«огневой воздух» (кислород) и выделяли «связанный воздух» (углекислоту). Ингенгуз и Сенебье примирили наблюдения обоих исследователей, признав за растением обе функции газообмена. Ингенгуз писал в своей работе, опубликованной в 1779 г.:
«Когда солнце, поднявшееся над горизонтом, разбудит своими лучами заснувшие за ночь растения, оно сделает их способными исполнять свою целительную функцию – исправлять воздух для животных; во мраке ночи эта деятельность совсем прекращается; днем же совершается с тем большим оживлением, чем светлее день и чем выгоднее расположено растение в отношении солнечных лучей. Затененные высокими зданиями или другими растениями, они не исправляют воздух, а, наоборот, выделяют вредный для дыхания животных воздух. К концу дня выработка очищенного воздуха ослабевает и при заходе солнца совершенно прекращается».
Первые точные исследования процесса дыхания у растений принадлежат Соссюру (1804). Он брал свежие листья и помещал их на ночь в сосуд, наполненный воздухом. При этом кислород воздуха поглощался и выделялся углекислый газ. Если на следующий день листья снова выставлялись на солнечный свет, то они выделяли почти такое же количество кислорода, какое поглотили ночью. Свои исследования Соссюр распространил и на незеленые части растений: стебли древесных растений, цветки, корни, плоды, и доказал, что дыхание наблюдается также в клетках этих органов. Он обнаружил, что при дыхании потеря в весе растения равна весу выделенного углерода.
Соссюр обратил внимание и на то, что молодые, растущие части растения, например новые побеги и распускающиеся цветки, дышат интенсивнее и потребляют кислорода больше, чем части растения, прекратившие рост. Особенно интересные данные получены Соссюром относительно дыхания цветков и связанного с ним повышения температуры.
До раскрывания цветков початок поглощал сравнительно малый объем кислорода, превосходивший объем початка не более чем в 8 раз, и оставался совершенно холодным. Как только цветки начинали распускаться, объем поглощаемого кислорода сразу возрастал, превосходя объем початка в 30 и более раз, и температура соцветия повышалась. Исследуя нагревание отдельных частей соцветия, Соссюр установил, что тычинки нагревались сильнее других частей цветка, что он объяснил их более интенсивным дыханием.

Зависимость интенсивности дыхания семян проса от влажности

Влияние температуры на интенсивность дыхания семян пшеницы
Дальнейшими исследованиями была установлена зависимость дыхания от внешних условий (освещения, температуры и т.п.). Выяснилось, что интенсивность дыхания возрастает почти прямо пропорционально температуре, но только до известных пределов (около 40 °С). При дальнейшем повышении температуры газообмен остается постоянным до гибели растения от перегрева.
Удалось установить косвенную зависимость дыхания растений от освещения, а также от стадии роста. Развитие дыхательных процессов у растущих частей выражается так называемой большой кривой дыхания растений. Она сходна с ранее открытой физиологами большой кривой роста: растение растет сначала медленно, затем постепенно скорость роста увеличивается, доходит до максимума и так же постепенно падает.
Со времен Лавуазье дыхание организмов отождествлялось с медленным горением, причем его сущностью считалось прямое окисление углеводов и жиров организма кислородом вдыхаемого воздуха. Однако уже давно было замечено, что при дыхании окисление органических веществ проходит гораздо полнее, чем вне организма при той же температуре. Это давало виталистам надежду на установление коренного различия между процессами, совершающимися в организме и вне его. В таком неопределенном положении находился вопрос о химизме дыхания до середины XIX в.
Переломный момент в изучении дыхания растений связан с открытием того, что даже в бескислородной среде растения (а равно и животные) продолжают выделять углекислоту. Теоретическое истолкование это явление впервые получило в работах Луи Пастера (1822–1895), который в 1872 г. обратил внимание ботаников на его сходство со спиртовым брожением, обычным у дрожжевых грибков.
Пастер предположил, что некоторые начальные этапы дыхательного процесса у высших растений и животных сходны с процессом анаэробного дыхания у микроорганизмов. Свои предположения Пастер обосновывал данными, полученными упомянутыми выше физиологами: в отсутствие кислорода высшие растения выделяли углекислоту, а в их тканях накапливался спирт.
