Владислав Вильгельмович Хлебович, д.б.н., проф., гл.н.с. Зоологического института РАН
Впервые концепцию о внутренней среде организмов и ее постоянстве сформулировал еще в 1878 г. великий французский физиолог К.Бернар:
"Эта внутренняя среда никогда не изменяется, атмосферные колебания не могут проникнуть за ее пределы, и потому можно с полным правом утверждать, что физиологические условия среды у высших животных неизменны. <…> Организм как бы укрыт в своего рода оранжерее. Непрерывные изменения внешних условий не достигают его, он не подвержен этим условиям, он остается свободным и независимым. <…> Все физиологические механизмы, сколь бы различны они ни были, имеют только одну цель - сохранение постоянства условий жизни во внутренней фазе" [1].
Именно это, описанное Бернаром, постоянство внутренней среды организмов животных, позволяющее им активно существовать при широких колебаниях среды внешней, американский физиолог У.Кеннон предложил в 1929 г. называть гомеостазом (окончательно термин был утвержден в 1932 г., когда была опубликована книга Кеннона "Мудрость тела" [2]).
В наши дни это понятие распространяют и на другие этажи биологической целостности - клетки, популяции, экосистемы и т.д.; мы же ограничимся рассмотрением гомеостаза на уровне организма. И Бернар, и Кеннон писали о постоянстве внутренней среды только высших организмов, прежде всего позвоночных и особенно человека. Это и понятно, - в природе существуют животные, у которых некоторые виды гомеостаза вообще отсутствуют или слабо выражены. Известный эволюционист Б.М.Медников считал становление и совершенствование гомеостаза важнейшим путем прогрессивной эволюции. Действительно, способность организмов к гомеостазу - величайшее эволюционное приобретение, позволившее животным освоить новые, ранее недоступные им среды и пространства, что стало возможным благодаря выработке соответствующих весьма сложных механизмов. К примеру, только терморегуляция позволила животным освоить приполярные области, а в умеренном климате оставаться активными даже суровой зимой и холодными ночами. Благодаря "осморегуляции" (значение кавычек поясню чуть позже) многие животные, занявшие различные ветви эволюционного древа, смогли проникнуть из морей в пресные воды с чрезвычайно низкой минерализацией. При этом гомеостаз мог выработаться по отношению к одному фактору среды и не проявиться - к другому. Так, организм может быть гомойосмотическим, т.е. поддерживать гомеостаз по отношению к осмотическому фактору (как большинство пресноводных организмов), и быть при этом пойкилотермным, т.е. имеющим ту же температуру, что окружающая их среда (этим отличаются холоднокровные животные - рыбы, раки, амфибии).
Есть основания полагать, что в зависимости от важности адаптации к тем или иным условиям могли возникать независимо в разное время и в разных группах. Но тогда хотелось бы знать, на каких именно уровнях в разных независимо произошедших группах поддерживается постоянство внутренней среды по тому или иному фактору. Иными словами, какой параметр фактора избирает природа, чтобы далее поддерживать его постоянство.
Рассмотрим ситуации с факторами, регуляция которых организмами привлекает внимание исследователей чаще других, - температурой, осмотическим давлением и концентрацией водородных ионов (pH).
Терморегуляция
Терморегуляция возникла в эволюции довольно поздно - она характерна только для птиц и млекопитающих. Благодаря приобретенной теплокровности звери и птицы в сравнении с их холоднокровными предками стали более независимы от температуры внешней среды. Сейчас, когда я пишу эти строки, на улице -27оС, а за окном вижу летающих голубей и ворон, гуляющих с хозяевами собак, атакующих кормушку с семечками синиц и усевшуюся на подоконник серебристую чайку - самую крупную дикую птицу Санкт-Петербурга. Ни один из процветавших в тепличных условиях мелового периода динозавров, бывших хозяев Земли, сегодняшнего мороза не выдержал бы. Да и при нуле он бы тоже не выжил.
