Кандидат биологических наук, доцент Н.Г. Беляев, Ставропольский государственный университет, Ставрополь
Устойчивая адаптация к физическим нагрузкам формируется в результате многолетних тренировок и сопровождается рядом морфофункциональных изменений, расширяющих возможности организма занимающегося. При этом не только возрастает энергетическая емкость систем и их функциональных резервов, но и происходит перестройка регуляторных механизмов. В результате формируются иные взаимоотношения нервных центров, гормональных, вегетативных и исполнительных органов, необходимые для решения задач приспособления организма к конкретным видам спортивной деятельности.
В случае, когда объем тренировочных и соревновательных нагрузок превосходит возможности организма, развивается дизадаптация, характеризующаяся нарушением нейроэндокринной регуляции, перенапряжением адаптивных механизмов и компенсаторных реакций. Возможность предупреждения "срыва" адаптации во многом определяется пониманием интимных механизмов ее развития, что позволяет, используя различные фармакологические средства и изменяя методы тренировок, вносить соответствующие коррективы.
С биологической точки зрения вполне оправданно применение адаптогенов растительного и животного происхождения [1, 6, 8,16]. Перспективны в этом плане также препараты, получаемые на основе корня солодки голой [3 - 5, 7].
Критерием функционального состояния организма может служить не только уровень специальной работоспособности, но и биохимический статус организма. Представляемые результаты демонстрируют, что одним из таких компонентов может оказаться общий кальций (Са). Подобное предположение основывается на важной роли, которую Са выполняет в процессах мышечного сокращения, нервной импульсации, секреции гормонов, в том числе гормонов симпато-адреналовой и гипоталамо-гипофизарно -надпочечниковой системы [10]. Са и циклические нуклеотиды считаются главными компонентами внутриклеточной сигнальной системы во время действия гормонов, изменяющих конкретные метаболические процессы. Учитывая столь многогранную роль Са, в процессе эволюции выработались жесткие механизмы регуляции его гомеостаза [15]. Следовательно, нарушение обмена Cа нужно рассматривать как фактор риска в становлении многих патологических процессов, в том числе и состояния перетренированности.
Согласно литературным данным и результатам собственных исследований уровень Са в крови при мышечных нагрузках определяется характером, интенсивностью и продолжительностью последней. Так, начальный этап выполнения работы нетренированными животными характеризуется увеличением концентрации общего Са в крови. Затем через 30-40 мин работы уровень общего Са начинал понижаться и достигал гипокальциемических величин на 60-180-й мин бега [2, 11, 12]. Принципиально такие же результаты были получены при определении общего Са в плазме людей, выполнявших велоэргометрическую нагрузку в течение 3 ч [13]. При физических нагрузках обнаружено также увеличение ионизированного кальция (Са++) в крови нетренированных людей [18, 20]. Мы в своих предыдущих исследованиях отмечали однонаправленные изменения как общего, так и ионизированного Са в период выполнения нагрузки, а также в восстановительном периоде [2]. Однако большая часть имеющихся результатов отражает динамику кальциевого обмена при разовых нагрузках, не прослеживается его изменение в процессе становления адаптивных реакций и тем более их поломки.
Таблица 1. Схема тренировки крыс микроциклами в течение 9 недель (в минутах)
| Дни недели |
Недели |
| 1-я |
2-я |
3-я |
4-я |
5-я |
6-я |
7-я |
8-я |
9-я |
| Понедельник |
1 |
7 |
3 |
3 |
3 |
3 |
27 |
27 |
3 |
| Вторник |
2 |
8 |
13 |
13 |
13 |
13 |
13 |
13 |
4 |
| Среда |
3 |
9 |
12 |
27 |
3 |
27 |
27 |
27 |
5 |
| Четверг |
4 |
10 |
3 |
27 |
13 |
27 |
3 |
3 |
5 |
| Пятница |
5 |
11 |
27 |
3 |
3 |
3 |
27 |
27 |
5 |
| Суббота |
6 |
12 |
13 |
12 |
12 |
13 |
3 |
27 |
6 |
Примечание: 1-4-я недели - скорость бега 20 м/мин, 5-я неделя - 23 м/мин, 6-8-я недели - 25 м/мин, 9-я неделя - 20 м/мин.