Идеей Пастера заинтересовался Пфеффер (1878). Одностадийное окисление углевода кислородом воздуха (С6Н12О6 + 6О2 = 6СО2 + 6Н2О), использовавшееся до того для описания дыхания, он разбил на два последовательных этапа:
1) С6Н12О6 = 2С2Н5ОН + 2СО2,
2) 2С2Н5ОН + 6О2 = 4СО2 + 6Н2О.
Первая стадия соответствовала бескислородному дыханию и представляла собой распад сахара на спирт и углекислоту. Вторая стадия, требующая кислорода, состояла в окислении спирта до углекислоты и воды.
Примерно через год Вортман (1879) предложил другую схему. По его мнению все количество углекислоты выделяется сразу в первой фазе брожения:
3С6Н12О6 = 6С2Н5ОН + 6СО2, а на второй стадии образовавшиеся молекулы спирта присоединяют молекулы кислорода воздуха, что приводит к восстановлению части исходного сахара и выделению воды:
6С2Н5ОН + 6О2 = 2С6Н12О6 + 6Н2О.
Обе теории были простыми, но благодаря им широкие круги химиков и физиологов обратили внимание на процесс брожения. С этого времени процессы брожения становятся объектом пристального изучения.
Во второй половине XIX в. область изучения процессов брожения представляла собой арену борьбы различных школ. Школа Бертло, открывшего фермент инвертазу, отстаивала точку зрения, согласно которой процессы брожения обусловлены особыми веществами – ферментами. Школа Пастера выдвигала теорию «ферментов-существ», т.е. считала микробы активным началом процессов брожения.
В 1883 г. японский химик Иошида установил ферментативную природу окисления органических веществ растительного сока. Как известно, японцы в свое время достигли большого совершенства в кустарном производстве черных лаковых изделий. Иошида решил выяснить, как образуется черное вещество японского лака из бесцветного сока лакового дерева. Оказалось, что этот процесс, состоящий в окислении сока лакового дерева кислородом воздуха, происходит только в присутствии особого фермента.
Молодой японский химик констатировал только ферментный характер этого процесса. Французский химик Бертран, более детально исследовал этот процесс. Он назвал фермент, ответственный за окисление сока, лакказой (от слова лак) и, исследуя другие растительные соки, пришел к убеждению, что лакказа является представителем целой группы окислительных ферментов, весьма распространенных в природе. Для них Бертран предложил название оксидазы.
В 1897 г. на самом пороге XX в., произошло событие, разрешившее длительный научный спор о «ферментах-существах» и «ферментах-веществах». В этом году химик Бухнер блистательно доказал, что спиртовое брожение – этот оплот виталистов – зависит не от какойто особенной «жизненной силы» дрожжевых грибков, а от вещества, которое Бухнеру удалось выделить из дрожжей при растирании их с песком. Это вещество оказалось таким же растворимым ферментом-веществом, как и ранее известные ферменты. Бухнер назвал найденный им фермент, вызывающий спиртовое брожение, зимазой.
Естественно, что столь крупные открытия в области изучения ферментов, как открытие зимазы (ответственной за спиртовое брожение в сахаристых растворах) и оксидаз (необходимых для окисления ряда растительных соков), поставило перед ботаниками-исследователями весьма серьезный вопрос: не лежат ли подобные ферменты и в основе процесса дыхания растений, не является ли обыкновенное кислородное дыхание растений результатом действия ферментов, подобных оксидазам?
За разрешение этого вопроса взялись работавший в Женеве русский химик А.Н. Бах, швейцарский ботаник Шода и два русских ботаника, работавших в Петербурге, профессора В.И. Палладин и С.П. Костычев.
Бах как химик разрабатывал, главным образом, вопросы тонких механизмов окислительных процессов, происходящих при дыхании. Путь к познанию химизма дыхания он видел в изучении так называемого медленного горения, или произвольного окисления.