Два класса высших позвоночных, птицы и млекопитающие, сформировались независимо друг от друга и, очевидно, столь же независимо выработали способность регулировать температуру тела, при этом "избрали" они в качестве оптимальной практически одинаковую температуру - около 37-40оС [3, 4]. Случайно ли?
Любопытно, что примерно до тех же значений температуры разогреваются в полете (или перед ним) мышцы некоторых насекомых, и эти же значения оказываются верхним температурным пределом для жизнедеятельности рыб, а также для сперматозоидов других самых разных холоднокровных животных. Не распространяется это лишь на обитателей гидротермальных вод, живущих при специфических условиях температуры, газового режима и давления на дне моря. Для прочих организмов, населяющих "типичные" биотопы Земли, это заключение, судя по всему, справедливо.
По мнению В.Р.Дольника, "избранная" птицами и млекопитающими температура "очень опасна, так как близка к летальной, приводящей к денатурации белков (43оС у млекопитающих и 45оС у птиц)" [3]. Эту близость температуры тела теплокровных животных к критическим значениям их организменной термоустойчивости пытались объяснить разными причинами. Наиболее правдоподобным выглядело объяснение "завышенной" температуры тела птиц и млекопитающих энергетическими преимуществами физиологии нагрева их тел по сравнению с охлаждением [3]. Действительно, нагрев тела, который происходит за счет усиления обменных процессов или произвольной и непроизвольной активности мышц (дрожи, тонического сокращения), оказывается энергетически выгоднее охлаждения, вызванного испарением воды при дыхании или с поверхности кожи. Казалось бы, именно поэтому теплокровные и "избрали" в качестве постоянной температуру самую высокую из возможных - чуть ниже тепловой смерти. Однако анализ других видов гомеостаза (о которых пойдет речь в следующих главах) не позволяет объяснить избранный уровень температуры тела теплокровных только энергетическими преимуществами теплопродукции перед тратами на охлаждение.
Осморегуляция
Существующие около полутора веков термин и понятие "осморегуляция" не совсем корректны. Дело в том, что осмотическое давление (тоничность) определяется количеством растворенных в объеме жидкости частиц независимо от их природы. А для функционирования организма далеко не безразлично, какие именно частицы растворены во внешней (вода) или внутренней (плазма крови, целомическая жидкость, лимфа, гемолимфа) средах. Как правило, обитающие в море организмы, как и пресноводные животные, нуждаются в некотором количестве определенного набора солей, весьма близкого по соотношению ионов к морской воде. Одно из ярких доказательств этого - сходство по составу и концентрации ионов морской воды и различных физиологических растворов, используемых в качестве заменителей плазмы и других компонентов крови (растворов Рингера). Это сходство дало основание ввести термин "соленость внутренней среды", выражая ее в принятых в океанологии единицах - в граммах солей в литре воды (промилле, ‰) [5].
Доля осмотического давления, создаваемого во внутренней среде органическими веществами - сахарами, аминокислотами, белками (так называемая онкотическая компонента тоничности), обычно очень мала. У некоторых вторично-морских позвоночных (пластиножаберных рыб и немногих видов амфибий) тоничность плазмы крови, соленость которой около 10-15 ‰, повышается до изоосмотичного (близкого по солености) морской среде состояния за счет накопления соответствующего количества мочевины. Так, соленость плазмы крови у обитающей в Юго-Восточной Азии лягушки-крабоеда (Rana cancrivora) равна 28‰, т.е. изоосмотична морской воде, где она проводит большую часть жизни. Эта единственная из всех лягушка способна жить в соленой воде, сохраняя в крови мочевину, и, что интересно, она умеет освобождаться от ее избытка, оказавшись в пресном водоеме, куда отправляется, как все ее сородичи, метать икру.