Таблица 2. Динамика изменения работоспособности животных при моделировании состояния перетренированности
| Условия опыта |
Дни наблюдений |
| 1-й |
5-й |
10-й |
15-й |
18-й |
19-й |
20-й |
25-й |
30-й |
| Продолжительность бега животных, мин контрольная группа |
40 |
90 |
140 |
190 |
203±7,8 |
175,4±6,3 |
170,1±6,4 |
89,6±5,8 |
32,8±3,6 |
| экспериментальная группа |
40 |
90 |
140 |
190 |
220 |
230 |
201,8±7,1 |
112±8,5 |
33,9±2,9 |
| Скорость движения ленты, м/мин |
20 |
25 |
30 |
35 |
38 |
39 |
40 |
30 |
25 |
С учетом этого обстоятельства целью данной работы было изучение концентрации общего Са, глюкозы, эритроцитов и белков плазмы ( общего белка, альбуминов) в процессе формирования адаптации и дизадаптации - перетренированности, а также изучение возможности использования экстракта солодки как средства, повышающего физическую работоспособность организма и предупреждающего возникновение состояния перетренированности.
Методы исследования. Экспериментальным материалом служили крысы линии Вистар, систематически выполнявшие мышечные нагрузки по методике, предложенной Ю.П. Похоленчуком [14]. В данной методике выделяются следующие этапы тренировочного цикла: I - подготовительный (1-4-я недели); II - разгрузочный (5-6-я недели); III - цикл интенсивной нагрузки (6-8-я недели); IV - цикл активного отдыха (9-я неделя). Схема тренировки представлена в табл. 1.
С целью повышения эффективности от используемых нагрузок мы внесли небольшие изменения в данную методику. В частности, во II и III циклах тренировок интенсивность нагрузки для животных была увеличена. Эксперимент проведен на предварительно подготовленных животных. С этой целью за 4-5 дней до основного эксперимента в течение 2-3 дней у животных вырабатывали условный рефлекс - бег по движущейся ленте. Для этого животных заставляли в течение 15-20 мин выполнять бег по движущейся ленте с невысокой скоростью. Животные, прекратившие передвижения по ленте, попадали на электростимулирующию решетку, роль которой заключается в выработке и подкреплении у крыс рефлекса "избегания". Тем самым формировался навык постоянного нахождения на движущейся ленте и выполнения заданной работы. Животные, у которых отмечалась низкая работоспособность или была затруднена выработка рефлекса бега по движущейся ленте, в эксперимент не включались. В том случае, если
у животных моделировалось состояние перетренированности, последний этап тренировочного микроцикла исключался. Начальная продолжительность бега для животных составляла 40 мин и затем постоянно увеличивалась при одновременном увеличении скорости движения ленты. В наших исследованиях работу подобного характера животные способны были выполнять в течение 17-20 дней (табл. 2), затем их работоспособность снижалась. Забор крови для биохимического анализа осуществлялся до начала тренировок, на различных этапах тренировочного цикла и через каждые 5 дней моделируемого состояния перетренированности. Определение концентрации общего Са осуществлялось методом тестирования [17], при подсчете эритроцитов использовалась камера Горяева. Общий белок, альбумины, глюкозу определяли с помощью прибора "Анализатор живых систем, ВТS-310" с использованием реактивов фирмы "Экомедполл", Москва.