Бах считал, что эти процессы протекают при обыкновенной температуре и не нуждаются в резкой активации кислорода путем расщепления его молекулы на свободные атомы. Это утверждение противоречило мнению крупнейших научных авторитетов того времени, утверждавших, что при всякой реакции окисления в организме происходит полный распад молекулы кислорода на два атома.
Бах указывал, что разложение молекулы кислорода на атомы требует слишком больших затрат энергии, которые не наблюдаются в физиологических реакциях окисления. По мнению Баха, в физиологических процессах из молекулы кислорода образуется активная группа, в которой разорвана лишь одна из двух связей в молекуле, но атомы не образуются.
Способное к медленному окислению вещество присоединяет эту группу к себе. При этом неизбежно должны образоваться перекиси. Именно эта реакция первичного образования перекисей в процессе медленного окисления и составляет основное ядро теории Баха. Перекиси, как весьма неустойчивые и химически активные вещества, могут подвергаться дальнейшим изменениям. Правильность своей теории Бах подтвердил на сотнях примеров и фактов, как известных до него, так и полученных им самим экспериментально.
История современного учения о дыхании растений неразрывно связана с именем академика В.И. Палладина.
В годы первого петербургского периода работы Палладин исследовал ферментативную природу дыхательного процесса. Палладин показал, что и анаэробная, и аэробная фазы дыхания обеспечиваются специфическими ферментами, последовательно перерабатывающими продукты дыхания. Итоги работ этого периода изложены в монографии В.И. Палладина «Дыхание как сумма ферментативных процессов» (1907).
На этом этапе исследовательской деятельности Палладин применил оригинальную методику исследований. Замораживая цельные органы растений при температуре минус 15–20 °С, он показал, что даже в замороженных, т.е. убитых, клетках работа ферментов, а следовательно и дыхательные процессы, продолжаются, хотя и с отклонением от нормы. Тем самым Палладину удалось сорвать завесу таинственности с процесса дыхания, который трактовался виталистами как специфический жизненный акт, как одно из важных проявлений пресловутой «жизненной силы».
Продолжая изучение ферментов специфических для реакций окисления, Палладин задумался над обстоятельством, тогда еще не обратившим на себя внимание западно-европейских физиологов: окислительная способность известных к тому времени ферментов (оксидаз и пироксидаз) была совершенно недостаточной для реакций прямого окисления их основного субстрата – углеводов. Следовательно, помимо оксидаз в клетках растений должны содержаться какие-то другие вещества, являющиеся посредниками в передаче кислорода основному дыхательному материалу.
Палладин обратил внимание на так называемые хромогены клеточной плазмы, которые он считал дыхательными пигментами. Дыхательные пигменты, по мысли Палладина, подобно гемоглобину крови, являются переносчиками кислорода, воспринимая его при содействии ферментов (оксидаз) и перенося далее к дыхательному материалу.
Основы своей теории дыхательных пигментов Палладин изложил в получившей широкую известность на Западе статье с сенсационным названием «Кровь растений». Однако последующие исследования, проведенные ученым в последние годы жизни, заставили его пересмотреть всю теорию дыхательного процесса.
Для объяснения новых фактов и наблюдений Палладину пришлось свою теорию «поставить на голову». Хромогены оказались факторами не окислительных, а, наоборот, восстановительных процессов. Публично признать неправильность своих недавних утверждений мог только человек, ставивший научную истину, выше самолюбия. Палладин нашел в себе мужество разрушить здание научной теории, построение которой отняло у него свыше 10 лет жизни.
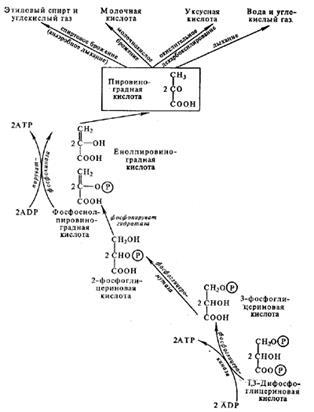 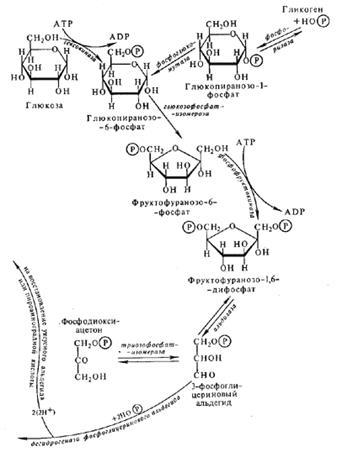
Схема процессов дыхания и брожения у растений
Вот что рассказывает об этом великом переломном моменте в развитии учения о дыхании растений ближайший ученик Палладина профессор С.Львов.