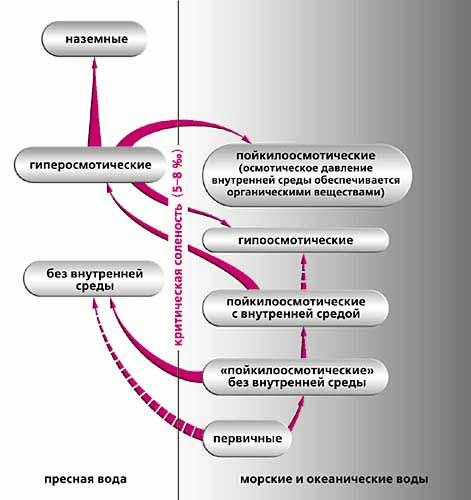
Эволюция осмотических (соленостных) отношений.
Первичные животные появились при солености выше 5-8 ‰, но ниже современной океанической. Произошедшие от них организмы без внутренней среды (нет полостных жидкостей, крови, лимфы и гемолимфы, как у современных турбеллярий) вышли в пресные воды, а также заняли морские со всем спектром солености. Они дали начало пойкилоосмотическим организмам с выраженной внутренней средой (например иглокожим, морским моллюскам, большинству ракообразных). Некоторые из этих животных, только приобретя гиперосмотическую регуляцию, смогли освоить пресные воды соленостью ниже критической (пресноводные раки, моллюски, рыбы и др.). Гиперосмотические пресноводные, сохранив характерные для них значения внутренней солености, дали начало наземным и вторично морским. Большинство вторично морских при этом стали гипоосмотическими (морские рыбы, креветки-палемониды), остальные - морскими пойкилоосмотическими, которые способны повышать тоничность внутренней среды за счет задержки в крови мочевины. Пунктиром показаны менее вероятные пути эволюции.
Румынский академик Э.Пора в свое время предложил рассматривать соленость сред как сочетание двух факторов - осмотического, определяемого общим количеством растворенных частиц, и рапического, обусловленного совместным специфическим действием растворенных ионов. Мы убедились в справедливости этого предположении, изучая физиологическую активность беломорских организмов - личинок-планул медуз Aurelia aurita и взрослых брюхоногих моллюсков Hydrobia ulvae. Животных помещали в сосуды с возрастающей соленостью 2, 4, 6, 8, 10, 12, 14, .., 25‰, куда добавляли многоатомный спирт маннит (он, в отличие, например, от сахаров, метаболически неактивен), доводя во всех случаях среду до общего осмотического давления, соответствующего типичной для Белого моря солености 25‰. Результаты эксперимента показали, что для нормальных реакций всем исследованным организмам обязательно требуется содержание морских солей в среде не менее 5 ‰, и только после этого они могут реагировать на добавленный маннит как на осмотический фактор [6].
Таким образом, когда речь идет об осморегуляции, чаще всего имеется в виду поддержание величин внутренней солености. Или, иными словами, осморегуляция осуществляется, прежде всего, механизмами ионорегуляции.
Отсутствие осморегуляции (пойкилоосмотическое состояние) характерно для большинства морских беспозвоночных, причем это состояние сохраняется в довольно большом диапазоне солености внешней среды - от 34.5‰ (океанические воды) до 5-8‰, очень редко до 3‰. Ниже этого предела вплоть до пресных вод существование организмов возможно лишь за счет работы механизмов гиперосмотической регуляции, "следящей" за тем, чтобы внутренняя соленость клеток и тканей не опускалась ниже 5-8‰ [5].
Выработанный для жизни в пресных водах уровень внутренней солености чуть выше 5-8‰ сохранился у вышедших на сушу потомков пресноводных организмов разных групп. У человека, например, соленость плазмы крови около 9-10‰ - морскую воду, разведенную до этих значений солености, медики Советского Союза и Великобритании использовали во время Второй мировой войны в качестве дешевого кровезаменителя.