Результаты исследования. Тренировка животных по предложенной схеме значительно повышала их работоспособность. Употребляемый экстракт корня солодки оказывался дополнительным стимулом в становлении и повышении адаптивных возможностей крыс. Так, если продолжительность бега в группе животных, находящихся на обычном пищевом рационе, возросла на 63,5%, то животных, получавших экстракт корня солодки в дозе 50 мг/кг массы тела, - на 120%.
Биохимический анализ крови позволил установить следующую динамику изучаемых показателей. Концентрация общего Са в процессе тренировок повышалась. Степень гиперкальциемии коррелировала с объемом и интенсивностью нагрузок. Наиболее значительное повышение Са было отмечено в период окончания 3-го микроцикла. После 4-го, когда объем нагрузок значительно снижался, понижалась и концентрация общего Са (рис. 1 А).
Содержание эритроцитов (рис. 1 Б) в крови в процессе тренировок возрастало, уровень глюкозы (рис. 1 В) снижался в период интенсивных мышечных нагрузок.
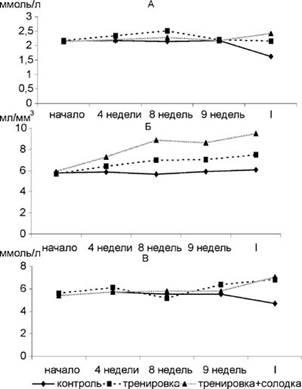
Рис. 1. Динамика общего кальция (А), эритроцитов (Б) и глюкозы (В) на разных этапах тренировочного микроцикла.
Примечание: I - после мышечной нагрузки. Уровень достоверности отличий по сравнению с контролем: р<0,001 - *, р<0,01 - **.
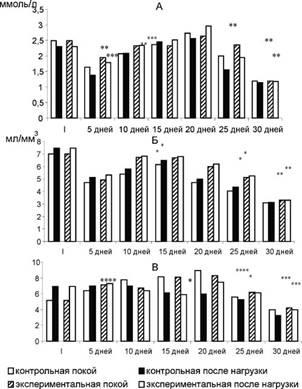
Рис. 2. Влияние дозированной мышечной нагрузки на содержание общего кальция (А), эритроцитов (Б), глюкозы (В) в крови крыс.
Примечание: I - исходный уровень. Уровень достоверности отличий по сравнению с контролем: р<0,001 - *, р<0,01 - **, р<0,02 - ***, р<0,05 - ****.
В том случае, если параллельно с тренировками осуществлялось введение экстракта солодки, волнообразные изменения концентрации Са и глюкозы были незначительными, а прирост количества эритроцитов - достоверно выше. Учитывая роль Са в протекании физиологических функций в живых системах, а также в секреции катехоламинов и гормонов гипоталамо-гипофизарно-надпочечниковой системы, гиперкальциемию, возникающую как в начале однократной нагрузки, так и при интенсификации тренировочного процесса, следует рассматривать как фактор, способствующий активированию адаптивных процессов и компенсаторных реакций организма. Употребление корня солодки снижало степень напряжения компенсаторных механизмов.
Таким образом, употребление экстракта корня солодки значительно облегчает адаптации организма к мышечным нагрузкам. Дополнительным подтверждением может служить концентрация глюкозы. В период интенсивных тренировок она снижалась в крови животных, не принимавших экстракт, что свидетельствует о частичном истощении энергетических запасов в их организме. В 3-й группе колебания концентрации глюкозы были незначительными. Употребление экстракта солодки, по всей видимости, способствовало более экономному расходованию углеводов. Необходимо отметить, что большинство адаптогенов усиливает энергетическое обеспечение работающих органов, стимулируя аэробные окислительные реакции, что, в свою очередь, расширяет круг окислительных субстратов за счет использования липидов. Об усилении мобилизации липидов из жирового депо под влиянием адаптогенов свидетельствует повышение липолитической активности жировой ткани и увеличение в крови их основной транспортной формы - неэстерифицированных жирных кислот. Повышение содержания липидов и их йодного числа в печени свидетельствует об интенсивном использовании жиров работающими тканями [16].