«Исходя из тех соображений, что дрожжи продолжают в присутствии кислорода вести распад сахаров по бродильному руслу лишь потому, что они лишены окислительного аппарата и, в частности, дыхательных хромогенов, Палладин предложил мне использовать соки, отжатые из растений, богатых хромогенами (шампиньоны, свекла и т. п.), и проследить, не вызовет ли добавка их к бродильной среде поворота процесса от брожения к дыханию.
Ожидаемого эффекта не получилось, но при производстве опытов бросилось в глаза неожиданное явление, что в период активного брожения черная и бурая окраски от добавочных соков быстро светлели, особенно в анаэробных условиях, т.е. хромогены вели себя совершенно аналогично метиленовой синьке. Иначе говоря, хромогены являются отнюдь не переносчиками кислорода, подобно гемоглобину, как полагал раньше Палладин, а переносчиками водорода. Они активизируют не кислород воздуха при помощи оксидаз, а водород дыхательного субстрата при помощи редуктаз. Дыхательный материал окисляется в первой стадии не в силу того, что он воспринимает доходящий до него активизированный кислород, а в силу того, что от него отщепляется активизируемый при помощи редуктаз и дыхательных хромогенов водород. Дыхательные хромогены должны быть переименованы в водородные акцепторы.
Все эти столь привычные для нас теперь представления в тот момент впервые получили свое зарождение. Я хорошо помню, с каким волнением присматривался Палладин к каждому новому моему опыту, ожидая его результатов. Длительные с ним беседы, во время которых зарождались новые понятия, естественно расширяли и мои горизонты и заполняли душу тем научным энтузиазмом, которым всегда горел Палладин.
Доклад об этой работе в научном обществе он предложил сделать мне самому, указывая, что такое выступление весьма полезно для начинающего научного работника, поскольку подготовка к докладу побудит меня еще глубже продумать всю проблему и активизирует дальнейшую работу…
Я помню период, когда Палладин, в отступление от своей обычной нормы, стал несколько реже появляться в лаборатории. При встречах в беседах он, особенно сосредоточенный и нервновозбужденный в это время, несколько раз упоминал при мне, что он дома, в спокойной обстановке, занят обдумыванием новой теории дыхания. Вскоре она была окончательно оформлена и предложена на суд и внимание товарищей по науке. Это и была та знаменитая теория дыхания, кратко формулированная в виде двух уравнений, которая произвела полный переворот в обычном понимании дыхательного процесса и теперь вошла в основные руководства по физиологии растений…
…Все ее содержание в концентрированной форме запечатлено в следующих двух уравнениях:
I. С6Н12О6 + 12R + 6Н2О = 6СО2 + 12RН2,
II. 12RН2 + 12R + 12Н2O.
Основным уравнением, выражающим собой принципиальную сущность дыхательного процесса, является первое. В этой первой фазе дыхательный материал (глюкоза) уже полностью распадается до своих простейших элементов – СО2 и водорода, но водород не выделяется в свободном состоянии, а захватывается при помощи редуктаз (дегидраз) водородными акцепторами, без которых распад глюкозы вообще не может иметь места. (Процесс идет чисто анаэробно.)
Кислород к этому основному акту дыхания не имеет никакого отношения. Весь СО2 – анаэробного происхождения, и уносимый с нею кислород заимствуется не из внешней атмосферы, а изнутри системы. Дыхание для своего осуществления не нуждается ни в кислороде, ни в оксидазах. Основными ферментами в этой фазе дыхания являются дегидразы.