Уровень внутренней солености, выработанный в пресных водах, оказался в чем-то настолько выгодным, что его сохранили потомки пресноводных животных, вновь вернувшиеся в море. У таких организмов (костных рыб, креветок-палемонид, ветвистоусых рачков Evadne и Podon) в море осуществляется гипоосмотическая регуляция - у них работают механизмы, снижающие внутреннюю соленость. Это достигается постоянным питьем морской воды и выведением наружу значительной части содержащихся в ней солей специальными клетками покровов или жабр. При солености внешней среды, близкой океанической, соленость плазмы крови таких организмов часто оказывается примерно в три раза меньшей. Именно поэтому французский врач и путешественник Ален Бомбар, переплывший Атлантический океан на надувной лодке, использовал для питья сок, выдавленный из пойманных рыб.
Таким образом, уровень гомойосмии как у гиперосмотических пресноводных, так у гипоосмотических морских примерно одинаков и определяется внутренней соленостью обычно не ниже 5-8‰, но близкой этому пределу. При этом замечу, что осмотический гомеостаз вырабатывался у животных разных таксонов (кольчатых червей, брюхоногих и двустворчатых моллюсков, ракообразных, рыб и др.), переместившихся из моря в пресные воды, независимо друг от друга.
Очевидно, нижний предел внутренней солености тоже располагается очень близко к границе устойчивости белковых комплексов. Об этом можно судить по резкому увеличению оптической плотности тканевых вытяжек разных пресноводных и морских организмов в морской воде соленостью ниже 5-8‰.
Регуляция pH
Здесь лучше всего процитировать монографию Г.А.Виноградова: "Анализ собственных и литературных данных позволяет считать, что у абсолютного большинства различных в таксономическом отношении групп солоноватоводных и пресноводных животных величина pH внутренней среды находится в пределах 7.4-8.0 и фактически не зависит от концентрации водородных ионов во внешней среде" [7, с.193]. Действительно, кровь морских и наземных организмов имеет близкие значения pH (у человека в норме 7.35-7.45). Судя же по широкому распространению в животном мире, гомеостаз концентрации водородных ионов очень древний. Поддерживается он бикарбонатным буфером: при высоких значениях рН угольная кислота выделяется через органы дыхания, бикарбонат - почками.
Нельзя не заметить, что слабощелочная реакция внутренней среды животных также близка к критическому значению - при pH ниже 7.0 среда становится качественно иной, кислой.
В юбилейном сборнике докладов, посвященном 200-летию Московского общества испытателей природы, опубликовано единственное научное сообщение лауреата конкурса студенческих работ - Д.А.Суплатова, четверокурсника МГУ им.М.В.Ломоносова. На примере бактериальных пенициллинацилаз он отметил, что этот фермент стабилен при нейтральной величине pH, а при подщелачивании или подкислении начинается диссоциация его молекулы. Очевидно, такие качественные изменения состояния характерны и для других белков.
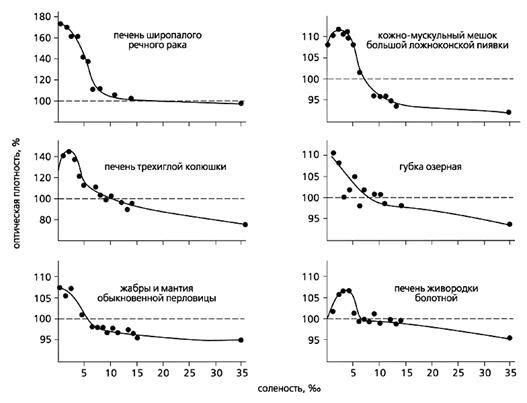
Оптическая плотность тканевых вытяжек пресноводных животных в средах разной солености.
Пунктиром отмечен контроль (100-) - морская вода, соответствующая нормальной внутренней солености организмов.
