возраст культуры в начальный момент культивирования.
Другим способом упрощения возрастной зависимости является разделение возрастного диапазона клеток на 2 класса – продуктивный (выше некоторого значения) и не продуктивный [14]:
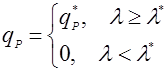 (24), (24),
где λ* – возраст зрелости;
qP* – удельная скорость биосинтеза клетки, по достижении ею возраста зрелости.
Теперь остается рассмотреть форму зависимости удельной скорости биосинтеза продукта qР от среднего возраста культуры: 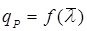 . .
Если зависимость имеет возрастающий характер с насыщением, то зависимость удобно выразить в форме, похожей на уравнение Моно:
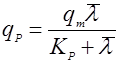 (25). (25).
Если, наоборот, она падает с возрастом, то лучше подходит выражение, подобное уравнению Иерусалимского:
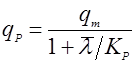 (26). (26).
Если зависимость имеет экстремум, то оно может быть выражена, например, с помощью аппроксимирующего полиномиального уравнения [15]:
 (27). (27).
Однозначная зависимость между qР и на практике встречается редко, часто зависимость скорости биосинтеза продукта от возраста учитывают в виде мультипликативного сомножителя, сопряженного с основной частью уравнения, учитывающего влияния остальных факторов.
Модели деградации (инактивации) продуктов метаболизма.
Не всегда синтезированные продукты метаболизма остаются устойчивыми; часто они настолько нестабильны, что разрушаются уже в процессе самой ферментации. Поэтому, описывая материальный баланс по продукту метаболизма, необходимо учитывать кинетику его инактивации:
 (28), (28),
где  – скорость деградации продукта метаболизма. – скорость деградации продукта метаболизма.
При рассмотрении синтеза метаболитов, использовалась удельная скорость, в случае деградации, вводить удельную скорость не корректно, т.к. продукт существует отдельно от биомассы, и его деградация не зависит в общем случае от ее концентрации.
Рассмотрим модели кинетики деградации:
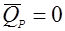 (29), (29),
деградация отсутствует.
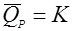 (30), (30),
деградация идет с постоянной скоростью. Такое выражение странно выглядит в начале процесса, когда продукта еще нет; из уравнения же получается, что концентрация продукта снижается ниже нуля, что не имеет физического смысла.
 (31), (31),
реакция разложения первого порядка, пропорционально количеству образовавшегося продукта [16].
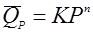 (32), (32),
реакция разложения n-ого порядка, при чем n может быть как больше 1, так и меньше и не быть целым числом.
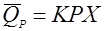 (33), (33),
реакция разложения зависит не только от концентрации продукта, но и от концентрации биомассы.
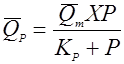 (34), (34),
скорость реакции разложения зависит от концентрации биомассы и возрастает с концентрацией продукта до какого-то предела.
Приведенные уравнения инактивации (29)-(34) наиболее распространенные, существуют также и другие более сложные зависимости.
Модель накопления продукта метаболизма на примере лейцина.
L-лейцин- незаменимая аминокислота, необходимая для промышленного получения лизина. Производство лизина базируется на лейцинозависимых штаммах. Годовое производство лизина составляет приблизительно 500000 т/г. Лизин широко используется в с/х в качестве кормовой добавки. Лейцин также применяется в спортивном питании, т.к. является предшественником незаменимых жирных кислот, входящих в состав клеточных мембран.
Элементная формула L-лейцина (L-a-аминоизокапроновая кислота): C5H10NH2COOH.
Основным способом производства L-лейцина является микробиологический синтез с использованием штамма Corynebacterium glutamicum. Биосинтез проводился в лабораторном биореакторе. В отбираемых пробах определялась оптическая плотность – Х, содержание лейцина – P и содержание редуцирующих веществ (РВ) по Бертрану – S.
Полученные результаты приводятся в таблице 1.
Таблица 1.
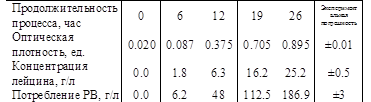
Простейшим предположением о механизме микробиологического биосинтетического процесса является обобщение данных о том, что биосинтез, с одной стороны, ассоциирован с ростом, а с другой, осуществляется покоящейся клеткой.