Кислород воздуха вмешивается в акт дыхания лишь во второй фазе, но, принципиально рассуждая, играет здесь второстепенную роль. Он должен восстановить активность водородных акцепторов, освободив их от захваченного водорода. Кислород окисляет не углерод дыхательного субстрата, а уже отщепленный от него водород, перенесенный на акцепторы. Сам кислород выступает лишь в роли последнего заключительного водородного акцептора…
…Эта принципиально новая точка зрения на процесс дыхания, выраженная Палладиным в столь ясно отчетливой форме, не была еще при нем достаточно широко обоснована экспериментальными данными и явилась в значительной мере плодом научной интуиции автора, хотя в ней вместе с тем мы должны видеть как бы подведение итогов многолетней работы Палладина над процессом дыхания…
Весь этот эпизод с нашей работой я рассказал подробно для того, чтобы исторически, как непосредственный участник этой работы, отметить, что принципиальная перестройка основных воззрений Палладина на роль дыхательных хромогенов произошла у него совершенно независимо от работ Виланда. Даже другие чисто химические работы Виланда, в которых показана возможность окисления в отсутствии кислорода, при наличии водородных акцепторов, таких веществ, как СО и SO, стали известны уже после выполнения нашей работы, а переход Виланда к изучению биологических проблем произошел и по времени позднее».
Одновременно с Палладиным проблемой дыхания занимались в целом ряде крупнейших научно-исследовательских институтов и лабораторий Западной Европы. Наибольшую популярность приобрели две новые школы – Виланда и Варбурга.
Т.Виланд развивал взгляды на роль дегидраз и водородных акцепторов, вполне аналогичные взглядам Палладина. Расхождение их теорий заключалось в том, что Виланд категорически отрицал какую бы то ни было роль оксидаз как специфических активаторов кислорода, считая молекулярный кислород способным самостоятельно отнимать водород от водородного акцептора. По мнению же Палладина, водородные акцепторы не могут самопроизвольно освобождаться от водорода, но требуют для этого участия оксидаз, которые поэтому являются обязательным фактором в реакции, выраженной во втором уравнении Палладина.
Противник Виланда, Варбург, считал, что молекулярный кислород не может вступить в организме в какой бы то ни было окислительный процесс, если в организме отсутствует система железоорганических соединений, типичным представителем которых он считал геминфермент. Варбург утверждал, что геминфермент активирует молекулярный кислород, т.е. как бы дает первый толчок к началу окислительных процессов, и без него никакой дыхательный процесс не может совершаться. Далее, по мнению Варбурга, окислительный импульс через промежуточные звенья (геминовые соединения) доходит до дыхательного субстрата и окисляет его. Резюмируя свои взгляды, Варбург утверждал, что дыхание осуществляется путем активации кислорода, а отнюдь не водорода. Но ведь Палладин как раз и говорил о той же необходимости активации молекулярного кислорода, защищая перед Виландом роль оксидаз в процессе дыхания.
Все различие в основных посылках Варбурга и Палладина заключается в том, что первый, работая по преимуществу с объектами животного происхождения, называл свой активатор молекулярного кислорода геминферментом, а Палладин, работавший с объектами растительного происхождения, сохранил за этим активатором ранее установившееся в науке название оксидазы. Но по существу оба говорили об одном и том же, протестуя против непримиримой позиции Виланда, отрицавшего необходимость энзиматической активации молекулярного кислорода.
Согласно взглядам большинства современных научных авторитетов Запада обе теории – и Виланда, и Варбурга – оказываются правильными, они охватывают различные фазы одного и того же чрезвычайно сложного процесса. Современная наука находит разрешение вопроса в синтезе обеих теорий, называя современное учение о внутреннем механизме дыхательных процессов теорией Виланда–Варбурга.

Цикл Кребса:
1, 6 – система окисления и декарбоксилирования; 2 – цитрат-синтеза: кофермент А; 3, 4 – аконитат-гидратаза; 5 – изоцитратдегидрогеназа; 7 – сукцинатдегидрогеназа; 8 – фумарат-гидрогеназа; 9 – малатдегидрогеназа; 10 – спонтанное превращение; 11 – пируваткарбоксилаза
|