Общие принципы гомеостаза
Избранные организмами разных таксономических групп уровни регулирования температуры и внутренней солености оказались близки границам устойчивости белковых комплексов, а уровень регулирования pH жидкостей внутренней среды, находясь в слабощелочной зоне, оказывается совсем рядом с нейтральной точкой, за которой среда становится кислой.
Очевидно, есть какие-то преимущества регулирования биологическими процессами термо-, ионорегуляции и регуляции концентрации водородных ионов вблизи границ устойчивости белковых комплексов [8]. Именно у этой опасной черты, но заведомо не переходя через нее, оказываются оптимальными биологические процессы во внутренней среде организмов. Не частные ли это проявления общего принципа преимуществ регулирования вблизи критических точек?
По утверждению В.И.Полежаева и Е.Б.Соболевой, резкие изменения свойств веществ вблизи критических точек могут быть вызваны минимальными воздействиями [9]. Вспомним также управляемые ядерные реакции распада или синтеза. Создаваемые в реакторе условия выделения полезной энергии можно рассматривать как своего рода предвзрыв. Они очень близки к тому, что может привести к взрыву, но делается все, чтобы этот взрыв не произошел. Только вблизи критической точки, не переходя ее, процессом можно управлять, например, в случае реакций распада - перемещением стержней. В теории управления функциональных систем сформулировано правило: "чем ближе реальное возмущающее воздействие приближается к детерминированным процессам, тем меньше мощность управления, требуемая для обеспечения устойчивости" [10].
Возможно, преимущества регулирования процессов вблизи их критических точек стоит рассматривать как проявление принципа минимального действия. Этот принцип Макс Планк считал самым крупным научным обобщением физики, вобравшим в себя как частность даже законы сохранения [11]. Принцип этот часто связывают с именем Мопертюи, но открывался он под разными названиями многими авторами [8, 12]. Известный закон минимума Либиха - частное проявление этого принципа. Он также известен как принцип наименьших сопротивлений Спенсера - Богданова [13]. Мне кажется, принцип минимального действия прекрасно выражен русской пословицей: "Где тонко, там и рвется". Если хотите разорвать - рвите здесь, если укрепить - здесь и штопайте, но в обоих случаях цель будет достигнута минимальными усилиями.
Список
литературы
1. Bernard C. Les phenomenes de la vie. Paris, 1878. (Цитируется в переводе А.Г.Гинецинского.)
2. Cannon W.B. The wisdom of the body. L., 1932.
3. Дольник В.Р. // Журнал общей биологии. 2003. Т.64. №6. С.451-462.
4. Зотин А.И., Зотин А.А. Направление, скорость и механизмы прогрессивной эволюции. М., 1999.
5. Хлебович В.В. Критическая соленость биологических процессов. М., 1974.
6. Хлебович В.В. // Доклады АН СССР. 1972. Т.208. №5. С.1221-1223.
7. Виноградов Г.А. Процессы ионной регуляции у пресноводных рыб и беспозвоночных. М., 2000.
8. Хлебович В.В. // Журнал общей биологии. 2005. Т.66. №5. С.431-435.
9. Полежаев В.И., Соболева Е.Б. Гидродинамика околокритических жидкостей // Природа. 2003. №10. С.17-26.
10. Петров Ю.П., Шишкин В.И. Управление и диагностика функциональных систем // Методические указания к курсу "Функциональные системы" Ч.2. СПб.,1993.
11. Планк М. Единство физической картины мира. М., 1966.
12. Вариационные принципы механики / Ред. Л.С.Полак. М., 1959.
13. Тахтаджян А.Л. Principia tektologika. Принципы организации и трансформации сложных систем: эволюционный подход. СПб., 1998.
14. Суплатов Д.А. pH-зависимость активности и стабильности бактериальных пенициллинацилаз // Доклады на юбилейном заседании, посвященном 200-летию Московского общества испытателей природы. М., 2005. С.48-62.
|