Поэтому было решено использовать следующее соотношение (8):
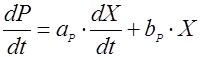 (35), (35),
где P – концентрация продукта (лейцина), г/л;
aP, bP – эмпирические константы.
aP, bP были определены методом наименьших квадратов: aP=13.27, bP=1.249, сумма квадратов отклонений QP составила 0,884, а средняя квадратичная ошибка SP равна ± 0.6648.
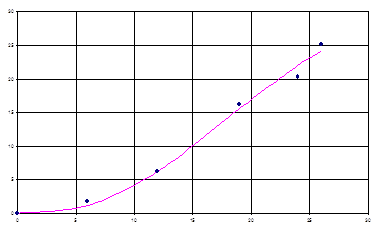
На рис. 3 показаны экспериментальные значения концентрации лейцина и модельные, из рисунка видно, что модель (8) в данном случае достаточно хорошо описывает биосинтез продуктов метаболизма.
Рис. 2. Графическая интерпретация модели биосинтеза лейцина (35).
Список литературы
1. Арзамасцев А.А., Андреев А.А Математические модели кинетики микробного синтеза: возможности использования и новые подходы к разработке // Вестн. Тамб. ун-та. Серия: Естеств. и техн. науки.– 2000.– т.V., № 1– с. 111-130.
2. Renss M. Моделирование и оптимизация процессов // 8th Int. Biotechnol. Symp., Paris– 1988.– vol. 1.– p. 523-536.
3. Zeng An-Ping Кинетическая модель получения продуктов микробных клеток и клеток млекопитающих // Biotechnol. and Bioeng.– 1994.– vol. 45., N 4.– p. 314-324.
4. Vanrolleghem P.A. Структурный подход для выбора среди кандидатов в модель схемы метаболизма и установление неизвестных стехиометрических коэффициентов // Biotechnol. and Bioeng.– 1998.– vol. 2., N 3.– p. 133-138.
5. Перт С.Дж. Основы культивирования микроорганизмов и клеток.– М.: Мир.– 1978.
6. Luedeking R., Piret E.L. A kinetic study of the lactic acid fermentation: Batch process at controlled pH // J. Biochem. Microbiol. Technol. Eng.– 1959.– vol. 1., N 4.– p. 393-412.
7. Осипов Д.С., Гусельникова Т.В. и др. Математическая модель биосинтеза L-лейцина // Труды МГУИЭ.– 2001.– т.V.– с. 19-23.
8. Mori A., Terui G. Kinetic studies on submerged acetic acid fermentation: Inhibition by ethanol // J. Ferment. Technol.– 1972.– vol. 50, N 11.– p. 776-786.
9. Музыченко Л.А., Валуев В.И. Использование полунепрерывного культивирования микроорганизмов для получения продуктов биосинтеза // В кн.: Теория и практика непрерывного культивирования микроорганизмов.– Красноярск.– 1978.– с. 112-113.
10. Bajpai R.K., Reuss M. A mechanistic model for penicillin production // J. Chem. Technol. and Biotechnol.– 1980.– vol. 30,– p. 332-344.
11. Баснакьян И.А., Бирюков В.В., Крылов Ю.М. Математическое описание основных кинетических закономерностей процесса культивирования микроорганизмов // В кн.: Итоги науки и техники. Микробиология. Т. 5. Управляемое и непрерывное культивирование микроорганизмов.– М. – 1976.– с. 5-75.
12. Shu P. Mathematical model for product accumulation in microbiological processes // J. Biochem. Microbiol. Technol. Eng.– 1961.– vol. 3, N 1,– p. 95-109.
13. Aiba S., Hara M. Conception of average cumulative age of microorganisms // J. Gen. and Appl. Microbiol.– 1965.– vol. 11,– p. 25-41.
14. Бирюков В.В., Кантере В.М. Оптимизация периодических процессов микробиологического синтеза.– М.: Наука.– 1985.
15. Фишман В.М. Математическое описание и оптимальное управление процессом биосинтеза антибиотиков // Дис. канд. техн. наук.– М.: Московский институт химического машиностроения– 1970.
16. Ettler P., Votruba J. Determination of the optimal feeding regime during biosynthesis of erythromycin // Folia microbiol.– 1980.– vol. 25,– p. 424-429.
|