Поступление липидов в печень имеет важное значение в энергетическом обмене, поскольку в этом органе липиды окисляются до легкоутилизируемых веществ, которые при выходе из печени используются мышцами.
Кроме того, более значительное увеличение числа эритроцитов в крови животных, употреблявших экстракт корня солодки, являлось дополнительным фактором, расширяющим адаптивные возможности организма.
Мы уже отмечали, что начальный этап действия любого стрессирующего фактора характеризуется гиперкальциемией. Поэтому повышение содержания Са в крови следует рассматривать как фактор, способствующий активированию эндокринных механизмов адаптации и создающий благоприятные условия для работающих систем. В отличие от этого гипокальциемия - фактор, ограничивающий эту активность и тем самым предохраняющий организм от чрезмерного расходования энергетических и пластических ресурсов. В соответствии с этим способность к поддержанию кальциевого гомеостаза предложено рассматривать как показатель адаптивных возможностей организма [9, 19]. В наших исследованиях выполнение дозированной нагрузки сопровождалось гипокальциемией для нетренированных животных, тенденцией к снижению у тренированных животных и высоким уровнем общего Са в крови крыс, принимавших экстракт солодки на протяжении всего тренировочного цикла. Более экономное расходование углеводов также отмечено в крови животных 3-й группы.
Второй частью наших исследований было изучение динамики исследуемых показателей в период моделирования состояния перетренированности и влияния на эти процессы экстракта солодки. Перетренированность провоцировалась по разработанной нами схеме (табл. 2) у предварительно адаптированных к мышечным нагрузкам животных. Исследования проводились на двух группах животных. В период предварительных тренировок животные находились на обычном пищевом рационе. При моделировании перетренированности животные экспериментальной группы получали экстракт солодки. Основным критерием формирующегося патологического состояния служили снижение массы тела и работоспособности животных, а также результаты биохимического анализа крови до и после выполнения мышечной нагрузки.
Согласно полученным данным снижение работоспособности животных контрольной группы было зарегистрировано после 17 дней эксперимента, т.е. к этому времени животные были не в состоянии полностью восстановиться от предшествующей нагрузки. Все последующие дни работоспособность также снижалась. На 25-й день наблюдений животные в данной группе способны были выполнять нагрузку в течение 89,6±5,8 мин, а через 30 дней - 32,8±3,6 мин. Таким образом, можно предположить, что данный период характеризуется развитием стойкой дизадаптации - перетренированностью. В указанные сроки отмечено и снижение массы тела животных, что также подтверждает развитие патологического состояния.
Биохимический анализ крови, проводимый в период моделирования состояния перетренированности, позволил выявить циклические изменения концентрации изучаемых показателей. Так, через 5 дней эксперимента отмечалось снижение уровня общего Са, эритроцитов и глюкозы (рис. 2), затем, после 20-го дня наблюдений, - повышение и очередное понижение. Содержание общего белка и альбуминов на начальном этапе эксперимента повышалось, а в последующем постоянно понижалось.
Включение в пищевой рацион экстракта солодки голой отодвигало сроки падения работоспособности, но не исключало развития перетренированности. Таким образом, экстракт корня солодки оптимизирует формирование адаптации к мышечным нагрузкам, но оказывается недостаточно эффективным в качестве профилактического средства при развитии такого патологического состояния, как перетренированность. Необходимо отметить, что большинство адаптогенов обладает очень мягким защитным свойством и свое защитное действие реализует посредством нормализации нарушенных функций организма. Этим объясняется слабый эффект или полное его отсутствие при применении адаптогенов в условиях, когда действие внешнего фактора носит ярко выраженный повреждающий характер.
Список литературы
1. Азизов А.Н. Влияние элеутерококка, элтона, левзеи и леветона на систему свертывания крови при физических нагрузках у спортсменов // Экспериментальная и клиническая фармакология. 1997, т. 60, № 5.
2. Беляев Н.Г. Нейроэндокринная регуляция обмена кальция в восстановительном периоде после мышечных нагрузок: Автореф. дис. М., 1987. - 22 с.
3. Беляев Н.Г., Давидянц С.Б., Зырянова О.В. Проблемы развития биологии на Северном Кавказе: Матер. науч. конф. Ставрополь, 1997, с.15-16.
4. Беляев Н.Г., Ермолова Л.С., Батурин В.А. Использование корня солодки как адаптогена // Вестник Российской академии сельскохозяйственных наук. 2001,№ 5, с. 27-29.
5. Беляев Н.Г., Батурин В.А., Солгалов Г.Д. и др. Перспективы использования экстракта солодки голой в спортивной практике // Теор. и практ. физ. культ. 2001, № 6, с. 41-44.
6. Брехман И.И. Женьшень. - Л.: Наука, 1957. - 186 с.
7. Губарева Л.И., Беляев Н.Г., Батурин В.А. и др. Актуальные проблемы экспериментальной и клинической фармакологии: Тез. докл. Всерос. науч. конф. Санкт-Петербург, 2-5 октября 1999 г., с. 56.
8. Дардымов И.В. Женьшень, элеутерококк и механизм биологического действия. - М.: Наука, 1976. - 190 с.
9. Држевецкая И.А., Мишина Н.Ф., Лиманский Н.Н. Гисто-гематические барьеры и нейрогуморальная регуляция: Тез. докл. науч. конф. М. 1981, c. 273-276.
10. Држевецкая И.А., Држевецкий Ю.М. Гормональная регуляция обмена кальция и секреторные процессы // Серия физиолог. человека и животных. ВИНИТИ, М. 1983, т. 27. -131 с.
11. Лиманский Н.Н. Тирокальцитонин: патогенетические основы применения в медицинской практике: Мат. науч.-практ. конф., посвященной 60-летию Великого Октября. Калинин. 1977, с. 94-96.
12. Лиманский Н.Н. Морфо-функциональные особенности растущего организма ребенка: Сб. науч. тр. / АПН СССР, НИИ общей педагогики. М. 1978, с. 43-45.
13. Лиманский Н.Н. Взаимодействие кальцитонина и гормонов гипоталамо-гипофизарно-адренокортикальной системы при мышечной деятельности: Автореф. дис. М., 1981. - 16 с.
14. Похоленчук Ю.П. Функциональное состояние коры надпочечников в ближайшем и отдаленном восстановительном периоде: Автореф. дис. Киев. - 18 с.
15. Романенко В.Д. Физиология кальциевого обмена. Киев, 1975. - 136 с.
16. Саратиков А.С., Краснов Е.А. Родиола розовая - ценное лекарственное растение: Золотой корень. - Томск: Изд-во Том. ун-та, 1987. - 254 с.
17. Селочник Л.И., Брискин А.И., Антонова Е.Е. Фотоэлектроколориметрическое определение концентрации кальция в плазме или сыворотке с применением ЭДТА и мурексида // Химико-фармацевт. журн.1978, т. 12, №10, с. 138-140.
18. Суздальницкий Р.С., Меньшиков И.В., Модера Е.А. Специфические изменения в метаболизме спортсменов, тренирующихся в разных биоэнергетических режимах, в ответ на стандартную физическую нагрузку // Теор. и практ. физ. культ. 2000. №3, с.16-20.
19. Цыбизов Г.Г. Гуморальная регуляция гомеостаза кальция и фосфора при физических нагрузках //Физиол. журн. СССР. 1979, т. 65, №10, с. 1539-1542.
20. Vora N.M., Kukrtja S.C., Iork P.A.J. et al. Effekt of exercise on cerum and parathyroid hormone // Clin. Endocrinol. and Metab. 1983. -V. 57. - №5.- Р. 1067-1069.
|